1. Introduction
Mesothelioma has a long latency period, usually emerging 20–40 years after asbestos exposure [
1]. Malignant pleural mesothelioma (MPM) is rare (prevalence 1–9/100,000), but the corresponding annual death toll worldwide is still estimated at about 40,000 [
2,
3]. Each year, 125 million people are exposed to asbestos, according to a World Health Organization report [
4]. The International Agency for Research on Cancer confirmed that all fibrous forms of asbestos are carcinogenic to humans. The main outcome of exposure is mesothelioma, but cancer at other sites, such as respiratory-tract tumors, are moderately frequent [
4]. Previous in vitro studies have demonstrated the cytotoxic effects of asbestos fibers [
5,
6].
A significant association between MPM and asbestos exposure has been reported, showing a clear, increasing trend in the odds ratio (OR) with increasing cumulative exposure among subjects exposed to over 10 fiber/mL-years [
7]. Another study reported that the incidence of malignant mesothelioma (MM) was strongly associated with the proximity of one’s residence to an asbestos exposure source [
8].
DNA methylation (DNAm) is an epigenetic mechanism involved in gene expression regulation. In particular, dysregulation of promoter DNAm and histone modification are epigenetic mechanisms involved in human malignancies [
9].
According to recent papers, both DNAm and genetic alterations may contribute to MPM tumorigenesis [
10,
11,
12,
13,
14,
15]. Whereas the genome remains consistent throughout one’s lifetime, factors like ageing, lifestyle, environmental exposures and diseases can modify DNAm. The adaptive nature of DNAm means that it can be used to link environmental factors to the development of pathologic phenotypes such as cancers. Fasanelli et al. observed an association between exposure to tobacco and site-specific CpG methylation. They also used peripheral blood DNA to evaluate the importance of these epigenetic alterations in the aetiology of lung cancer [
16].
There is less information on the mechanisms and clinical outcomes of epigenetic derangements in MPM [
17,
18,
19]. Several studies have evaluated DNAm alterations in MM samples [
20,
21,
22], but few of them focused on DNAm alteration in blood as a circulating marker. Fischer et al. examined serum DNAm of nine gene-specific promoters from MM cases [
23]. A more recent paper identified hypomethylation of a single CpG in
FKBP5 in whole blood cells as a predictor of overall survival in MPM cases [
13]. Guarrera et al. evaluated methylation levels in DNA from whole blood leukocytes as potential diagnostic markers for MPM and found a differential methylation between asbestos-exposed MPM cases and controls, mainly in genes related to the immune system [
11]. The identification of reliable DNAm biomarkers with high sensitivity and specificity for MPM risk assessment would be a major advancement.
This study was undertaken with the goal to identify new biomarkers for MPM risk assessment and to determine if peripheral blood DNAm profiles have any predictive value. The second goal was to evaluate the interaction effect of asbestos exposure with DNAm on MPM risk. Currently, there are no sensitive testing methods available for the screening of asbestos-exposed individuals who are at high risk of developing MPM. Thus, the identification of reliable MPM diagnostic biomarkers in peripheral blood might provide a tool for detecting the disease at an early stage.
2. Results
2.1. Epigenome-Wide Association Study (EWAS)
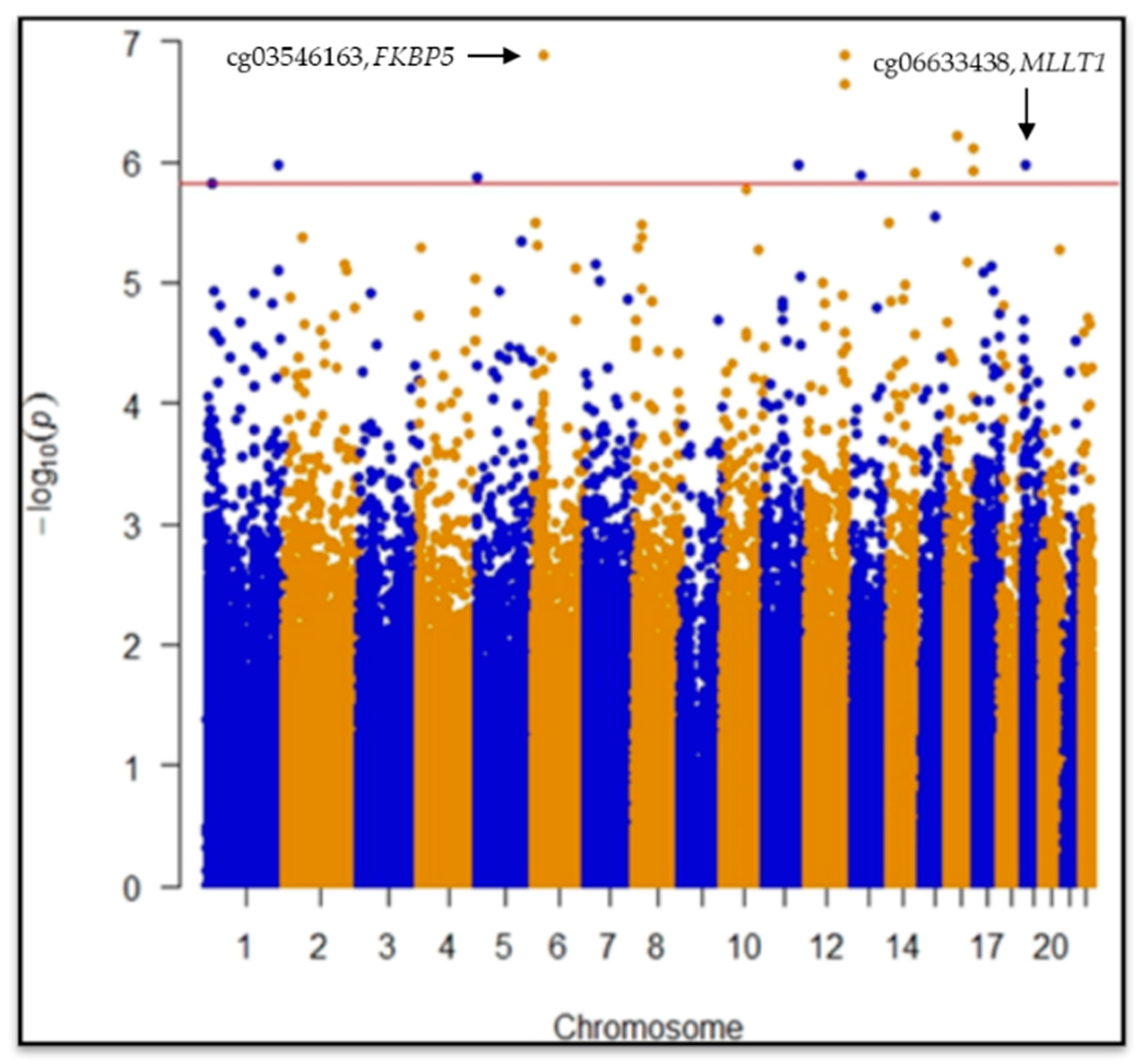
CpGs (445,254) passed quality control procedures and were considered for statistical analyses. EWAS revealed two statistically significant differentially methylated single-CpGs between case and control groups: cg03546163 in the
FKBP5 gene (Mean Difference in beta values (MD) = 0.09, 95% CI = −0.12|−0.06,
p = 1.2 × 10
−7,
p = 0.028) and cg06633438 in the
MLLT1 gene (MD = 0.07, 95% CI = 0.04|0.10,
p = 1.0 × 10
−6,
p = 0.049) after False Discovery Rate (FDR) post hoc correction (
Figure 1;
Table 1).
Another 10 CpGs showed hypo/hypermethylation in MPM considering FDR < 0.05 but not effect size (MD) cut off ≥ |0.05| (
Table 1).
Bootstrap was computed to estimate measures of accuracy using random sampling methods. The bias-corrected and accelerated (BCa) bootstrap interval was calculated for cg03546163 in FKBP5 (95% CIBCa = −0.16|−0.10, z0 = −0.008, a = 0.002) and cg06633438 in MLLT1 (95% CIBCa = −0.06|−0.1, z0 = −0.011, a = 0.0004) genes, confirming the robustness of the results considering the sample under study.
Statistically significant differences in MD between cases and controls were found in the WBCs estimated (monocytes, p = 6.0 × 10−3; granulocytes, p = 2.2 × 10−16; B cells, p = 1.1 × 10−12; NK cells, p = 3.6 × 10−4; CD4+ T, p = 2.2 × 10−16; CD8+ T, p = 6.8 × 10−11; Naïve CD4T, p = 0.012; Naïve CD8T, p = 7.0 × 10−3).
In order to assess if smoking status, classified as current, former and never-smokers, could modify DNAm profiles, we performed a multivariate regression analysis with the same model used for the discovery phase. No evidence of methylation differences linked to different smoking levels was found for any of the twelve statistically significant CpGs.
2.2. Receiver Operation Characteristics (ROC) for Case-Control Discrimination
The baseline model (BM) including age, gender, asbestos exposure and WBCs was compared with BM adding the DNAm levels of cg03546163 or cg06633438. Receiver Operation Characteristics 8ROC9 curves showed a significant increase in MPM discrimination when DNAm information was added in the model (
Table 2).
2.3. Interaction Analysis
CpG sites and asbestos exposure were considered as predictors of MPM risk in the interaction model. Categorical variables (quantile information) were used considering median values.
We tested the interaction between asbestos exposure and DNAm levels at cg03546163 in FKBP5 and cg06633438 in MLLT1.
Considering cg03546163 in
FKBP5, DNA hypermethylation and low asbestos exposure levels were used as references, while for cg06633438 in
MLLT1, DNA hypomethylation and low asbestos exposure levels were set as references (
Table 3).
The OR was estimated as the relationship between the combination of single-CpGs DNAm levels and asbestos exposure quantile, and the reference (low median asbestos exposure and hypermethylation status for cg03546163, or hypomethylation status for cg06633438). Age, gender, population stratification, and WBCs were included in the GLM (family = binomial) to adjust the interaction effect.
The relationship between asbestos exposures and single-CpG DNAm levels was evaluated. An increase of one unit of asbestos exposure (rank transformed fibers/mL years) was related to the FKBP5 gene (β = −0.016, 95% CI = −0.031|−0.001, p = 0.044) and MLLT1 gene (β = −0.014, 95% CI = 0.001|0.026, p = 0.035) methylation level variations.
Strong association between asbestos exposure and MPM risk, considering dichotomous distribution of asbestos exposure, was found (OR = 6.11, 95% CI = 3.73|10.20, p = 1.8 × 10−12). Quartile distribution of asbestos exposure was evaluated to estimate the potential incremental association with MPM risk (1st quartile: used as reference; 2nd quartile: OR = 1.83, 95% CI = 0.93|3.69, p = 0.09; 3rd quartile: OR = 6.63, 95% CI = 3.30|13.81, p = 2.1 × 10−7; 4th quartile: OR = 11.00, 95% CI = 5.26|24.30, p = 7.3 × 10−10).
2.4. Validation and Replication
For the replication and validation approaches, an independent sample of 140 MPM cases and 104 cancer-free asbestos-exposed controls from the same areas were considered, using a targeted DNAm analysis technique.
The direction and magnitude of the association was consistent for cg03546163 and cg06633438 DNAm. Patients showed significantly lower DNAm at cg03546163 (MD = −0.061, 95% CI = −0.087|−0.036, p = 4.5 × 10−6) and higher DNAm at cg06633438 (MD = 0.024, 95% CI = 0.061|0.013, p = 4.0 × 10−2) compared with controls. A multivariate analysis confirmed that DNAm at cg03546163 in FKBP5 and cg06633438 in MLLT1 were independently associated with MPM detection.
3. Discussion
In the present study, we used a whole genome microarray approach to investigate DNAm in WBCs from MPM cases and asbestos-exposed cancer-free controls from a region with a history of asbestos exposure (Piedmont, Italy) [
10] in order to identify new noninvasive epigenetic markers related to MPM. The identification of reliable MPM diagnostic biomarkers in peripheral blood might improve risk assessment.
We observed hypomethylation of CpG cg03546163, located in the 5′ UTR region of FKBP5 gene, in MPM cases compared to controls.
Epigenetic activation of the FKBP Prolyl Isomerase 5 (
FKBP5) gene has been shown to be associated with increased stress sensitivity and the risk of psychiatric disorders [
24].
FKBP5 is an immunophilin and has an important role in immunoregulation and in protein folding and trafficking. It plays a role in transcriptional complexes and acts as a cotranscription factor, along with other proteins in the
FKBP family [
25]. The suggestion of a possible role of
FKBP5 in the development and progression of different types of cancer has stemmed from several studies. In particular, high protein expression has been linked to either suppression or promotion of tumour growth, depending on tumour type and microenvironment [
26,
27].
FKBP5 is involved in the NF-kB and AKT signaling pathways, both of which are implicated in tumorigenesis [
28]. Notably, NF-kB appears to be frequently constitutively activated in malignant tumours and involved in the modulation of genes linked to cell motility, neoangiogenesis, proliferation and programmed cell death [
29]. An epigenetic upregulation of
FKBP5 could promote NF-kB activation [
30]. STAT3-NFkB activity is involved in chemoresistance in MM cells [
31], and NFkB was shown to be constitutively active as a result of asbestos-induced chronic inflammation [
32].
CpG cg06633438 located in the body region of the MLLT1 gene was hypermethylated in cases compared to controls.
The
MLLT1 gene encodes the ENL protein, a histone acetylation reader component of the super elongation complex (SEC), which promotes transcription at the elongation stage by suppressing transient pausing by the polymerase at multiple sites along the DNA. In acute myeloid leukemia,
MLLT1 regulates chromatin remodeling and gene expression of many important proto-oncogenes [
31]. Yoshikawa and colleagues suggested that mesothelioma may be the consequence of the somatic inactivation of chromatin-remodeling complexes and/or histone modifiers, including
MLLT1 [
30].
In mesothelioma patients with short-term recurrence after surgery, frequent 19p13.2 loss was reported. This region encompasses several putative tumor suppressors or oncogenes, including
MLLT1 [
32].
Interestingly,
MLLT1 and
FKBP5 showed opposite behavior, increasing and decreasing DNAm levels, respectively, in relation to MPM. This finding could reflect the opposite expression profiles of the two genes among all the different subtypes of white blood cells in normal human hematopoiesis, as reported in the Blood Spot database (
http://servers.binf.ku.dk/bloodspot/, accessed on 26 May 2021) (
Figure 2) [
33].
Our interaction analysis showed that considering DNAm levels at FKBP5 and MLLT1 genes together with asbestos exposure levels may help to better define MPM risk for asbestos-exposed subjects.
Six single-CpGs showed differential methylation in patients, including those located in
C13orf30, CDC42BPB, ZNF629 and
ALG9 genes; the other six were not annotated to named genes.
ALG9 is a glycogene whose reduced expression has been described during the epithelial-to-mesenchymal transition, an essential process also involved in cancer progression [
34]. The
CDC42BPB gene is ubiquitously expressed in mammals and encodes a serine/threonine protein kinase, a member of the MRCK family [
35]. The role of MRCKs in cytoskeletal reorganization during cell migration and invasion has been characterized [
36]. The biological function of
C13orf30 and
ZNF629, a DNA-binding transcription factor, is still to be established.
MPM cases and asbestos-exposed controls showed different proportions of estimated WBCs, which may denote the crucial implication of the immune system. It is known that in cancer, including mesothelioma, the immune system is affected [
37], and there is evidence that asbestos directs antigen overstimulation, and that reactive oxygen species production induces functional changes in WBCs [
38]. Indeed, in MPM cases, we showed a reduction of estimated CD4+ and CD8+ T lymphocytes, suggesting a weaker adaptive immune system [
39]. This may reflect the possible occurrence of functional changes in WBC subtypes in MPM [
40,
41].
The need for reliable biomarkers is of extreme relevance for a disease such as MPM, which is characterized by the accumulation and persistence of asbestos fibers in the lungs, leading to a long latency period before clear clinical signs of the tumor are detectable. Several biomarkers for early MPM detection (e.g., mesothelin, osteopontin and fibulin-3) have been proposed so far; however, some of them are still under investigation [
42]. In this context, DNAm changes in easily-accessible WBCs may provide a useful tool to better assess MPM risk in asbestos-exposed subjects.
Our findings that DNAm levels in single-CpGs in FKBP5 and MLLT1 genes are independent markers of MPM in asbestos-exposed subjects suggest the potential use of blood DNAm analysis as a noninvasive test for MPM detection.
Some somatic gene alterations in lung cancer have been linked to tobacco smoke, but few data are available on the role of asbestos fibers: Andujar and colleagues investigate the mechanism of P16/CDKN2A alterations in lung cancer including asbestos-exposed patients. P16/CDKN2A gene inactivation in asbestos-exposed non-small-cell lung carcinoma (NSCLC) cases, a tumor independent of tobacco smoking but associated with asbestos exposure, mainly occurs via promoter hypermethylation, loss of heterozygosity and homozygous deletion, suggesting a possible relationship with an effect of asbestos fibers [
43].We observed epigenetic deregulations in the blood of MPM patients compared to that of cancer-free controls, suggesting the potential use of DNAm for risk stratification among asbestos-exposed individuals.
If this observation can be verified in prospectively collected samples, it may be possible to use CpGs methylation to further improve MPM risk estimation for subjects with occupational and/or environmental asbestos exposure.
Limitation of the Study
Leukocyte DNAm may be a nonspecific marker related to a general, tumour-induced inflammatory status rather than a specific MPM biomarker. Further studies are therefore needed to support our findings.
One main limitation of the functional interpretation of our results is that all our cases had already developed MPM at recruitment: thus, our findings likely reflect disease status rather than being markers of the dynamic processes leading to MPM onset. The lack of MPM tissue from the same subjects also poses major constraints to the functional interpretation of our findings.
Notwithstanding the above limitations, the discrimination between MPM cases and asbestos-exposed cancer-free controls improved when DNAm levels were considered together with asbestos exposure levels.


 ,
, 





{kind=link}
{kind=link}