
Licorice Extract Isoliquiritigenin Protects Endothelial Function in Type 2 Diabetic Mice

 ,
, 
Abstract
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Materials and Methods
2.1. Drugs and Solutions
2.2. Animal Models
2.3. Measurements of Basic Parameters
2.4. Mice Arterial Preparation and Vascular Reactivity Study Determined by Wire Myograph
2.5. Cell Culture
2.6. Reactive Oxygen Species (ROS) Measurement
2.7. Quantitative Polymerase Chain Reaction (qPCR) Analysis
2.8. Statistical Analysis
3. Results
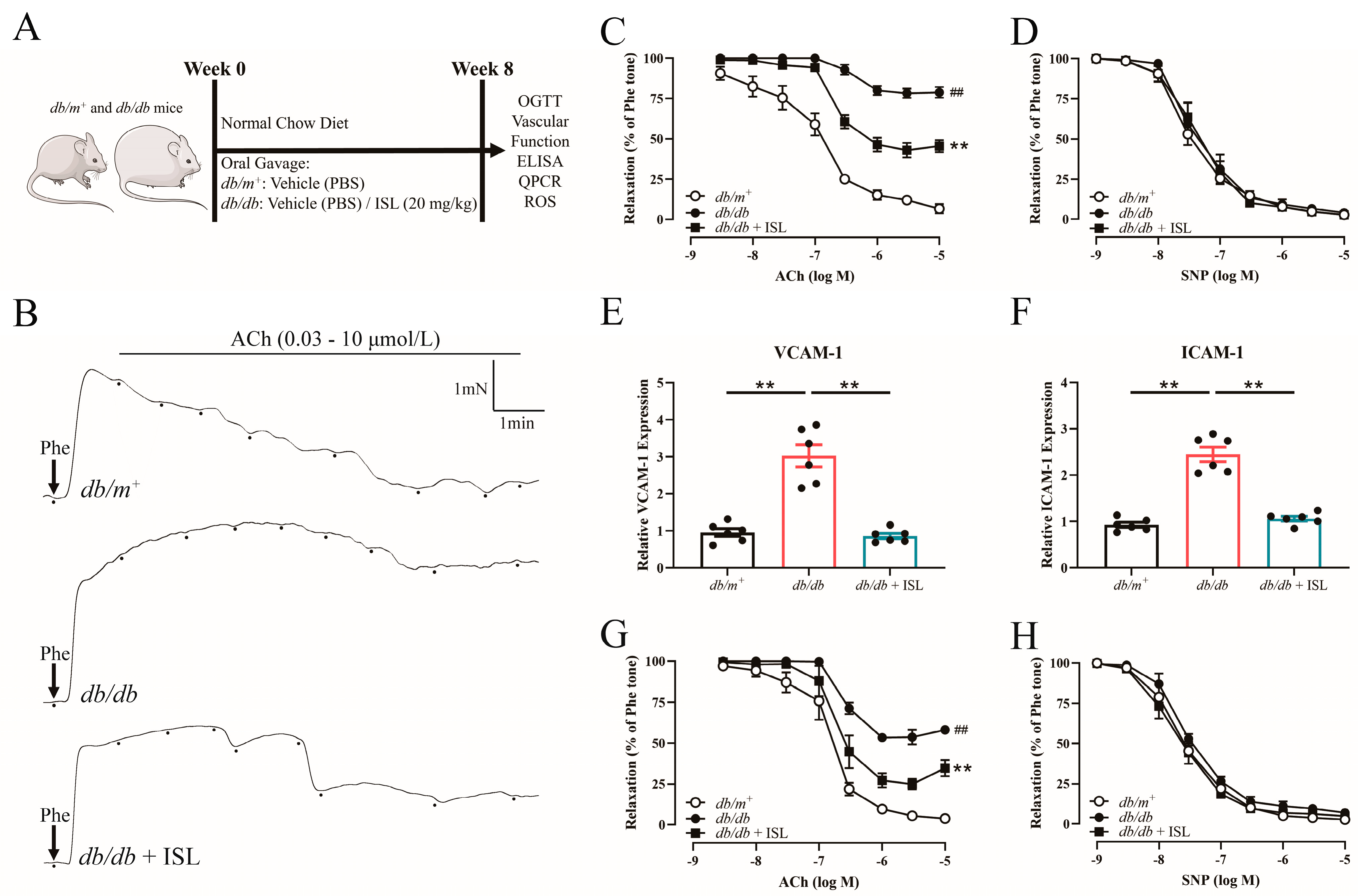
3.1. ISL Oral Treatment Reverses Endothelial Dysfunction in db/db Mouse Aortas
3.2. ISL Reverses the Impairment of Endothelium-Dependent Relaxations (EDRs) in db/db Mouse Aortas in Ex Vivo Studies
3.3. Basic Parameters of Mice Treated with ISL
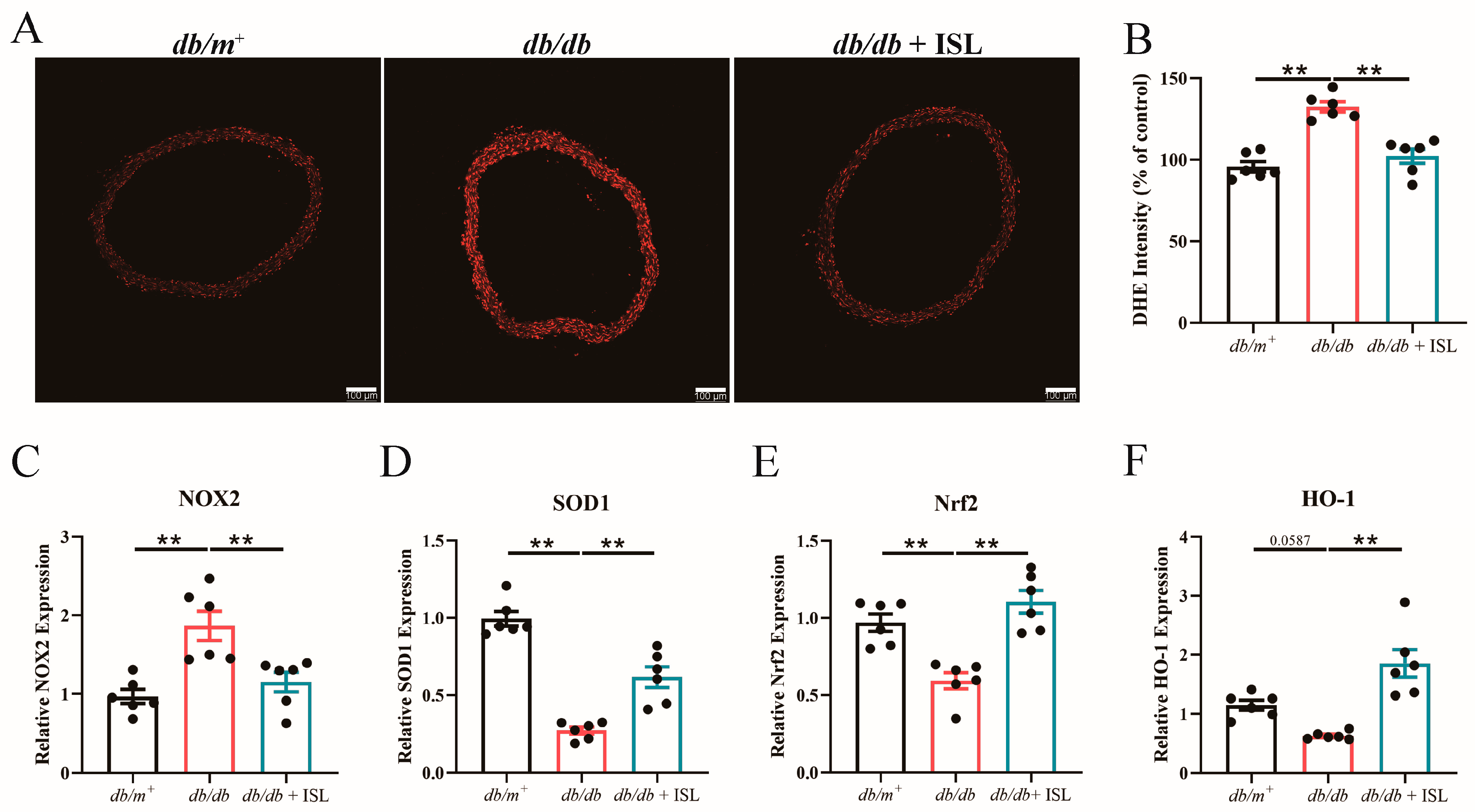
3.4. Oral Treatment of ISL Attenuates Oxidative Stress in db/db Mice
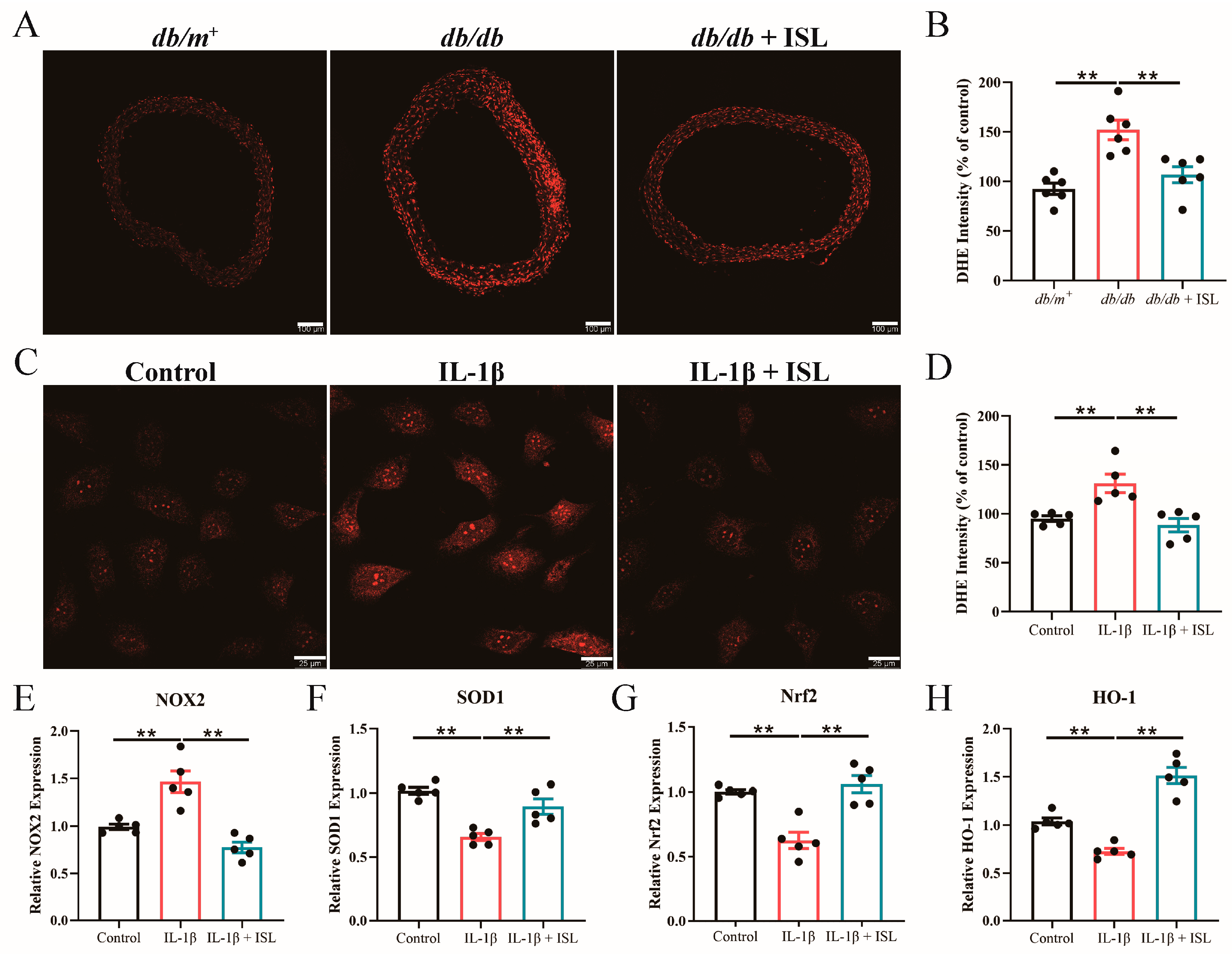
3.5. ISL Reduces ROS Levels in db/db Mouse Aortas in Ex Vivo Studies
3.6. ISL Alleviates Oxidative Stress Triggered by IL-1β in Endothelial Cells
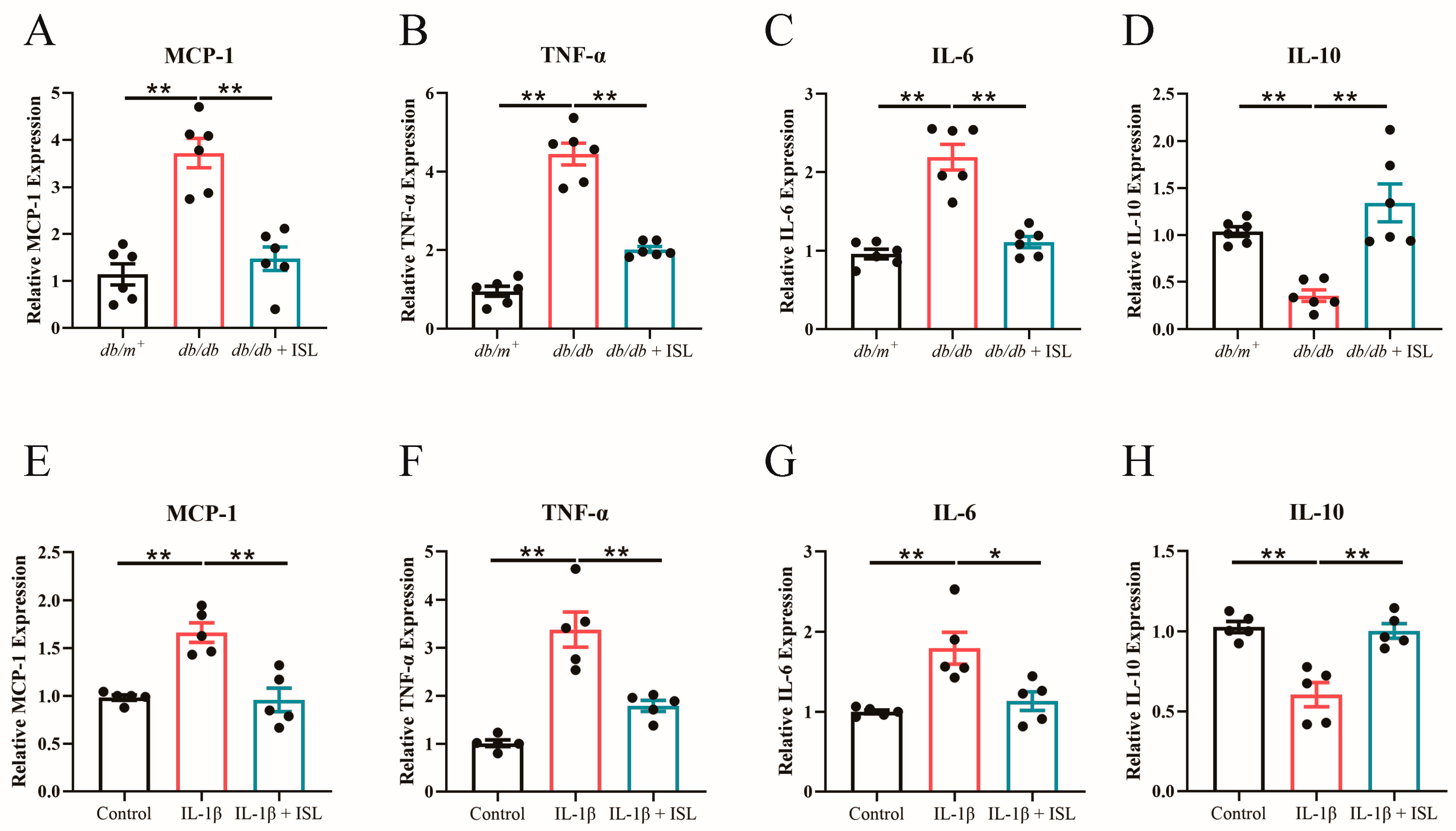
3.7. ISL Attenuates Inflammation in db/db Mouse Aortas and Endothelial Cells
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Gaziano, T.; Reddy, K.S.; Paccaud, F.; Horton, S.; Chaturvedi, V. Cardiovascular disease. In Disease Control Priorities in Developing Countries, 2nd ed.; The International Bank for Reconstruction and Development: Washington, DC, USA, 2006. [Google Scholar]
- Hadi, H.A.; Carr, C.S.; Al Suwaidi, J. Endothelial dysfunction: Cardiovascular risk factors, therapy, and outcome. Vasc. Health Risk Manag. 2005, 1, 183–198. [Google Scholar] [PubMed]
- Kasprzak, J.D.; Kłosińska, M.; Drozdz, J. Clinical aspects of assessment of endothelial function. Pharmacol. Rep. 2006, 58, 33–40. [Google Scholar] [PubMed]
- Park, K.H.; Park, W.J. Endothelial Dysfunction: Clinical Implications in Cardiovascular Disease and Therapeutic Approaches. J. Korean Med. Sci. 2015, 30, 1213–1225. [Google Scholar] [CrossRef] [PubMed]
- Versari, D.; Daghini, E.; Virdis, A.; Ghiadoni, L.; Taddei, S. Endothelial dysfunction as a target for prevention of cardiovascular disease. Diabetes Care 2009, 32 (Suppl. S2), S314. [Google Scholar] [CrossRef]
- Gimbrone, M.A., Jr.; García-Cardeña, G. Endothelial cell dysfunction and the pathobiology of atherosclerosis. Circ. Res. 2016, 118, 620–636. [Google Scholar] [CrossRef]
- Tian, Q.; Leung, F.P.; Chen, F.M.; Tian, X.Y.; Chen, Z.; Tse, G.; Ma, S.; Wong, W.T. Butyrate protects endothelial function through PPARδ/miR-181b signaling. Pharmacol. Res. 2021, 169, 105681. [Google Scholar] [CrossRef]
- Roglic, G. WHO Global report on diabetes: A summary. Int. J. Noncommun. Dis. 2016, 1, 3–8. [Google Scholar] [CrossRef]
- DeFronzo, R.A.; Ferrannini, E.; Groop, L.; Henry, R.R.; Herman, W.H.; Holst, J.J.; Hu, F.B.; Kahn, C.R.; Raz, I.; Shulman, G.I.; et al. Type 2 diabetes mellitus. Nat. Rev. Dis. Primers 2015, 1, 15019. [Google Scholar] [CrossRef]
- Cheang, W.S.; Wong, W.T.; Tian, X.Y.; Yang, Q.; Lee, H.K.; He, G.-W.; Yao, X.; Huang, Y. Endothelial nitric oxide synthase enhancer reduces oxidative stress and restores endothelial function in db/db mice. Cardiovasc. Res. 2011, 92, 267–275. [Google Scholar] [CrossRef]
- Einarson, T.R.; Acs, A.; Ludwig, C.; Panton, U.H. Prevalence of cardiovascular disease in type 2 diabetes: A systematic literature review of scientific evidence from across the world in 2007–2017. Cardiovasc. Diabetol. 2018, 17, 83. [Google Scholar] [CrossRef]
- Turnbull, F.; Abraira, C.; Anderson, R.; Byington, R.; Chalmers, J.; Duckworth, W.; Evans, G.; Gerstein, H.; Holman, R.; Moritz, T. Intensive glucose control and macrovascular outcomes in type 2 diabetes. Diabetologia 2009, 52, 2288–2298. [Google Scholar] [CrossRef] [PubMed]
- Kelly, T.N.; Bazzano, L.A.; Fonseca, V.A.; Thethi, T.K.; Reynolds, K.; He, J. Systematic review: Glucose control and cardiovascular disease in type 2 diabetes. Ann. Intern. Med. 2009, 151, 394–403. [Google Scholar] [CrossRef] [PubMed]
- Giacco, F.; Brownlee, M. Oxidative stress and diabetic complications. Circ. Res. 2010, 107, 1058–1070. [Google Scholar] [CrossRef] [PubMed]
- Chook, C.Y.B.; Chen, F.M.; Tse, G.; Leung, F.P.; Wong, W.T. Crocodile blood supplementation protects vascular function in diabetic mice. Food Prod. Process. Nutr. 2021, 3, 19. [Google Scholar] [CrossRef]
- Chen, Z.; Ding, W.; Yang, X.; Lu, T.; Liu, Y. Isoliquiritigenin, a potential therapeutic agent for treatment of inflammation-associated diseases. J. Ethnopharmacol. 2024, 318, 117059. [Google Scholar] [CrossRef]
- Wang, K.-L.; Yu, Y.-C.; Hsia, S.-M. Perspectives on the Role of Isoliquiritigenin in Cancer. Cancers 2021, 13, 115. [Google Scholar] [CrossRef]
- Mamedov, N.A.; Egamberdieva, D. Phytochemical constituents and pharmacological effects of licorice: A review. Plant Hum. Health Pharmacol. Ther. Uses 2019, 3, 1–21. [Google Scholar]
- Peng, F.; Du, Q.; Peng, C.; Wang, N.; Tang, H.; Xie, X.; Shen, J.; Chen, J. A review: The pharmacology of isoliquiritigenin. Phytother. Res. 2015, 29, 969–977. [Google Scholar] [CrossRef]
- Gao, Y.; Lv, X.; Yang, H.; Peng, L.; Ci, X. Isoliquiritigenin exerts antioxidative and anti-inflammatory effects via activating the KEAP-1/Nrf2 pathway and inhibiting the NF-κB and NLRP3 pathways in carrageenan-induced pleurisy. Food Funct. 2020, 11, 2522–2534. [Google Scholar] [CrossRef]
- Honda, H.; Nagai, Y.; Matsunaga, T.; Okamoto, N.; Watanabe, Y.; Tsuneyama, K.; Hayashi, H.; Fujii, I.; Ikutani, M.; Hirai, Y. Isoliquiritigenin is a potent inhibitor of NLRP3 inflammasome activation and diet-induced adipose tissue inflammation. J. Leukoc. Biol. 2014, 96, 1087–1100. [Google Scholar] [CrossRef]
- Zhang, M.; Huang, L.-L.; Teng, C.-H.; Wu, F.-F.; Ge, L.-y.; Shi, Y.-J.; He, Z.-L.; Liu, L.; Jiang, C.-J.; Hou, R.-N. Isoliquiritigenin provides protection and attenuates oxidative stress-induced injuries via the Nrf2-ARE signaling pathway after traumatic brain injury. Neurochem. Res. 2018, 43, 2435–2445. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.-F.; Liu, J.; Yang, Y.-A.; Zhu, H.-L. A review: The anti-inflammatory, anticancer and antibacterial properties of four kinds of licorice flavonoids isolated from licorice. Curr. Med. Chem. 2020, 27, 1997–2011. [Google Scholar] [CrossRef] [PubMed]
- Yang, L.; Wang, D.; Zhang, Z.; Jiang, Y.; Liu, Y. Isoliquiritigenin alleviates diabetic symptoms via activating AMPK and inhibiting mTORC1 signaling in diet-induced diabetic mice. Phytomedicine 2022, 98, 153950. [Google Scholar] [CrossRef] [PubMed]
- Huang, X.; Shi, Y.; Chen, H.; Le, R.; Gong, X.; Xu, K.; Zhu, Q.; Shen, F.; Chen, Z.; Gu, X.; et al. Isoliquiritigenin prevents hyperglycemia-induced renal injuries by inhibiting inflammation and oxidative stress via SIRT1-dependent mechanism. Cell Death Dis. 2020, 11, 1040. [Google Scholar] [CrossRef] [PubMed]
- Gu, X.; Shi, Y.; Chen, X.; Sun, Z.; Luo, W.; Hu, X.; Jin, G.; You, S.; Qian, Y.; Wu, W.; et al. Isoliquiritigenin attenuates diabetic cardiomyopathy via inhibition of hyperglycemia-induced inflammatory response and oxidative stress. Phytomedicine 2020, 78, 153319. [Google Scholar] [CrossRef] [PubMed]
- Burke, S.J.; Batdorf, H.M.; Burk, D.H.; Noland, R.C.; Eder, A.E.; Boulos, M.S.; Karlstad, M.D.; Collier, J.J. db/db Mice Exhibit Features of Human Type 2 Diabetes That Are Not Present in Weight-Matched C57BL/6J Mice Fed a Western Diet. J. Diabetes Res. 2017, 2017, 8503754. [Google Scholar] [CrossRef] [PubMed]
- Faul, F.; Erdfelder, E.; Buchner, A.; Lang, A.-G. Statistical power analyses using G*Power 3.1: Tests for correlation and regression analyses. Behav. Res. Methods 2009, 41, 1149–1160. [Google Scholar] [CrossRef]
- Peiró, C.; Romacho, T.; Azcutia, V.; Villalobos, L.; Fernández, E.; Bolaños, J.P.; Moncada, S.; Sánchez-Ferrer, C.F. Inflammation, glucose, and vascular cell damage: The role of the pentose phosphate pathway. Cardiovasc. Diabetol. 2016, 15, 1–15. [Google Scholar] [CrossRef]
- Sprague, A.H.; Khalil, R.A. Inflammatory cytokines in vascular dysfunction and vascular disease. Biochem. Pharmacol. 2009, 78, 539–552. [Google Scholar] [CrossRef]
- Tabit, C.E.; Chung, W.B.; Hamburg, N.M.; Vita, J.A. Endothelial dysfunction in diabetes mellitus: Molecular mechanisms and clinical implications. Rev. Endocr. Metab. Disord. 2010, 11, 61–74. [Google Scholar] [CrossRef]
- Bendall, J.K.; Rinze, R.; Adlam, D.; Tatham, A.L.; de Bono, J.; Channon, K.M. Endothelial Nox2 overexpression potentiates vascular oxidative stress and hemodynamic response to angiotensin II: Studies in endothelial-targeted Nox2 transgenic mice. Circ. Res. 2007, 100, 1016–1025. [Google Scholar] [CrossRef] [PubMed]
- Herder, C.; Dalmas, E.; Böni-Schnetzler, M.; Donath, M.Y. The IL-1 pathway in type 2 diabetes and cardiovascular complications. Trends Endocrinol. Metab. 2015, 26, 551–563. [Google Scholar] [CrossRef]
- Peiró, C.; Lorenzo, Ó.; Carraro, R.; Sánchez-Ferrer, C.F. IL-1β inhibition in cardiovascular complications associated to diabetes mellitus. Front. Pharmacol. 2017, 8, 254176. [Google Scholar] [CrossRef]
- Xu, J.; Zou, M.-H. Molecular Insights and Therapeutic Targets for Diabetic Endothelial Dysfunction. Circulation 2009, 120, 1266–1286. [Google Scholar] [CrossRef]
- Takeda, Y.; Matoba, K.; Sekiguchi, K.; Nagai, Y.; Yokota, T.; Utsunomiya, K.; Nishimura, R. Endothelial Dysfunction in Diabetes. Biomedicines 2020, 8, 182. [Google Scholar] [CrossRef]
- Tian, X.; Wong, W.; Xu, A.; Chen, Z.; Lu, Y.; Liu, L.; Lee, V.; Lau, C.; Yao, X.; Huang, Y. Rosuvastatin improves endothelial function in db/db mice: Role of angiotensin II type 1 receptors and oxidative stress. Br. J. Pharmacol. 2011, 164, 598–606. [Google Scholar] [CrossRef]
- Spanier, G.; Xu, H.; Xia, N.; Tobias, S.; Deng, S.; Wojnowski, L.; Forstermann, U.; Li, H. Resveratrol reduces endothelial oxidative stress by modulating the gene expression of superoxide dismutase 1 (SOD1), glutathione peroxidase 1 (GPx1) and NADPH oxidase subunit (Nox4). J. Physiol. Pharmacol. 2009, 60, 111–116. [Google Scholar]
- Chen, B.; Lu, Y.; Chen, Y.; Cheng, J. The role of Nrf2 in oxidative stress-induced endothelial injuries. J. Endocrinol. 2015, 225, R83–R99. [Google Scholar] [CrossRef]
- Oguntibeju, O.O. Type 2 diabetes mellitus, oxidative stress and inflammation: Examining the links. Int. J. Physiol. Pathophysiol. Pharmacol. 2019, 11, 45–63. [Google Scholar]
- Chen, Q.; Hao, H.; Guo, Z.; Zuo, Y.; Cheng, C.K.; Zhang, C.-L.; Wang, L.; Lu, A.; Huang, Y.; He, L. Pien Tze Huang (PZH) protects endothelial function in diabetic mice. Life Sci. 2024, 349, 122723. [Google Scholar] [CrossRef]
- Li, H.; Zhang, Q. Research Progress of Flavonoids Regulating Endothelial Function. Pharmaceuticals 2023, 16, 1201. [Google Scholar] [CrossRef]
- Unnikrishnan, M.K.; Veerapur, V.; Nayak, Y.; Mudgal, P.P.; Mathew, G. Chapter 13—Antidiabetic, Antihyperlipidemic and Antioxidant Effects of the Flavonoids. In Polyphenols in Human Health and Disease; Watson, R.R., Preedy, V.R., Zibadi, S., Eds.; Academic Press: San Diego, CA, USA, 2014; pp. 143–161. [Google Scholar]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, L.; Zhu, R.; He, C.; Li, H.; Zhang, Q.; Cheung, Y.M.; Leung, F.P.; Wong, W.T. Licorice Extract Isoliquiritigenin Protects Endothelial Function in Type 2 Diabetic Mice. Nutrients 2024, 16, 3160. https://doi.org/10.3390/nu16183160
Wang L, Zhu R, He C, Li H, Zhang Q, Cheung YM, Leung FP, Wong WT. Licorice Extract Isoliquiritigenin Protects Endothelial Function in Type 2 Diabetic Mice. Nutrients. 2024; 16(18):3160. https://doi.org/10.3390/nu16183160
Chicago/Turabian StyleWang, Lin, Ruiwen Zhu, Chufeng He, Huixian Li, Qile Zhang, Yiu Ming Cheung, Fung Ping Leung, and Wing Tak Wong. 2024. "Licorice Extract Isoliquiritigenin Protects Endothelial Function in Type 2 Diabetic Mice" Nutrients 16, no. 18: 3160. https://doi.org/10.3390/nu16183160