Autism Spectrum Disorder (ASD) with and without Mental Regression Is Associated with Changes in the Fecal Microbiota

 ,
,  ,
,  ,
,  and
and
Abstract
:1. Introduction
2. Material and Methods
2.1. Ethical Statement
2.2. Participants
2.3. Assessment of Diet
2.4. Metagenomic Analysis
2.4.1. DNA Extraction
2.4.2. Sequencing Analysis
2.4.3. Taxonomic Analysis
2.5. Statistical Analysis
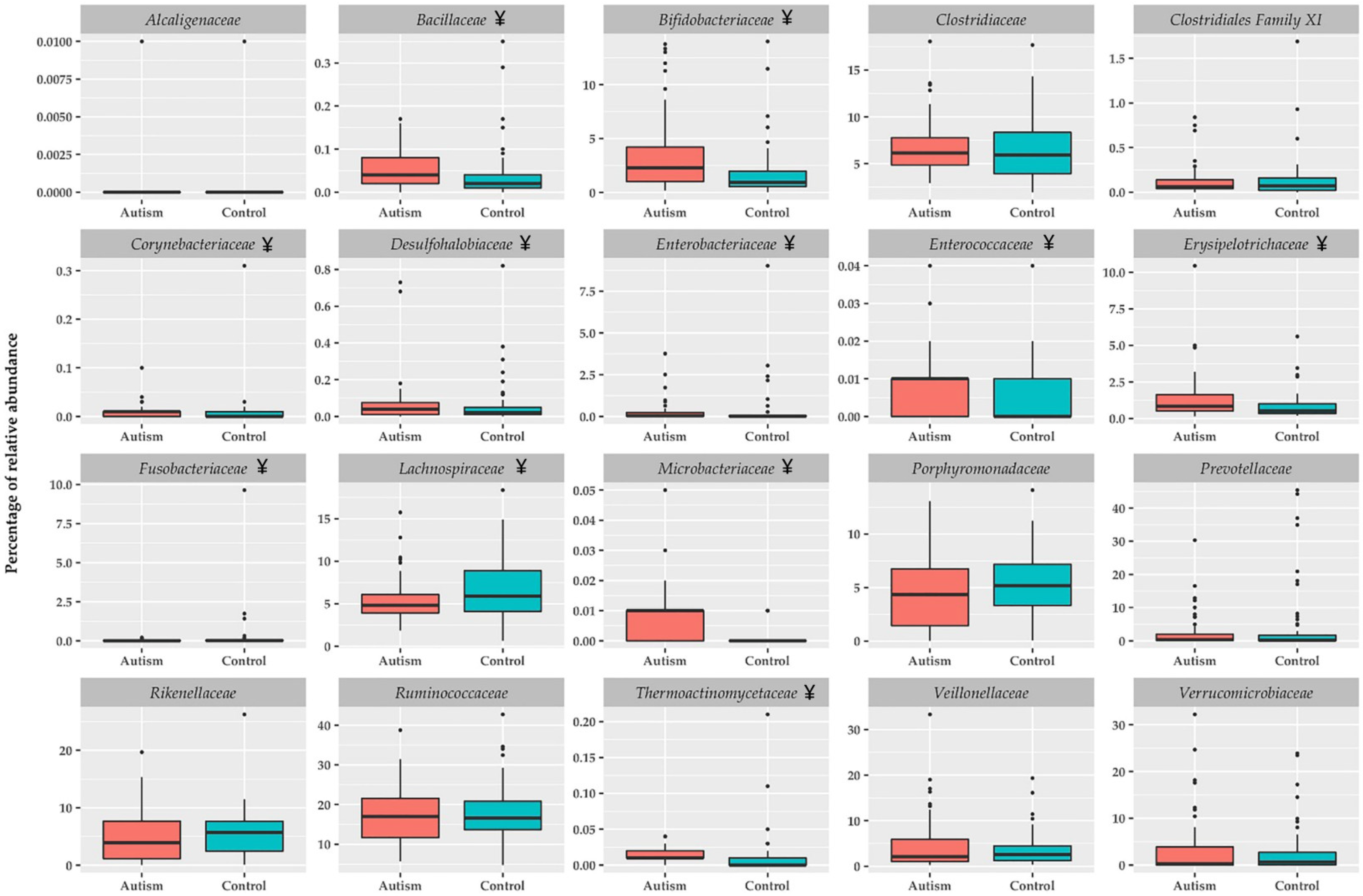
3. Results
ASD Children Show Fecal Metagenomic Differences Compared to Healthy Children
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Gilbert, J.A.; Krajmalnik-Brown, R.; Porazinska, D.L.; Weiss, S.J.; Knight, R. Toward effective probiotics for autism and other neurodevelopmental disorders. Cell 2013, 155, 1446–1448. [Google Scholar] [CrossRef]
- Plaza-Diaz, J.; Ruiz-Ojeda, F.J.; Vilchez-Padial, L.M.; Gil, A. Evidence of the Anti-Inflammatory Effects of Probiotics and Synbiotics in Intestinal Chronic Diseases. Nutrients 2017, 9, 555. [Google Scholar] [CrossRef] [PubMed]
- Plaza-Diaz, J.; Ruiz-Ojeda, F.J.; Gil-Campos, M.; Gil, A. Immune-Mediated Mechanisms of Action of Probiotics and Synbiotics in Treating Pediatric Intestinal Diseases. Nutrients 2018, 10, 42. [Google Scholar] [CrossRef]
- De la Fuente-Nunez, C.; Meneguetti, B.T.; Franco, O.L.; Lu, T.K. Neuromicrobiology: How Microbes Influence the Brain. ACS Chem. Neurosci. 2018, 9, 141–150. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Tian, J.; Yang, B. Targeting gut microbiome: A novel and potential therapy for autism. Life Sci. 2018, 194, 111–119. [Google Scholar] [CrossRef]
- Baio, J.; Wiggins, L.; Christensen, D.L.; Maenner, M.J.; Daniels, J.; Warren, Z.; Kurzius-Spencer, M.; Zahorodny, W.; Robinson Rosenberg, C.; White, T.; et al. Prevalence of Autism Spectrum Disorder Among Children Aged 8 Years—Autism and Developmental Disabilities Monitoring Network, 11 Sites, United States, 2014. Morb. Mortal. Wkly. Rep. Surveill. Summ. 2018, 67, 1–23. [Google Scholar] [CrossRef] [PubMed]
- Morales-Hidalgo, P.; Roige-Castellvi, J.; Hernandez-Martinez, C.; Voltas, N.; Canals, J. Prevalence and Characteristics of Autism Spectrum Disorder Among Spanish School-Age Children. J. Autism Dev. Disord. 2018, 48, 3176–3190. [Google Scholar] [CrossRef]
- Li, Q.; Zhou, J.M. The microbiota-gut-brain axis and its potential therapeutic role in autism spectrum disorder. Neuroscience 2016, 324, 131–139. [Google Scholar] [CrossRef]
- Schieve, L.A.; Gonzalez, V.; Boulet, S.L.; Visser, S.N.; Rice, C.E.; Van Naarden Braun, K.; Boyle, C.A. Concurrent medical conditions and health care use and needs among children with learning and behavioral developmental disabilities, National Health Interview Survey, 2006-2010. Res. Dev. Disabil. 2012, 33, 467–476. [Google Scholar] [CrossRef]
- Kohane, I.S.; McMurry, A.; Weber, G.; MacFadden, D.; Rappaport, L.; Kunkel, L.; Bickel, J.; Wattanasin, N.; Spence, S.; Murphy, S.; et al. The co-morbidity burden of children and young adults with autism spectrum disorders. PLoS ONE 2012, 7, e33224. [Google Scholar] [CrossRef]
- Buie, T.; Fuchs, G.J., 3rd; Furuta, G.T.; Kooros, K.; Levy, J.; Lewis, J.D.; Wershil, B.K.; Winter, H. Recommendations for evaluation and treatment of common gastrointestinal problems in children with ASDs. Pediatrics 2010, 125 (Suppl. 1), S19–S29. [Google Scholar] [CrossRef] [PubMed]
- Emanuele, E.; Orsi, P.; Boso, M.; Broglia, D.; Brondino, N.; Barale, F.; di Nemi, S.U.; Politi, P. Low-grade endotoxemia in patients with severe autism. Neurosci. Lett. 2010, 471, 162–165. [Google Scholar] [CrossRef]
- Ding, H.T.; Taur, Y.; Walkup, J.T. Gut Microbiota and Autism: Key Concepts and Findings. J. Autism Dev. Disord. 2017, 47, 480–489. [Google Scholar] [CrossRef]
- Hsiao, E.Y.; McBride, S.W.; Hsien, S.; Sharon, G.; Hyde, E.R.; McCue, T.; Codelli, J.A.; Chow, J.; Reisman, S.E.; Petrosino, J.F.; et al. Microbiota modulate behavioral and physiological abnormalities associated with neurodevelopmental disorders. Cell 2013, 155, 1451–1463. [Google Scholar] [CrossRef] [PubMed]
- Foley, K.A.; MacFabe, D.F.; Kavaliers, M.; Ossenkopp, K.P. Sexually dimorphic effects of prenatal exposure to lipopolysaccharide, and prenatal and postnatal exposure to propionic acid, on acoustic startle response and prepulse inhibition in adolescent rats: Relevance to autism spectrum disorders. Behav. Brain Res. 2015, 278, 244–256. [Google Scholar] [CrossRef] [PubMed]
- O’Mahony, S.M.; Clarke, G.; Borre, Y.E.; Dinan, T.G.; Cryan, J.F. Serotonin, tryptophan metabolism and the brain-gut-microbiome axis. Behav. Brain Res. 2015, 277, 32–48. [Google Scholar] [CrossRef] [PubMed]
- Hornig, M. The role of microbes and autoimmunity in the pathogenesis of neuropsychiatric illness. Curr. Opin. Rheumatol. 2013, 25, 488–795. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Christophersen, C.T.; Sorich, M.J.; Gerber, J.P.; Angley, M.T.; Conlon, M.A. Low relative abundances of the mucolytic bacterium Akkermansia muciniphila and Bifidobacterium spp. in feces of children with autism. Appl. Environ. Microbiol. 2011, 77, 6718–6721. [Google Scholar] [CrossRef]
- Sandler, R.H.; Finegold, S.M.; Bolte, E.R.; Buchanan, C.P.; Maxwell, A.P.; Vaisanen, M.L.; Nelson, M.N.; Wexler, H.M. Short-term benefit from oral vancomycin treatment of regressive-onset autism. J. Child Neurol. 2000, 15, 429–435. [Google Scholar] [CrossRef]
- Finegold, S.M. Therapy and epidemiology of autism--clostridial spores as key elements. Med. Hypotheses 2008, 70, 508–511. [Google Scholar] [CrossRef]
- Finegold, S.M.; Dowd, S.E.; Gontcharova, V.; Liu, C.; Henley, K.E.; Wolcott, R.D.; Youn, E.; Summanen, P.H.; Granpeesheh, D.; Dixon, D.; et al. Pyrosequencing study of fecal microflora of autistic and control children. Anaerobe 2010, 16, 444–453. [Google Scholar] [CrossRef]
- Gomez-Fernandez, A.; de la Torre-Aguilar, M.J.; Gil-Campos, M.; Flores-Rojas, K.; Cruz-Rico, M.D.; Martin-Borreguero, P.; Perez-Navero, J.L. Children with Autism Spectrum Disorder With Regression Exhibit a Different Profile in Plasma Cytokines and Adhesion Molecules Compared to Children Without Such Regression. Front. Pediatr. 2018, 6, 264. [Google Scholar] [CrossRef] [PubMed]
- Kern, J.K.; Geier, D.A.; Geier, M.R. Evaluation of regression in autism spectrum disorder based on parental reports. N. Am. J. Med. Sci. 2014, 6, 41–47. [Google Scholar] [CrossRef] [PubMed]
- Iovene, M.R.; Bombace, F.; Maresca, R.; Sapone, A.; Iardino, P.; Picardi, A.; Marotta, R.; Schiraldi, C.; Siniscalco, D.; Serra, N.; et al. Intestinal Dysbiosis and Yeast Isolation in Stool of Subjects with Autism Spectrum Disorders. Mycopathologia 2017, 182, 349–363. [Google Scholar] [CrossRef]
- Carding, S.; Verbeke, K.; Vipond, D.T.; Corfe, B.M.; Owen, L.J. Dysbiosis of the gut microbiota in disease. Microb. Ecol. Health Dis. 2015, 26, 26191. [Google Scholar] [CrossRef] [PubMed]
- Buie, T. Potential Etiologic Factors of Microbiome Disruption in Autism. Clin. Ther. 2015, 37, 976–983. [Google Scholar] [CrossRef] [PubMed]
- Adams, J.B.; Johansen, L.J.; Powell, L.D.; Quig, D.; Rubin, R.A. Gastrointestinal flora and gastrointestinal status in children with autism--comparisons to typical children and correlation with autism severity. BMC Gastroenterol. 2011, 11, 22. [Google Scholar] [CrossRef]
- Mulak, A.; Bonaz, B. Irritable bowel syndrome: A model of the brain-gut interactions. Med. Sci. Monit. 2004, 10, RA55–RA62. [Google Scholar]
- Forsythe, P.; Bienenstock, J.; Kunze, W.A. Vagal pathways for microbiome-brain-gut axis communication. Adv. Exp. Med. Biol. 2014, 817, 115–133. [Google Scholar] [CrossRef]
- Bravo, J.A.; Forsythe, P.; Chew, M.V.; Escaravage, E.; Savignac, H.M.; Dinan, T.G.; Bienenstock, J.; Cryan, J.F. Ingestion of Lactobacillus strain regulates emotional behavior and central GABA receptor expression in a mouse via the vagus nerve. Proc. Natl. Acad. Sci. USA 2011, 108, 16050–16055. [Google Scholar] [CrossRef]
- Zhu, X.; Han, Y.; Du, J.; Liu, R.; Jin, K.; Yi, W. Microbiota-gut-brain axis and the central nervous system. Oncotarget 2017, 8, 53829–53838. [Google Scholar] [CrossRef] [PubMed]
- Araque, A.; Carmignoto, G.; Haydon, P.G.; Oliet, S.H.; Robitaille, R.; Volterra, A. Gliotransmitters travel in time and space. Neuron 2014, 81, 728–739. [Google Scholar] [CrossRef] [PubMed]
- Chung, W.; Choi, S.Y.; Lee, E.; Park, H.; Kang, J.; Park, H.; Choi, Y.; Lee, D.; Park, S.G.; Kim, R.; et al. Social deficits in IRSp53 mutant mice improved by NMDAR and mGluR5 suppression. Nat. Neurosci. 2015, 18, 435–443. [Google Scholar] [CrossRef] [PubMed]
- Erny, D.; Hrabe de Angelis, A.L.; Jaitin, D.; Wieghofer, P.; Staszewski, O.; David, E.; Keren-Shaul, H.; Mahlakoiv, T.; Jakobshagen, K.; Buch, T.; et al. Host microbiota constantly control maturation and function of microglia in the CNS. Nat. Neurosci. 2015, 18, 965–977. [Google Scholar] [CrossRef] [PubMed]
- Morgan, J.T.; Chana, G.; Abramson, I.; Semendeferi, K.; Courchesne, E.; Everall, I.P. Abnormal microglial-neuronal spatial organization in the dorsolateral prefrontal cortex in autism. Brain Res. 2012, 1456, 72–81. [Google Scholar] [CrossRef] [PubMed]
- Edmonson, C.; Ziats, M.N.; Rennert, O.M. Altered glial marker expression in autistic post-mortem prefrontal cortex and cerebellum. Mol. Autism 2014, 5, 3. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, K.; Sugihara, G.; Ouchi, Y.; Nakamura, K.; Futatsubashi, M.; Takebayashi, K.; Yoshihara, Y.; Omata, K.; Matsumoto, K.; Tsuchiya, K.J.; et al. Microglial activation in young adults with autism spectrum disorder. JAMA Psychiatry 2013, 70, 49–58. [Google Scholar] [CrossRef] [PubMed]
- Sharon, G.; Sampson, T.R.; Geschwind, D.H.; Mazmanian, S.K. The Central Nervous System and the Gut Microbiome. Cell 2016, 167, 915–932. [Google Scholar] [CrossRef]
- Lu, J.; Lu, L.; Yu, Y.; Cluette-Brown, J.; Martin, C.R.; Claud, E.C. Effects of Intestinal Microbiota on Brain Development in Humanized Gnotobiotic Mice. Sci. Rep. 2018, 8, 5443. [Google Scholar] [CrossRef]
- Bjorklund, G. The role of zinc and copper in autism spectrum disorders. Acta Neurobiol. Exp. 2013, 73, 225–236. [Google Scholar]
- Ong, I.M.; Gonzalez, J.G.; McIlwain, S.J.; Sawin, E.A.; Schoen, A.J.; Adluru, N.; Alexander, A.L.; Yu, J.J. Gut microbiome populations are associated with structure-specific changes in white matter architecture. Transl. Psychiatry 2018, 8, 6. [Google Scholar] [CrossRef] [PubMed]
- World Health Organization. The ICD-10 Classification of Mental and Behavioural Disorders: Clinical Descriptions and Diagnostic Guidelines; World Health Organization: Geneva, Switzerland, 1992; Volume 1. [Google Scholar]
- American Psychiatric Pub. Diagnostic and Statistical Manual of Mental Disorders (DSM-5®); American Psychiatric Pub: Washington, DC, USA, 2013. [Google Scholar]
- Kim, S.H.; Lord, C. New autism diagnostic interview-revised algorithms for toddlers and young preschoolers from 12 to 47 months of age. J. Autism Dev. Disord. 2012, 42, 82–93. [Google Scholar] [CrossRef] [PubMed]
- Aranceta Bartrina, J. Role of gastronomy and new technologies in shaping a healthy diet. Nutricion Hospitalaria 2018, 35, 3–9. [Google Scholar] [CrossRef] [PubMed]
- Herlemann, D.P.; Labrenz, M.; Jurgens, K.; Bertilsson, S.; Waniek, J.J.; Andersson, A.F. Transitions in bacterial communities along the 2000 km salinity gradient of the Baltic Sea. ISME J. 2011, 5, 1571–1579. [Google Scholar] [CrossRef] [PubMed]
- Schloss, P.D.; Westcott, S.L.; Ryabin, T.; Hall, J.R.; Hartmann, M.; Hollister, E.B.; Lesniewski, R.A.; Oakley, B.B.; Parks, D.H.; Robinson, C.J.; et al. Introducing mothur: Open-source, platform-independent, community-supported software for describing and comparing microbial communities. Appl. Environ. Microbiol. 2009, 75, 7537–7541. [Google Scholar] [CrossRef] [PubMed]
- Meyer, F.; Paarmann, D.; D’Souza, M.; Olson, R.; Glass, E.M.; Kubal, M.; Paczian, T.; Rodriguez, A.; Stevens, R.; Wilke, A.; et al. The metagenomics RAST server—A public resource for the automatic phylogenetic and functional analysis of metagenomes. BMC Bioinform. 2008, 9, 386. [Google Scholar] [CrossRef] [PubMed]
- Marques, R.C.; Bernardi, J.V.; Dorea, J.G.; Bastos, W.R.; Malm, O. Principal component analysis and discrimination of variables associated with pre- and post-natal exposure to mercury. Int. J. Hyg. Environ. Health 2008, 211, 606–614. [Google Scholar] [CrossRef]
- Weiss, R.; Dziura, J.; Burgert, T.S.; Tamborlane, W.V.; Taksali, S.E.; Yeckel, C.W.; Allen, K.; Lopes, M.; Savoye, M.; Morrison, J.; et al. Obesity and the metabolic syndrome in children and adolescents. N. Engl. J. Med. 2004, 350, 2362–2374. [Google Scholar] [CrossRef]
- De Angelis, M.; Francavilla, R.; Piccolo, M.; De Giacomo, A.; Gobbetti, M. Autism spectrum disorders and intestinal microbiota. Gut Microbes 2015, 6, 207–213. [Google Scholar] [CrossRef]
- Mead, J.; Ashwood, P. Evidence supporting an altered immune response in ASD. Immunol. Lett. 2015, 163, 49–55. [Google Scholar] [CrossRef]
- De Angelis, M.; Piccolo, M.; Vannini, L.; Siragusa, S.; De Giacomo, A.; Serrazzanetti, D.I.; Cristofori, F.; Guerzoni, M.E.; Gobbetti, M.; Francavilla, R. Fecal microbiota and metabolome of children with autism and pervasive developmental disorder not otherwise specified. PLoS ONE 2013, 8, e76993. [Google Scholar] [CrossRef] [PubMed]
- Parracho, H.M.; Bingham, M.O.; Gibson, G.R.; McCartney, A.L. Differences between the gut microflora of children with autistic spectrum disorders and that of healthy children. J. Med. Microbiol. 2005, 54, 987–991. [Google Scholar] [CrossRef] [PubMed]
- Ruiz, L.; Delgado, S.; Ruas-Madiedo, P.; Sanchez, B.; Margolles, A. Bifidobacteria and Their Molecular Communication with the Immune System. Front. Microbiol. 2017, 8, 2345. [Google Scholar] [CrossRef] [PubMed]
- Bermudez-Brito, M.; Plaza-Diaz, J.; Munoz-Quezada, S.; Gomez-Llorente, C.; Gil, A. Probiotic mechanisms of action. Ann. Nutr. Metab. 2012, 61, 160–174. [Google Scholar] [CrossRef] [PubMed]
- Shiby, V.K.; Mishra, H.N. Fermented milks and milk products as functional foods—A review. Crit. Rev. Food Sci. Nutr. 2013, 53, 482–496. [Google Scholar] [CrossRef] [PubMed]
- Palaria, A.; Johnson-Kanda, I.; O’Sullivan, D.J. Effect of a synbiotic yogurt on levels of fecal bifidobacteria, clostridia, and enterobacteria. Appl. Environ. Microbiol. 2012, 78, 933–940. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, Y.; Takami, K.; Nishijima, T.; Aoki, R.; Mawatari, T.; Ikeda, T. Short- and long-term dynamics in the intestinal microbiota following ingestion of Bifidobacterium animalis subsp. lactis GCL2505. Biosci. Microbiota Food Health 2015, 34, 77–85. [Google Scholar] [CrossRef] [PubMed]
- Plovier, H.; Everard, A.; Druart, C.; Depommier, C.; Van Hul, M.; Geurts, L.; Chilloux, J.; Ottman, N.; Duparc, T.; Lichtenstein, L.; et al. A purified membrane protein from Akkermansia muciniphila or the pasteurized bacterium improves metabolism in obese and diabetic mice. Nat. Med. 2017, 23, 107–113. [Google Scholar] [CrossRef]
- Kang, D.W.; Park, J.G.; Ilhan, Z.E.; Wallstrom, G.; Labaer, J.; Adams, J.B.; Krajmalnik-Brown, R. Reduced incidence of Prevotella and other fermenters in intestinal microflora of autistic children. PLoS ONE 2013, 8, e68322. [Google Scholar] [CrossRef]
- Ley, R.E. Gut microbiota in 2015: Prevotella in the gut: Choose carefully. Nat. Rev. Gastroenterol. Hepatol. 2016, 13, 69–70. [Google Scholar] [CrossRef]
- Kovatcheva-Datchary, P.; Nilsson, A.; Akrami, R.; Lee, Y.S.; De Vadder, F.; Arora, T.; Hallen, A.; Martens, E.; Bjorck, I.; Backhed, F. Dietary Fiber-Induced Improvement in Glucose Metabolism Is Associated with Increased Abundance of Prevotella. Cell Metab. 2015, 22, 971–982. [Google Scholar] [CrossRef] [PubMed]
- Quigley, E.M. Leaky gut—Concept or clinical entity? Curr. Opin. Gastroenterol. 2016, 32, 74–79. [Google Scholar] [CrossRef]
- Onore, C.; Careaga, M.; Ashwood, P. The role of immune dysfunction in the pathophysiology of autism. Brain Behav. Immunity 2012, 26, 383–392. [Google Scholar] [CrossRef] [PubMed]
- Williams, B.L.; Hornig, M.; Parekh, T.; Lipkin, W.I. Application of novel PCR-based methods for detection, quantitation, and phylogenetic characterization of Sutterella species in intestinal biopsy samples from children with autism and gastrointestinal disturbances. mBio 2012, 3. [Google Scholar] [CrossRef]
- Dethlefsen, L.; McFall-Ngai, M.; Relman, D.A. An ecological and evolutionary perspective on human-microbe mutualism and disease. Nature 2007, 449, 811–818. [Google Scholar] [CrossRef]
- Connolly, N.; Anixt, J.; Manning, P.; Ping, I.L.D.; Marsolo, K.A.; Bowers, K. Maternal metabolic risk factors for autism spectrum disorder-An analysis of electronic medical records and linked birth data. Autism Res. 2016, 9, 829–837. [Google Scholar] [CrossRef]
- Buffington, S.A.; Di Prisco, G.V.; Auchtung, T.A.; Ajami, N.J.; Petrosino, J.F.; Costa-Mattioli, M. Microbial Reconstitution Reverses Maternal Diet-Induced Social and Synaptic Deficits in Offspring. Cell 2016, 165, 1762–1775. [Google Scholar] [CrossRef]
- Mari-Bauset, S.; Zazpe, I.; Mari-Sanchis, A.; Llopis-Gonzalez, A.; Morales-Suarez-Varela, M. Evidence of the gluten-free and casein-free diet in autism spectrum disorders: A systematic review. J. Child Neurol. 2014, 29, 1718–1727. [Google Scholar] [CrossRef] [PubMed]
- Patusco, R.; Ziegler, J. Role of Probiotics in Managing Gastrointestinal Dysfunction in Children with Autism Spectrum Disorder: An Update for Practitioners. Adv. Nutr. 2018, 9, 637–650. [Google Scholar] [CrossRef] [PubMed]
- Berding, K.; Donovan, S.M. Microbiome and nutrition in autism spectrum disorder: Current knowledge and research needs. Nutr. Rev. 2016, 74, 723–736. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Variables | ASD | Control (n = 57) | P Value | ||
|---|---|---|---|---|---|
| ANMR (n = 30) | AMR (n = 18) | Total (n = 48) | |||
| Age (months) | 44.51 ± 2.06 | 43.69 ± 2.7 | 44.19 ± 1.6 | 51.00 ± 2.59 | n.s. |
| Weight (kg) | 16.57 ± 0.56 | 17.10 ± 0.95 | 16.77 ± 0.50 | 17.1 ± 0.6 | n.s. |
| Height (cm) | 103.3 ± 1.55 | 101.50 ± 1.92 | 102.5 ± 1.20 | 102.0 ± 1.5 | n.s. |
| BMI (kg/cm2) | 15.52 ± 0.33 | 16.37 ± 0.39 | 15.86 ± 0.26 | 16.2 ± 0.2 | n.s. |
| Battelle test | 59.6 ± 2.6 | 47.5 ± 2.6 | 54.98 ± 2.0 | - | 0.003 |
| CARS test | 30.6 ± 1.1 | 35.9 ± 1.9 | 32.7 ± 1.1 | - | 0.021 |
| PDDBI test | 46.8 ± 2.3 | 53.6 ± 1.6 | 50.9 ± 1.6 | - | 0.026 |
| Variables | ASD | |||
|---|---|---|---|---|
| ANMR Group (n = 30) | AMR Group (n = 18) | Total ASD (n = 48) | Healthy Control Group (n = 57) | |
| Actinobacteria (phylum) | 2.6 (0.6–14.9) a | 3.2 (0.3–16.8) ab | 2.9 (0.3–16.8) * | 1.8 (0.1–18.3) b |
| Bacteroidetes (phylum) | 43.4 (2.0–58.1) | 39.5 (11.0–51.7) | 43.0 (2.0–58.1) | 42.9 (8.5–67.9) |
| Firmicutes (phylum) | 45.4 (31.0–82.9) | 44.0 (19.6–61.6) | 44.7 (19.6–82.9) | 42.2 (19.2–81.1) |
| Proteobacteria (phylum) | 0.2 (0.0–4.1) a | 0.4 (0.1–2.8) b | 0.4 (0.0–4.1) * | 0.2 (0.0–8.9) a |
| Verrucomicrobia (phylum) | 0.1 (0.0–24.5) | 1.1 (0.0–30.9) | 0.3 (0.0–30.9) | 0.7 (0.0–23.2) |
| Actinobacteria (class) | 2.6 (0.6–14.9) a | 3.2 (0.3–16.8) ab | 2.9 (0.3–16.8) * | 1.8 (0.0–18.3) b |
| Bacilli (class) | 0.4 (0.1–6.4) | 0.4 (0.0–1.9) | 0.4 (0.0–6.4) * | 0.3 (0.0–2.4) |
| Bacteroidia (class) | 43.3 (1.9–58.1) | 39.5 (7.7–51.7) | 42.9 (1.9–58.1) | 42.9 (8.5–67.9) |
| Clostridia (class) | 36.0 (22.9–51.3) | 35.6 (17.2–57.1) | 35.6 (17.2–57.1) | 37.5 (12.5–65.8) |
| Deltaproteobacteria (class) | 0.1 (0.0–0.8) | 0.1 (0.0–1.4) | 0.1 (0.0–1.4) | 0.07 (0.0–0.9) |
| Erysipelotrichi (class) | 0.6 (0.2–10.1) | 0.9 (0.1–4.8) | 0.8 (0.1–10.1) * | 0.5 (0.2–5.5) |
| Gammaproteobacteria (class) | 0.1 (0.0–3.7) | 0.2 (0.0–2.7) | 0.1 (0.0–3.7) * | 0.04 (0.0–8.8) |
| Negativicutes (class) | 4.6 (0.2–32.4) | 1.9 (0.4–15.4) | 2.8 (0.2–32.4) | 2.9 (0.5–20.8) |
| Verrucomicrobiae (class) | 0.1 (0.0–24.5) | 1.1 (0.0–30.9) | 0.3 (0.0–30.9) | 0.7 (0.0–23.2) |
| Unclassified sequences derived from Bacteria | 4.7 (1.4–13.3) a | 9.7 (2.2–32.3) b | 5.9 (1.4–32.3) | 8.0 (1.4–31.8) b |
| Alpha diversity | 33.5 (17.0–86.0) | 28.5 (9.0–55.0) | 30.5 (9.0–86.0) | 32.0 (12.0–62.0) |
| Variables | ASD | Control (n = 57) | ||
|---|---|---|---|---|
| ANMR (n = 30) | AMR (n = 18) | Total (n = 48) | ||
| Akkermansia | 0.06 (0.0–24.8) | 1.1 (0.0–32.3) | 0.30 (0.0–32.3) | 0.7 (0.0–24.2) |
| Alistipes | 3.9 (0.0–13.8) | 3.8 (0.0–20.0) | 3.9 (0.0–20.0) | 5.7 (0.0–26.1) |
| Bacillus | 0.03 (0.0–0.1) | 0.03 (0.0–0.1) | 0.03 (0.0–0.1) * | 0.02 (0.0–0.4) |
| Bacteroides | 30.3 (1.1–60.8) | 23.2 (4.0–48.4) | 28.6 (1.1–60.8) | 29.4 (2.4–51.3) |
| Bifidobacterium | 2.2 (0.2–13.8) a | 2.4 (0.3–14.2) a | 2.3 (0.2–14.2) * | 0.9 (0.0–14.0) b |
| Butyrivibrio | 1.0 (0.2–5.3) | 1.0 (0.1–4.7) | 1.0 (0.1–5.3) * | 1.4 (0.2–6.8) |
| Clostridium | 6.1 (3.0–11.7) | 5.7 (3.0–16.9) | 5.8 (3.0–16.9) | 5.4 (1.8–16.9) |
| Collinsella | 0.4 (0.0–4.4) | 0.2 (0.0–8.2) | 0.4 (0.0–8.2) | 0.5 (0.0–5.0) |
| Desulfovibrio | 0.001 (0.0–0.5) | 0.002 (0.0–1.3) | 0.002 (0.0–1.3) | 0.0005 (0.0–0.3) |
| Enterococcus | 0.002 (0.0–0.04) a | 0.004 (0.0–0.01) b | 0.004 (0.0–0.04) * | 0.001 (0.0–0.04) a |
| Eubacterium | 4.0 (0.5–10.9) | 2.6 (0.3–8.4) | 3.6 (0.3–10.9) | 2.6 (0.5–13.5) |
| Faecalibacterium | 11.7 (1.4–22.8) | 9.6 (1.8–37.0) | 10.7 (1.4–37.0) | 11.4 (2.7–37.1) |
| Hespellia | 0.2 (0.0–1.9) | 0.2 (0.0–0.7) | 0.2 (0.0–1.9) * | 0.1 (0.0–0.9) |
| Lactobacillus | 0.05 (0.0–3.5) | 0.03 (0.0–0.6) | 0.04 (0.0–3.5) | 0.03 (0.0–1.0) |
| Parabacteroides | 2.6 (0.0–8.3) | 2.1 (0.0–5.5) | 2.4 (0.0–8.3) | 1.9 (0.0–10.0) |
| Prevotella | 0.3 (0.0–30.2) | 0.2 (0.0–11.9) | 0.3 (0.0–30.2) * | 0.1 (0.0–43.7) |
| Ruminococcus | 3.1 (1.1–10.3) | 3.3 (0.9–15.8) | 3.1 (0.9–15.8) | 3.2 (0.7–26.5) |
| Veillonella | 0.4 (0.1–34.9) | 0.6 (0.0–5.8) | 0.5 (0.0–34.9) | 0.7 (0.0–19.3) |
| Bacteroides fragilis | 0.33 (0.0–9.1) | 0.39 (0.0–7.9) | 0.35 (0.0–9.1) | 0.34 (0.0–32.1) |
| Bacteroides vulgatus | 2.9 (0.0–28.1) | 1.2 (0.0–36.7) | 1.4 (0.0–36.7) | 8.5 (0.0–36.3) |
| Clostridium bolteae | 0.1 (0.0–1.7) a | 0.1 (0.0–1.2) a | 0.1 (0.0–1.7) * | 0.04 (0.0–1.4) b |
| Clostridium difficile | 0.08 (0.0–0.6) | 0.04 (0.0–0.6) | 0.09 (0.0–0.6) * | 0.06 (0.0–0.8) |
| Faecalibacterium prausnitzii | 11.3 (1.4–21.9) | 9.1 (1.7–36.0) | 10.3 (1.4–36.0) | 11.3 (2.6–35.7) |
| Ruminococcus gnavus | 0.3 (0.0–4.5) | 0.2 (0.0–1.7) | 0.3 (0.0–4.5) | 0.3 (0.0–7.6) |
| Ruminococcus torques | 0.1 (0.0–0.9) | 0.1 (0.0–3.2) | 0.1 (0.0–3.2) | 0.08 (0.0–5.0) |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Plaza-Díaz, J.; Gómez-Fernández, A.; Chueca, N.; Torre-Aguilar, M.J.d.l.; Gil, Á.; Perez-Navero, J.L.; Flores-Rojas, K.; Martín-Borreguero, P.; Solis-Urra, P.; Ruiz-Ojeda, F.J.; et al. Autism Spectrum Disorder (ASD) with and without Mental Regression Is Associated with Changes in the Fecal Microbiota. Nutrients 2019, 11, 337. https://doi.org/10.3390/nu11020337
Plaza-Díaz J, Gómez-Fernández A, Chueca N, Torre-Aguilar MJdl, Gil Á, Perez-Navero JL, Flores-Rojas K, Martín-Borreguero P, Solis-Urra P, Ruiz-Ojeda FJ, et al. Autism Spectrum Disorder (ASD) with and without Mental Regression Is Associated with Changes in the Fecal Microbiota. Nutrients. 2019; 11(2):337. https://doi.org/10.3390/nu11020337
Chicago/Turabian StylePlaza-Díaz, Julio, Antonio Gómez-Fernández, Natalia Chueca, María José de la Torre-Aguilar, Ángel Gil, Juan Luis Perez-Navero, Katherine Flores-Rojas, Pilar Martín-Borreguero, Patricio Solis-Urra, Francisco Javier Ruiz-Ojeda, and et al. 2019. "Autism Spectrum Disorder (ASD) with and without Mental Regression Is Associated with Changes in the Fecal Microbiota" Nutrients 11, no. 2: 337. https://doi.org/10.3390/nu11020337
APA StylePlaza-Díaz, J., Gómez-Fernández, A., Chueca, N., Torre-Aguilar, M. J. d. l., Gil, Á., Perez-Navero, J. L., Flores-Rojas, K., Martín-Borreguero, P., Solis-Urra, P., Ruiz-Ojeda, F. J., Garcia, F., & Gil-Campos, M. (2019). Autism Spectrum Disorder (ASD) with and without Mental Regression Is Associated with Changes in the Fecal Microbiota. Nutrients, 11(2), 337. https://doi.org/10.3390/nu11020337