
In Vitro Screening and Field Performance of EMS-Treated Eggplants for the Selection of Shoot and Fruit Borer-Resistant Plants

 , , , , and
, , , , and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
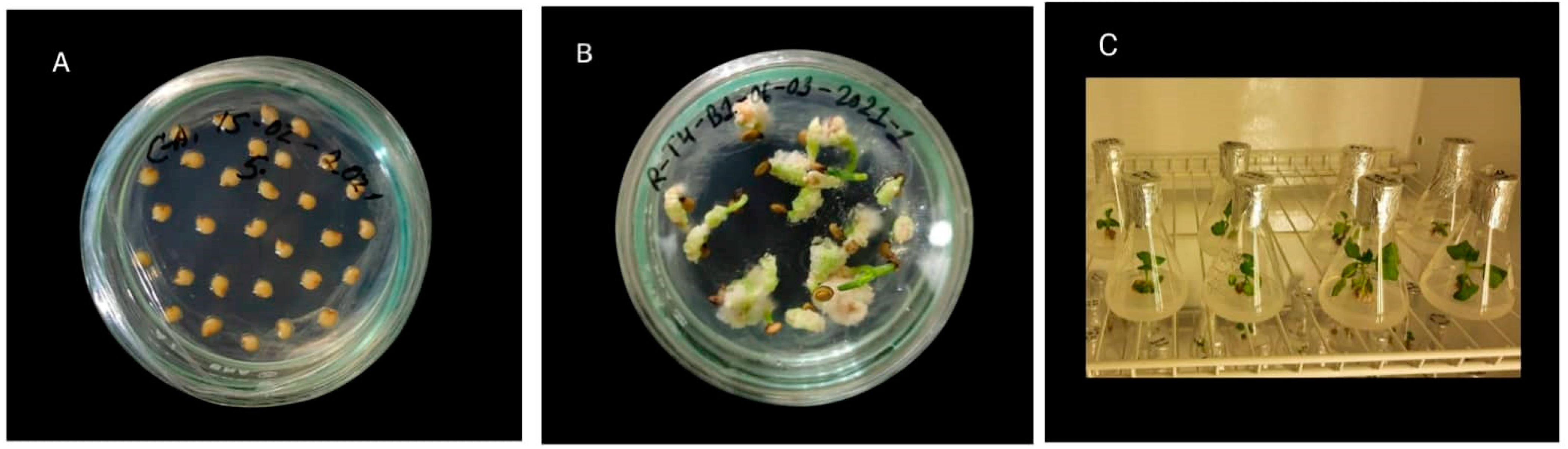
2.1. Plant Material and In Vitro Mutagenesis
2.2. Biochemical Analysis
2.2.1. Total Phenol Determination
2.2.2. Antioxidant Determination
2.2.3. Chlorophyll Determination
2.2.4. Solasodine Measurement
2.2.5. Total Sugar Content
2.2.6. Estimation of Minerals
2.3. Data Collection and Analysis
3. Results
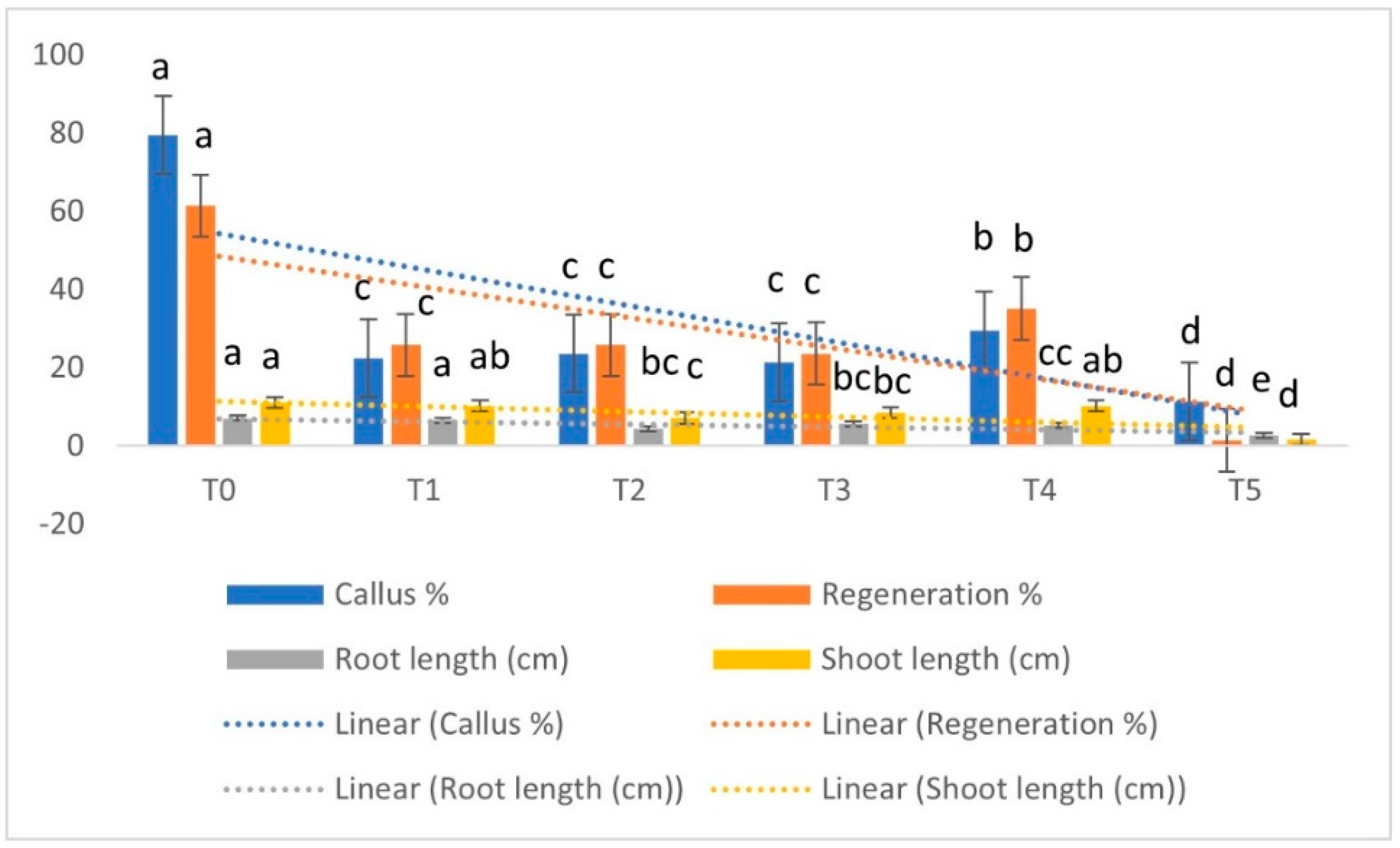
3.1. Effect of EMS on In Vitro Performance
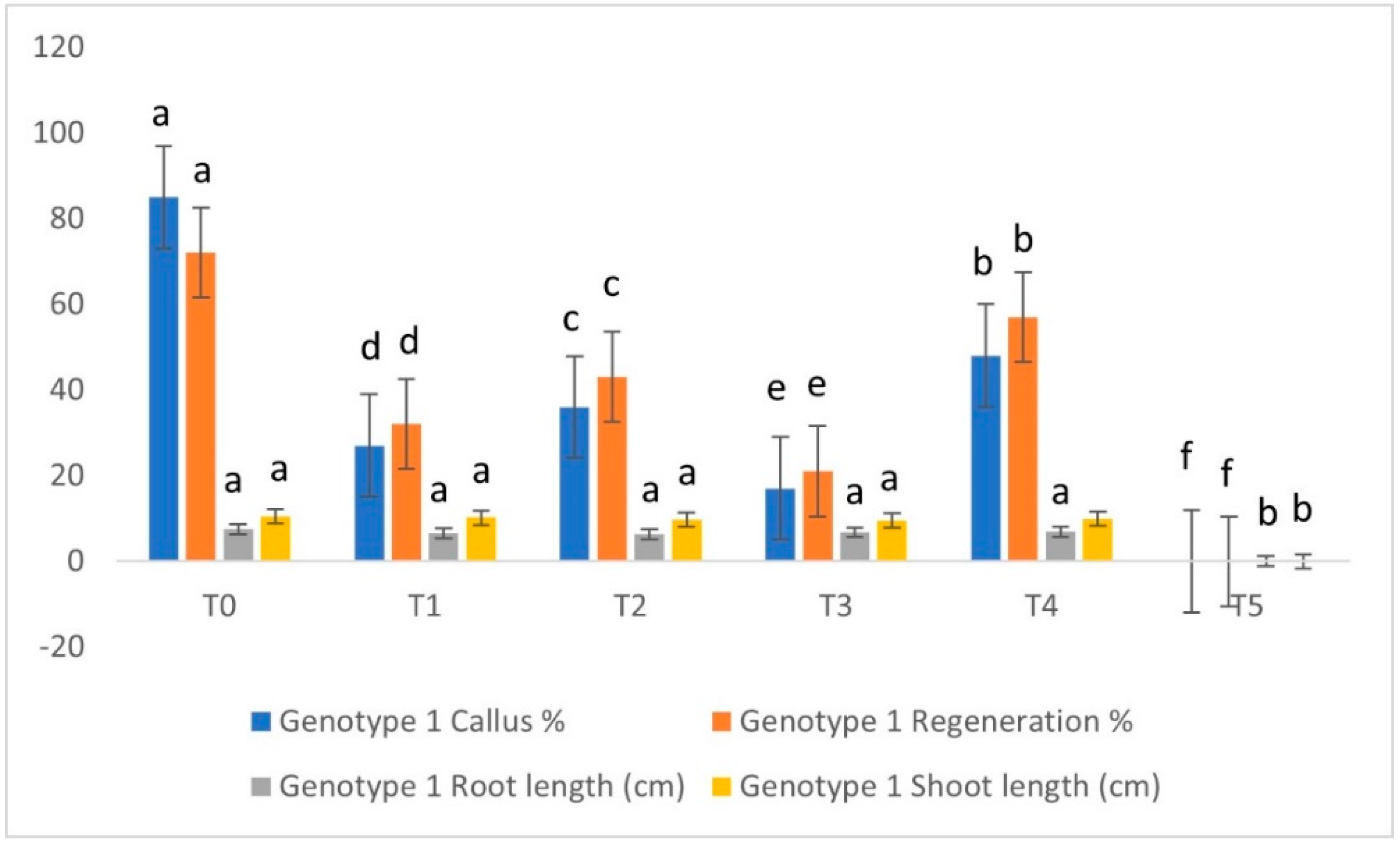
3.2. Treatment-Wise Genotypic Performance
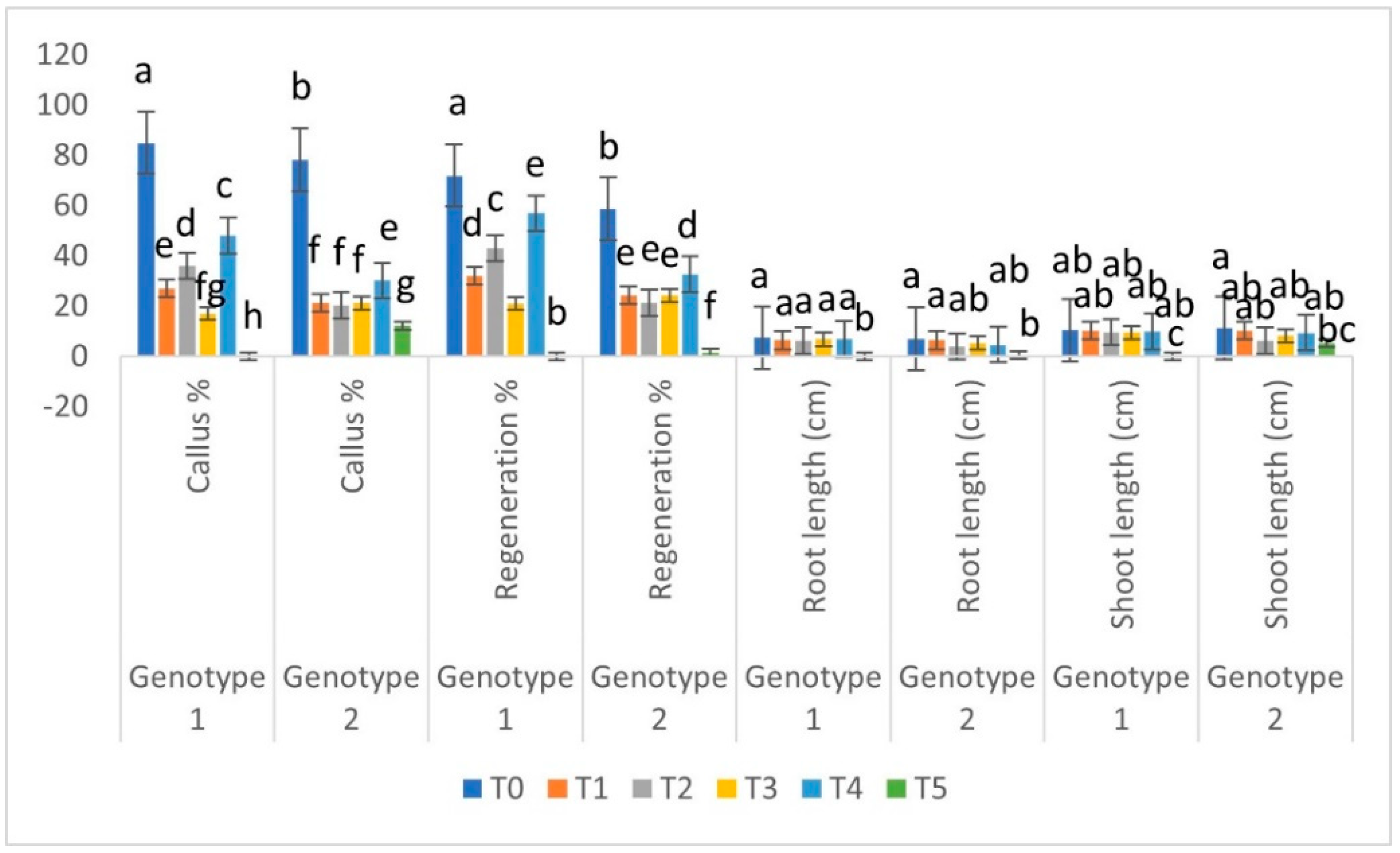
3.3. Genotypic Comparison
3.4. Field Performance of Regenerated Plantlets
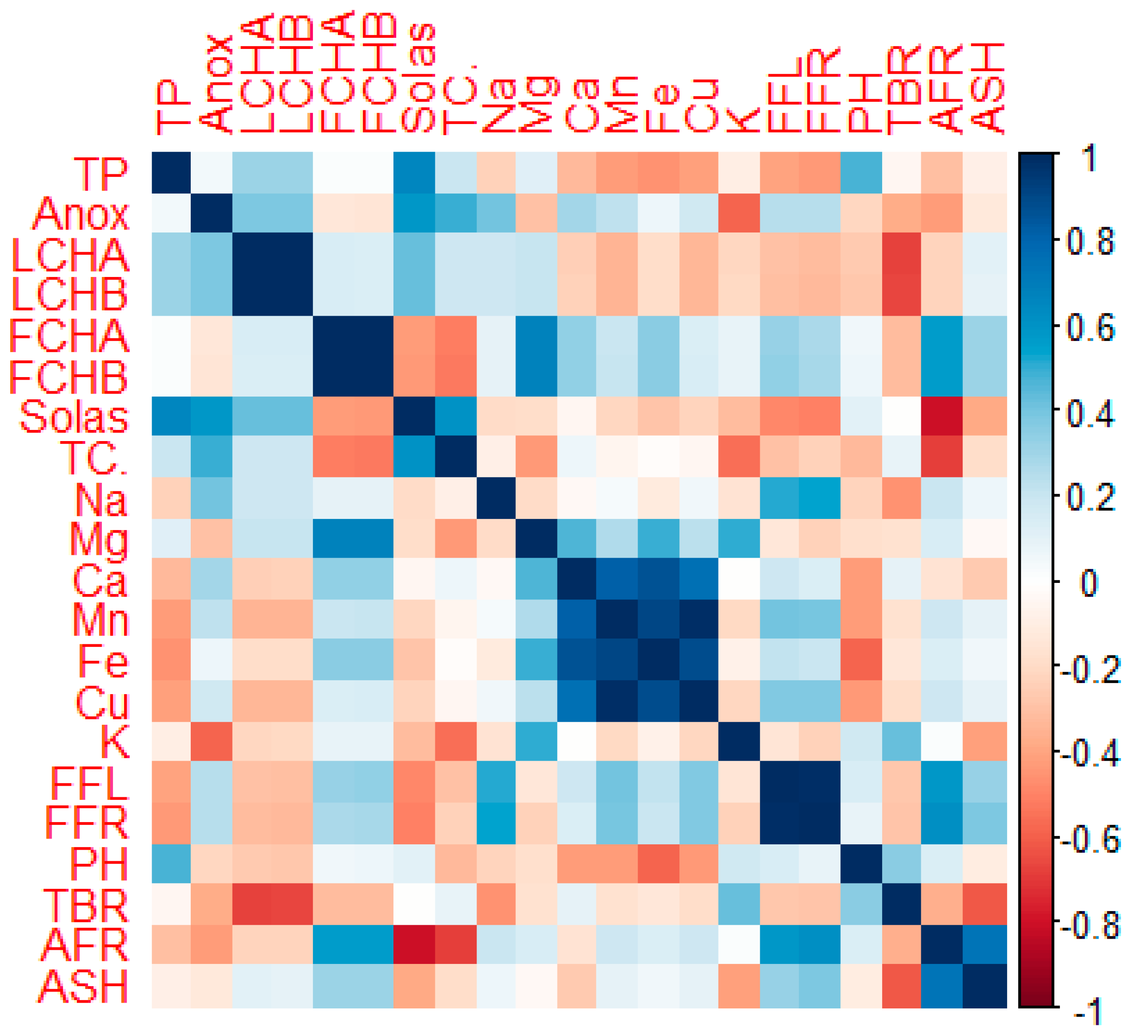
3.4.1. Correlation of Phenotypic and Biochemical Traits
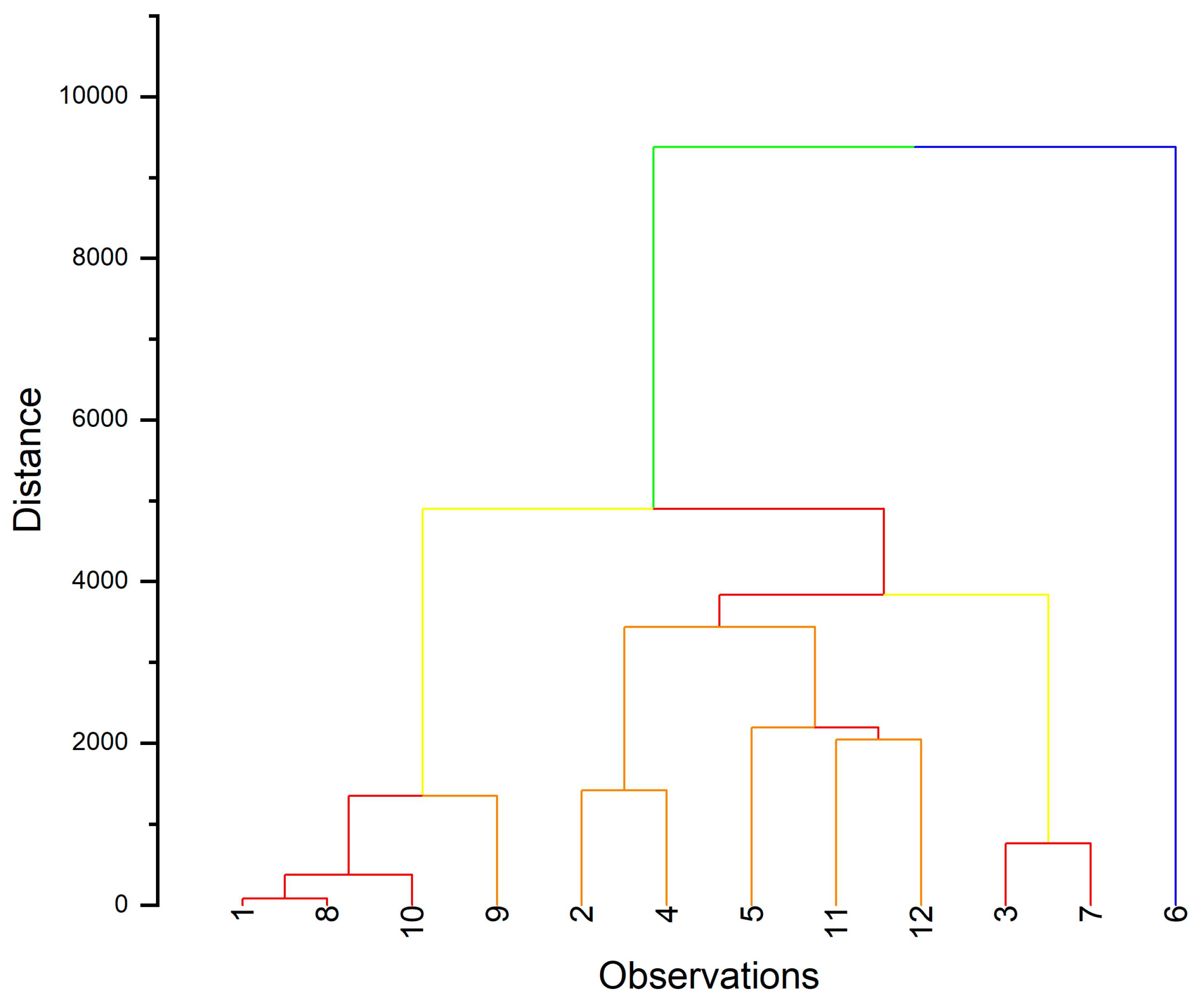
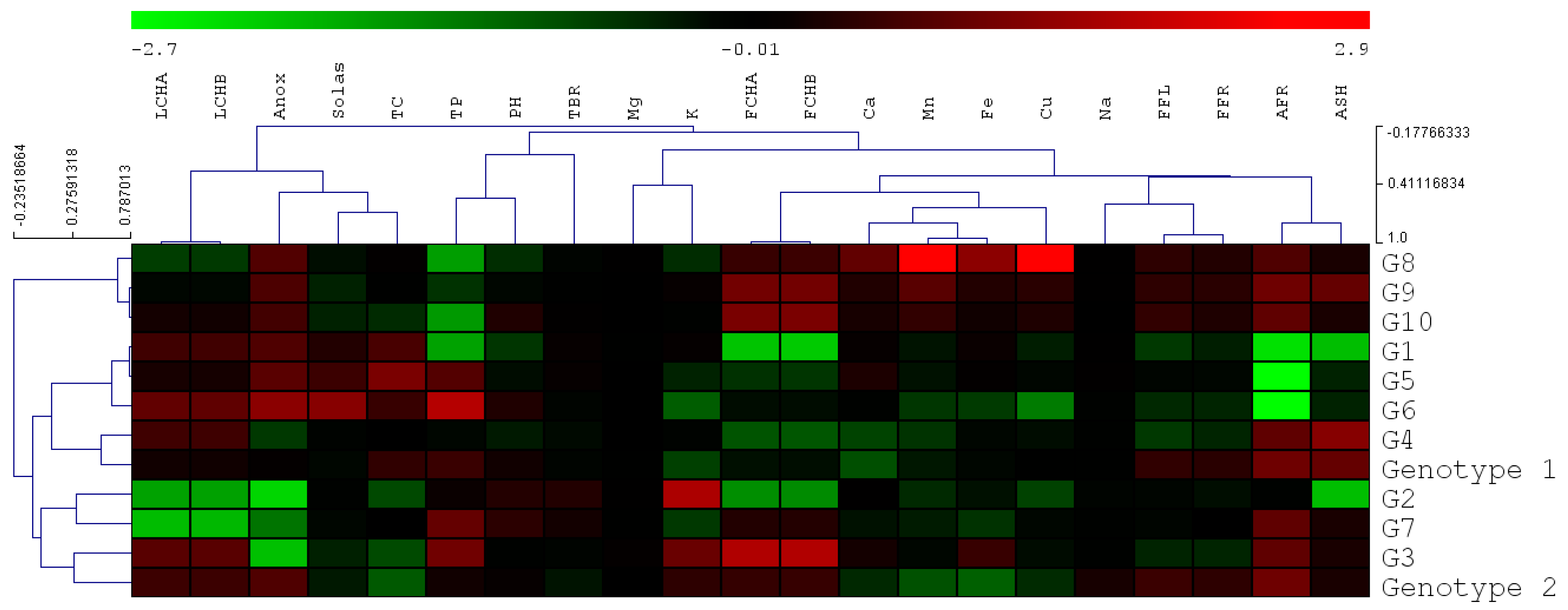
3.4.2. Classification of Mutants
4. Discussion
4.1. Optimization of EMS Treatment Methods
4.2. Field Performance of Regenerated Plants
Screening for Eggplant Shoot and Fruit Borer Resistance
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- FAO. FAOSTAT Food and Agriculture. 2019. Available online: http://www.fao.org/faostat (accessed on 28 June 2022).
- Boulekbache-Makhlouf, L.; Medouni, L.; Medouni-Adrar, S.; Arkoub, L.; Madani, K. Effect of solvents extraction on phenolic content and antioxidant activity of the byproduct of eggplant. Ind. Crop. Prod. 2013, 49, 668–674. [Google Scholar] [CrossRef]
- Prohens, J.; Whitaker, B.; Plazas, M.; Vilanova, S.; Hurtado, M.; Blasco, M.; Gramazio, P.; Stommel, J. Genetic diversity in morphological characters and phenolic acids content resulting from an interspecific cross between eggplant, Solanum melongena, and its wild ancestor (S. incanum). Ann. Appl. Biol. 2013, 162, 242–257. [Google Scholar] [CrossRef]
- Gürbüz, N.; Uluişik, S.; Frary, A.; Frary, A.; Doğanlar, S. Health benefits and bioactive compounds of eggplant. Food Chem. 2018, 268, 602–610. [Google Scholar] [CrossRef] [PubMed]
- Sharma, M.; Kaushik, P. Biochemical Composition of Eggplant Fruits: A Review. Appl. Sci. 2021, 11, 7078. [Google Scholar] [CrossRef]
- Okmen, B.; Sigva, H.O.; Mutlu, S.; Doganlar, S.; Yemenicioglu, A.; Frary, A. Total Antioxidant Activity and Total Phenolic Contents in Different Turkish Eggplant (Solanum Melongena L.) Cultivars. Int. J. Food Prop. 2009, 12, 616–624. [Google Scholar] [CrossRef] [Green Version]
- Kudlu, C.; Stone, G.D. The trials of genetically modified food: Bt eggplant and Ayurvedic medicine in India. Food Cult. Soc. 2013, 16, 21–42. [Google Scholar] [CrossRef]
- Taghipour, F.; Jamshidi, S.; Huyop, F. High performant eggplant in vitro regeneration and organogenesis. Agroecol. J. 2015, 11, 24–29. [Google Scholar]
- Friedman, M. Chemistry and Anticarcinogenic Mechanisms of Glycoalkaloids Produced by Eggplants, Potatoes, and Tomatoes. J. Agric. Food Chem. 2015, 63, 3323–3337. [Google Scholar] [CrossRef]
- Lelario, F.; De Maria, S.; Rivelli, A.R.; Russo, D.; Milella, L.; Bufo, S.A.; Scrano, L. A Complete Survey of Glycoalkaloids Using LC-FTICR-MS and IRMPD in a Commercial Variety and a Local Landrace of Eggplant (Solanum melongena L.) and their Anticholinesterase and Antioxidant Activities. Toxins 2019, 11, 230. [Google Scholar] [CrossRef] [Green Version]
- Rotino, G.L.; Sala, T.; Toppino, L. Eggplant. In Alien Gene Transfer in Crop Plants; Springer: New York, NY, USA, 2014; Volume 2, pp. 381–409. [Google Scholar]
- Sánchez-Mata, M.C.; Yokoyama, W.E.; Hong, Y.J.; Prohens, J. α-Solasonine and α-solamargine contents of gboma (Solanum macrocarpon L.) and scarlet (Solanum aethiopicum L.) eggplants. J. Agric. Food Chem. 2010, 58, 5502–5508. [Google Scholar] [CrossRef]
- Bagheri, M.; Bushehri, A.A.S.; Hassandokht, M.R.; Naghavi, M.R. Evaluation of Solasonine Content and Expression Patterns of SGT1 Gene in Different Tissues of Two Iranian Eggplant (Solanum melongena L.) Genotypes. Food Technol. Biotechnol. 2017, 55, 236–242. [Google Scholar] [CrossRef] [PubMed]
- Villano, C.; D’Amelia, V.; Esposito, S.; Adelfi, M.G.; Contaldi, F.; Ferracane, R.; Vitaglione, P.; Aversano, R.; Carputo, D. Genome-Wide HMG Family Investigation and Its Role in Glycoalkaloid Accumulation in Wild Tuber-Bearing Solanum commersonii. Life 2020, 10, 37. [Google Scholar] [CrossRef] [Green Version]
- Zhao, D.K.; Zhao, Y.; Chen, S.Y.; Kennelly, E.J. Solanum steroidal glycoalkaloids: Structural diversity, biological activities, and biosynthesis. Nat. Prod. Rep. 2021, 38, 1423–1444. [Google Scholar] [CrossRef] [PubMed]
- Alam, I.; Salimullah, M. Genetic engineering of eggplant (Solanum melongena L.): Progress, controversy and potential. Horticulturae 2021, 7, 78. [Google Scholar] [CrossRef]
- Collonnier, C.; Fock, I.; Kashyap, V.; Rotino, G.; Daunay, M.; Lian, Y.; Mariska, I.; Rajam, M.; Servaes, A.; Ducreux, G.; et al. Applications of biotechnology in eggplant. Plant. Cell Tissue Organ. Cult. 2001, 65, 91–107. [Google Scholar] [CrossRef]
- Hannachi, S.; Werbrouck, S.; Bahrini, I.; Abdelgadir, A.; Siddiqui, H.A.; Van Labeke, M.C. Obtaining Salt Stress-Tolerant Eggplant Somaclonal Variants from In Vitro Selection. Plants 2021, 10, 2539. [Google Scholar] [CrossRef]
- Krishna, H.; Alizadeh, M.; Singh, D.; Singh, U.; Chauhan, N.; Eftekhari, M.; Sadh, R.K. Somaclonal variations and their applications in horticultural crops improvement. 3 Biotech 2016, 6, 1–18. [Google Scholar] [CrossRef] [Green Version]
- Bado, S.; Forster, B.P.; Nielen, S.; Ali, A.M.; Lagoda, P.J.; Till, B.J.; Laimer, M. Plant mutation breeding: Current progress and future assessment. Plant. Breed. Rev. 2015, 39, 23–88. [Google Scholar]
- Chaudhary, J.; Alisha, A.; Bhatt, V.; Chandanshive, S.; Kumar, N.; Mir, Z.; Kumar, A.; Yadav, S.K.; Shivaraj, S.M.; Sonah, H.; et al. Mutation Breeding in Tomato: Advances, Applicability and Challenges. Plants 2019, 8, 128. [Google Scholar] [CrossRef] [Green Version]
- Gillmor, C.S.; Lukowitz, W. EMS Mutagenesis of Arabidopsis Seeds. Methods Mol. Biol. 2020, 2122, 15–23. [Google Scholar]
- Siddique, M.I.; Back, S.; Lee, J.-H.; Jo, J.; Jang, S.; Han, K.; Venkatesh, J.; Kwon, J.-K.; Jo, Y.D.; Kang, B.-C. Development and Characterization of an Ethyl Methane Sulfonate (EMS) Induced Mutant Population in Capsicum annuum L. Plants 2020, 9, 396. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Prodhan, M.Z.H.; Hasan, M.T.; Chowdhury, M.M.I.; Alam, M.S.; Rahman, M.L.; Azad, A.K.; Hossain, M.J.; Naranjo, S.E.; Shelton, A.M. Bt eggplant (Solanum melongena L.) in Bangladesh: Fruit production and control of eggplant fruit and shoot borer (Leucinodes orbonalis Guenee), effects on non-target arthropods and economic returns. PLoS ONE 2018, 13, e0205713. [Google Scholar] [CrossRef] [PubMed]
- Jahan, T.; Ali, M.A.; Raihan, M.S.; Rahman, M.M.; Abdullah, H.M.; Huda, N.; Hasan, M. Indigenous Mota-named coarse rice germplasm is distinct from fine-grained rice collected from south-central coastal Bangladesh as compared with the morphological descriptors and molecular markers. Braz. J. Bot. 2020, 43, 933–945. [Google Scholar] [CrossRef]
- Kashtwari, M.; Wani, A.A.; Dhar, M.K.; Jan, S.; Kamili, A.N. Development of an efficient in vitro mutagenesis protocol for genetic improvement of saffron (Crocus sativus L.). Physiol. Mol. Biol. Plants 2018, 24, 951–962. [Google Scholar] [CrossRef] [PubMed]
- Monika, Y.; Vinita, R.; Saharan, R.S.; Sehrawat, S.K. Influence of gamma radiations and EMS on morphological characteristics of gladiolus cv. PINK BEAUTY. AJH 2016, 11, 114–118. [Google Scholar]
- Arisha, M.H.; Shah, S.N.M.; Gong, Z.-H.; Jing, H.; Li, C.; Zhang, H.-X. Ethyl methane sulfonate induced mutations in M2 generation and physiological variations in M1 generation of peppers (Capsicum annuum L.). Front. Plant. Sci. 2015, 6, 399. [Google Scholar] [CrossRef] [Green Version]
- Ebrahimie, E.; Habashy, A.A.; Mohammadie-Dehcheshmeh, M.; Ghannadha, M.R.; Ghareyazie, B.; Yazdi-Amadi, B. Direct shoot regeneration from mature embryo as a rapid and genotype-independent pathway in tissue culture of heterogeneous diverse sets of cumin (Cuminum cyminum L.) genotypes. In Vitr. Cell. Dev. Biol. Plant 2006, 42, 455–460. [Google Scholar] [CrossRef]
- Ke, C.; Guan, W.; Bu, S.; Li, X.; Deng, Y.; Wei, Z.; Wu, W.; Zheng, Y. Determination of absorption dose in chemical mutagenesis in plants. PLoS ONE 2019, 14, e0210596. [Google Scholar] [CrossRef] [Green Version]
- Jankowicz-Cieslak, J.; Till, B.J. Chemical Mutagenesis of Seed and Vegetatively Propagated Plants Using EMS. Curr. Protoc. Plant. Biol. 2016, 1, 617–635. [Google Scholar] [CrossRef]
- Thangwana, A.; Gwata, E.T.; Zhou, M.M. Impact of chemical mutagenesis using ethyl methane sulphonate on tepary bean seedling vigour and adult plant performance. Heliyon 2021, 7, e06103. [Google Scholar] [CrossRef]
- Raina, A.; Laskar, R.A.; Wani, M.R.; Jan, B.L.; Ali, S.; Khan, S. Comparative Mutagenic Effectiveness and Efficiency of Gamma Rays and Sodium Azide in Inducing Chlorophyll and Morphological Mutants of Cowpea. Plants 2022, 11, 1322. [Google Scholar] [CrossRef] [PubMed]
- Batista, D.S.; Felipe, S.H.S.; Silva, T.D.; de Castro, K.M.; Mamedes-Rodrigues, T.C.; Miranda, N.A.; Ríos-Ríos, A.M.; Faria, D.V.; Fortini, E.A.; Chagas, K.; et al. Light quality in plant tissue culture: Does it matter? In Vitr. Cell. Dev. Biol. Plant 2018, 54, 195–215. [Google Scholar] [CrossRef]
- Monica, S.; Seetharaman, N. Physical and chemical mutagenic effect on pollen fertility in M1 generation of Garden Bean [Lablab purpureus (L.) Sweet var. typicus cv. CO (Gb) 14]. Eur. J. Exp. Biol. 2015, 5, 6–9. [Google Scholar]
- Kariyanna, B.; Prabhuraj, A.; Asokan, R.; Agrawal, A.; Gracy, R.G.; Jyoti, P.; Venkatesan, T.; Bheemanna, M.; Kalmath, B.; Diwan, J.R.; et al. Genome Mining and Expression Analysis of Carboxylesterase and Glutathione S-Transferase Genes Involved in Insecticide Resistance in Eggplant Shoot and Fruit Borer, Leucinodes orbonalis (Lepidoptera: Crambidae). Front. Physiol. 2020, 11, 594845. [Google Scholar] [CrossRef]
- Martínez-Ispizua, E.; Calatayud, Á.; Marsal, J.I.; Mateos-Fernández, R.; Díez, M.J.; Soler, S.; Valcárcel, J.V.; Martínez-Cuenca, M.-R. Phenotyping Local Eggplant Varieties: Commitment to Biodiversity and Nutritional Quality Preservation. Front. Plant. Sci. 2021, 12, 696272. [Google Scholar] [CrossRef] [PubMed]
- Kwon, Y.-I.; Apostolidis, E.; Shetty, K. In vitro studies of eggplant (Solanum melongena) phenolics as inhibitors of key enzymes relevant for type 2 diabetes and hypertension. Bioresour. Technol. 2008, 99, 2981–2988. [Google Scholar] [CrossRef] [PubMed]
- Plazas, M.; López-Gresa, M.P.; Vilanova, S.; Torres, C.; Hurtado, M.; Gramazio, P.; Andújar, I.; Herráiz, F.J.; Bellés, J.M.; Prohens, J. Diversity and Relationships in Key Traits for Functional and Apparent Quality in a Collection of Eggplant: Fruit Phenolics Content, Antioxidant Activity, Polyphenol Oxidase Activity, and Browning. J. Agric. Food Chem. 2013, 61, 8871–8879. [Google Scholar] [CrossRef]
- Lingampally, V.; Solanki, V.R.; Anuradha, D.L.; Sabita Raja, S. Effect of solasodine against last instar larvae of Tribolium confusum. J. Entomol. Zool. Stud. 2014, 2, 118–120. [Google Scholar]
- Bhalsing, S.R.; Teli, N.P.; Pawar, P.K.; Maheshwari, V.L. Isolation and characterization of solasodine from cultured cells of Solanum khasianum. J. Plant. Biol. 2000, 27, 185–187. [Google Scholar]
- Dar, S.A.; Wani, A.R.; Rather, B.A.; Parey, S.H.; Kandoo, A. Biochemical basis of resistance in brinjal genotypes against shoot and fruit borer (Leucinodes Orbonalis, Guenee). Chem. Sci. Rev. Lett. 2017, 6, 1062–1073. [Google Scholar]


Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Islam, M.A.; Bin Mohi Uddin, M.M.; Rasul, M.G.; Haque Swapon, M.A.; Ahmed, M.; Hasan, M. In Vitro Screening and Field Performance of EMS-Treated Eggplants for the Selection of Shoot and Fruit Borer-Resistant Plants. Agronomy 2022, 12, 1832. https://doi.org/10.3390/agronomy12081832
Islam MA, Bin Mohi Uddin MM, Rasul MG, Haque Swapon MA, Ahmed M, Hasan M. In Vitro Screening and Field Performance of EMS-Treated Eggplants for the Selection of Shoot and Fruit Borer-Resistant Plants. Agronomy. 2022; 12(8):1832. https://doi.org/10.3390/agronomy12081832
Chicago/Turabian StyleIslam, Md Ashraful, Md Muntasir Bin Mohi Uddin, Md Golam Rasul, Md Ahsanul Haque Swapon, Minhaz Ahmed, and Mehfuz Hasan. 2022. "In Vitro Screening and Field Performance of EMS-Treated Eggplants for the Selection of Shoot and Fruit Borer-Resistant Plants" Agronomy 12, no. 8: 1832. https://doi.org/10.3390/agronomy12081832