1. Introduction
Hydrangea macrophylla is a popular perennial summer-flowering shrub with an enormous inflorescence and top growth. It is one of the most promising ornamental flower species for use as potted plants, landscape plants, and cut flowers [
1,
2]. It originates from eastern Asia and North America, and is widely planted in the temperate regions and extends southward into the tropics of both hemispheres [
3]. The
H. macrophylla extract is effective against the human malaria parasite, diabetes, and urinary tract infection [
4,
5,
6]. Furthermore, its root bark has been used to treat numerous kidney diseases [
7]. Commercially,
H. macrophylla is propagated either by seeds or stem cuttings. Although seed germination is not difficult, the variability of seedlings is high, and therefore seed propagation does not always lead to the desired characteristics and morphology for the producer. On the other hand, cutting propagation is slow to establish, and plants from cuttings lack good basal branching [
8,
9]. Micropropagation is widely used in ornamental horticulture. However, it is limited for many in vitro perennial trees and vegetative shrubs, and few studies have been conducted with in vitro
H. macrophylla [
10].
Iron (Fe) participates in photosynthesis, mitochondrial respiration, nitrogen assimilation, hormone (ethylene, gibberellic acid, jasmonic acid, etc.) biosynthesis, production and scavenging of reactive oxygen species, osmoprotection, and pathogen defense [
11]. Moreover, Fe influences the formation of coproporphyrin in the tetrapyrrole biosynthetic pathway and is also necessary for chlorophyll synthesis [
12]. The uptake, distribution, and storage of Fe are tightly regulated in plants [
13]. There are two different responses of plant roots induced by Fe deficiency. Dicotyledons and non-gramineous plants utilize the strategy I. Firstly, the activity of a plasma membrane-bound reductase in the root increases when Fe is deficient, which enhances the rate of Fe III reduction and correspondingly splits Fe-III-chelates at the plasma membrane by increasing the acidification of the rhizosphere. Meanwhile, roots produce ferric chelate reductase (FCR) to catalyze Fe
3+ to Fe
2+ [
14]. Then the iron regulated transporter 1 (
IRT1) transports the Fe
2+ into the cells of the root. Simply, it is an acidification-reduction-transport mechanism, and proton pump H
+-ATPase 2 (AHA2), ferric chelate reductase 2 (
FRO2), and IRT1 can assemble an iron-acquisition complex to optimize Fe uptake during this progress [
15]. The expression of these three transporters is regulated by bHLH transcription factors,
FER-like iron deficiency induced transcription factor 1 (
FIT1), which is the homolog gene of
ferritin (
FER) can regulate the accumulation of
FRO2 and
IRT1 [
16]. Gramineae plants utilize a different strategy (strategy II) based on chelation, they can produce the mugineic acid family phytosiderophores (MAs) to complex sparingly soluble inorganic Fe
3+ The iron-related bHLH transcription factor 2 (
IRO2) has a key role to regulate the expression of MAs biosynthetic genes [
17]. Then the TOM1 transports the MAs outflow to the rhizosphere. Finally, the Fe-MAs complexes enter the root by the yellow stripe-like 1 (YSL1) [
18]. Nonetheless, these two strategies are similar; for example, rice can use either strategy I or II according to the concentration of substrate Fe [
17]. The metabolites of the partial strategy I plants have the characteristic of strategy II plants [
19]. Moreover, the transportation of Fe is dependent on the nicotianamine (NA) both in strategy I and strategy II in the symplast [
20]. Although the molecular mechanism of Fe-deficiency responses operating in roots is well documented, the genes controlling these activities in shoots are unknown. The transportation of Fe is not only located in the root but also regulated by the whole plant. For instance, the expressions of
IRT1 and
FEO2 genes and accumulation of MAs protein increased during the day and down-regulated at night, however, accumulation increased during the night when the plant suffered Fe-deficiency. Besides, Fe transport is regulated by the signals of shoot-borne and nutrients such as sugar [
21]. In
Arabidopsis, many of the regulators which have been identified in roots were also found in leaves under Fe deficiency, including the photosynthesis-related genes photosystem I subunit F (
PsAF) and photosystem I subunit N (
PsAN), ferredoxin (
FED), and proteins family light-harvesting complexes-related (LHCB). However, there is little information about the inference of Fe signal transduction in chloroplast and leaf [
22].
It is clear that the effects of substrates on Fe absorption and transport, all related genes of strategy I were decreased at pH 7.5 [
23], and a high concentration of HCO
3− limits the rate of Fe uptake due to the solubility of Fe decreases 1000-fold for each unit increase of pH. [
24]. Therefore, local acidification of calcareous soils is found to be an effective, inexpensive means to alleviate iron chlorosis [
25]. Besides, injecting iron solution into the trunks of trees or the surrounding soils, and mixing iron into the root systems at the transplanting time are corrective procedures for Fe deficiency. However, different plants have different sensitivities or tolerances to iron deficiency. For acidophilus plants, the most obvious effect of Fe deficiency is the youngest leaves turning yellow and stunt plant growth due to the decrease in photosynthetic efficiency and electron transport through PSII [
13]. Interestingly, the activity of PSI in chlorosis leaves was only slightly inhibited under Fe deficiency [
26]. Besides, at the ultrastructural level, although the thylakoid system markedly decreases, other Fe-containing organelles as mitochondria and peroxisomes are immune [
27]. One view is that the Fe deficiency induces a large xanthophyll cycle. This cycle is beneficial to photoprotection and reduces the injury by increasing leaves oxygen concentration [
28,
29]. However, another view is the xanthophyll cycle and the resynthesis of photosynthetic pigments in Fe deficiency plants occur after Fe resupply [
30]. Therefore, the effect of Fe on photosynthesis and even for the physiological activities of the whole plant is still incomplete.
The
H. macrophylla is susceptible and slowly dies in response to Fe deficiency. However, the physiological activity of Fe as affected by deficiency or supplementation is unknown. It is necessary to investigate the responses to different Fe sources and pH levels. In our previous studies, supplemental FeSO
4 or Fe-EDTA have been found to have a growth-promoting effect on sensitive plants
Sorbus commixta [
31]. However, the regulation mode of Fe is not discussed in depth. In this study, the photosynthesis and the content of the pigment of leaves were measured, and the macro- and micronutrients in leaves were analyzed. Lastly, we studied the relative expressions of Fe transfer and photosynthesis genes, to dissect the response of
H. macrophylla Fe deficiency, and preliminary explore the relationship between Fe nutrient and photosynthesis, and also to find the optimal combination for alleviating chlorosis for
H.
macrophylla. 4. Discussion
The concept of pH was derived from the ion product of water and was defined in terms of the hydrogen ion activity; namely, the negative logarithm of the hydrogen ion concentration [H
+]. Researchers have found that the color of
H. macrophylla sepals is very sensitive to the cultivation conditions and changes easily from red through purple, as the pH changes from 4.5 to 6.5 [
41]. The pH influences the chemical solubility and the availability of essential nutrients for plants. Since H
+ is a cation, it will compete with other cations for exchange sites. In alkaline soils, metal availability is often inhibited and causes micronutrient deficiencies in plants [
42,
43]. In general,
H. macrophylla develops perfectly in acidic soils [
44]. Moreover, a soil pH lower than 6.0 is beneficial to the formation of inflorescence and prolonged flowering duration, and increases plant growth [
45,
46]. In order to prevent Fe deficiency and assist in coloration, the suggested substrate pH range is 5.2 to 5.6 for greenhouse cultivation of
H. macrophylla [
47]. For Calcisol soil, only adjusting pH is not a good way to reduce the chlorosis, organic acids such as citric acid (CA), oxalic acid (OA), salicylic acid (SA), or humic complexes (HCs) combined with Fe chelate improved the quality of tomato [
48]. In this study, the Fe accumulation in plantlets was promoted by a lower pH. On the contrary, the FCR activities were greatly enhanced at higher pH levels. Since the concentration of bio-available Fe is low, plants of strategy Ι usually up-regulated the FCR enzymes to take up the Fe in a controlled manner into the roots [
49]. The enzymatic reduction of any Fe (III) complex must interact with the FCR to transfer the site of the electron [
50]. However, the FCR reaction was most optimal at a pH of 5.0–5.5 [
51]. A huge decrease (40 times) in the FCR reaction was observed in sugar beets (
Beta vulgaris L.) at pH levels exceeding 6.2 [
52]. Moreover, plants can reduce Fe uptake by rapidly decreasing FCR activity when Fe is excess. For example, the FCR activity in the rhizosphere of spinach and kale markedly decreased with increased supplemental Fe [
53]. In this study, the Fe content was high in treatments with supplementary Fe-EDTA, as the Fe-EDTA reduced Fe to Fe
2+ through the metal charge transfer reaction and the ligand is oxidatively degraded instead of being precipitated as the insoluble Fe oxides [
54,
55]. And the FCR reduction rates were slightly decreased due to the high stability of Fe-chelates [
56]. Therefore, in treatments with supplementary Fe-EDTA, the FCR activities were lower than in treatments with supplementary FeSO
4. However, for chlorosis plants, Fe concentration is more important compared with FCR activity, where foliar Fe-EDTA sprays were more effective than foliar FeSO
4 sprays for correcting Fe deficiency in calibrachoa (
Calibrachoa hybrida) [
57].
Plants require different amounts of different nutrients. Macronutrients are needed in large amounts by plants, whereas micronutrients are essential to plants but only in small amounts. These nutrients interact with each other, in which deficiency or excess of these nutrients can upset this balance. Fe is indispensable for maintaining the metal homeostasis in plants. The researcher showed that the phytosiderophores also increase Zn, Cu, Mn, and Ni concentrations in wheat [
58]. However, using Cu, Mn, and Zn as fertilizers for a long time is toxic to plants and may cause Fe deficiency. In this study, the content change of Fe also leads to this variation of Mn and Cu contents. Due to the Fe antagonistically interacting with other cationic micronutrients [
59]. Differently, the Zn content does not seem to be significantly affected by supplementary Fe-EDTA, as Fe-EDTA exchanges and re-complexes to Zn-EDTA, resulting plants absorb Zn-chelate [
60]. Similar result was found that both can increase the contents of Zn and Fe in plants by supplemental Zn and Fe fertilizer or only EDTA [
61,
62]. The leaf is the main sink of metal nutrients and chloroplasts allocate 80% of the Fe [
63]. Furthermore, chloroplasts are a major site of the Fe function and ferritin storage for photosynthesis [
49]. Fe influences photosynthesis through material structures and electron transport. Within the chloroplasts, 70~72% of the Fe is located in the thylakoids [
64]. Thus, Fe deficiency will cause thylakoid growth to stagnate, and decrease the contents of chlorophyll and cytochrome
f [
63]. It is more objective to use the chlorophyll concentration than to use visual scores to test the Fe deficiency chlorosis [
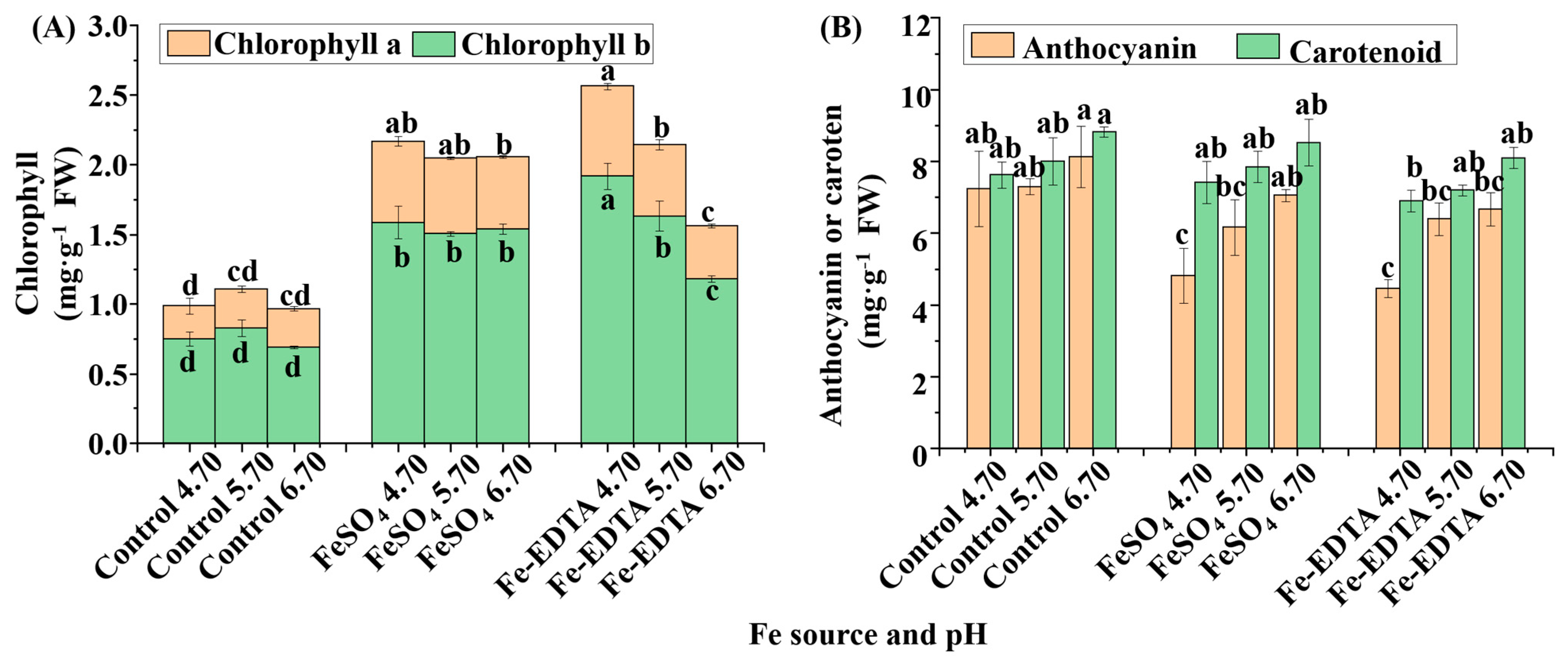
65]. In this study, we observed that the Fe deficiency led to greatly reduced chlorophyll contents, and the chlorophyll content decreased with the decrease in the Fe content. It was illustrated that the utilization of supplementary FeSO
4 and Fe-EDTA could decrease chlorosis, which was in agreement with the findings in groundnut (
Arachis hypogaea L.) and peppermint (
Mentha piperita L.) [
66,
67]. Moreover, Fe deficiency caused reduction of the stomatal pores length, cuticular weight per unit surface, cuticular waxes, and stomatal turgor [
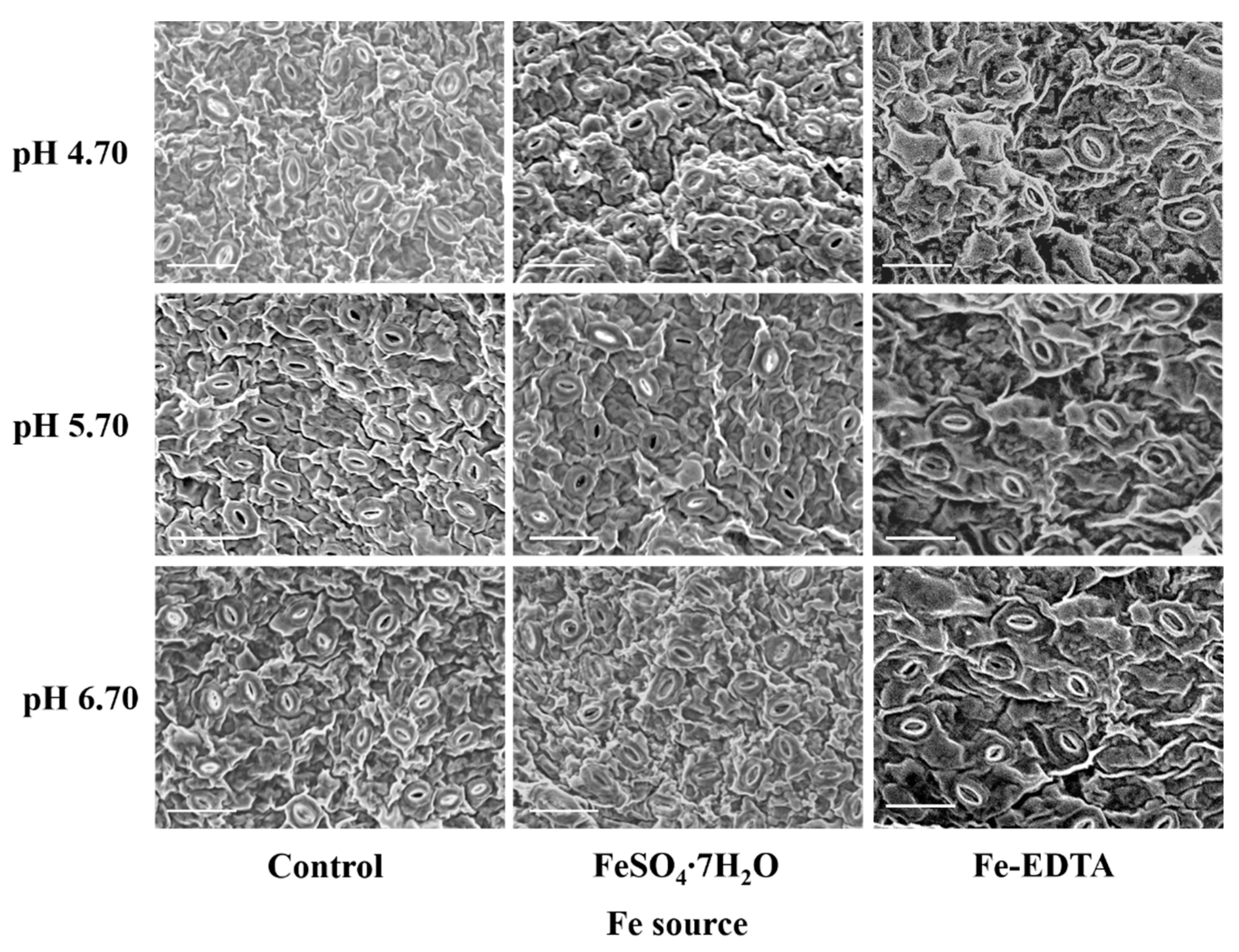
68]. In this study, the treatments with supplementary Fe led to a lower stomatal density and higher stomatal size. In general, the stomatal density was negatively correlated with the stomatal size. Nevertheless, guard cells of stomata should be bigger than their nucleus [
69], and according to the rule of “one-cell-spacing”, the stomata are separated by at least one epidermal cell in order to provide enough space for the other structures [
70]. Besides, stomatal clustering will negatively impact the gas exchange rate [
71]. Therefore, the plantlets with a lower number of larger stomata are more fit to maintain an appropriate photosynthetic rate [
72]. It was worth noting that the shoot weight of pH 4.70 is higher than that of pH 6.70 with supplemental FeSO
4, despite the leaf area and width of the latter were significantly higher than that of the former. The reason is that leaves with higher Fe content had better cuticle and epidermis development, and the biomass of petiol and stem were higher. In the photosynthetic electron chain, Fe acts as the ligature for PsbA and PsbD in PSII, PSII, and cytochrome b6/
f (Cyt b6/
f) in hemeprotein, and Cyt b6/
f and ferredoxin in Fe–Sulfur [Fe–S] [
73]. Therefore, chlorosis impairs the PSII while the PSΙ mostly unaffected [
26]. As shown by the results, Fe deficiency decreased the Fv/F0 and Fv/Fm, findings which are consistent with those in pear and sugar beet [
74,
75]. Moreover, Fe deficiency decreased the activity of Rubisco, which regulates the Rubisco carboxylation efficiency [
76], it has been reported that there are a high correlations among apparent Rubisco carboxylation efficiencies, leaf absorptances, and PSII efficiencies [
77]. To sum up, the evidence is that Fe deficiency reduces photosynthesis by decreased chlorophyll, gas exchange, and activities of PS. However, more research is still needed.
The relationships between the leaf color, and the contents of chlorophyll, anthocyanin, and carotenoid are noteworthy. Supplementary Fe increased the anthocyanin contents in leaves. Similarly, it was reported that Fe regulates the anthocyanin biosynthetic pathway and Fe supply increased the total anthocyanins content in
Vitis vinifera [
78]. However, it has not yet been elucidated if Fe has a direct or indirect effect on anthocyanins and their biosynthesis pathway. Although the accumulation of carotenoids and flavonoids mainly affects the flower color, they also influence the leaf color. Anthocyanin has a negative effect on photosynthesis, which could absorb more green light and accelerate the degradation of thylakoids, which causes a decrease in the photosynthetic rate [
79]. Another suggestion is that anthocyanin acts as an antioxidant scavenging free radical and reduces the electron chain in photosynthesis [
80]. The process is similar to the xanthophyll cycle. It has been reported that Fe deficient increased zeaxanthin to form light protection in pear (
Pyrus communis) [
74]. However, there is no evidence that anthocyanin was prominently photo-inhibitory or antioxidant in this study. It is can be determined that changes in the anthocyanins often indicate the variety of the antioxidant system. The SOD defends against oxidation and produces H
2O
2, then the H
2O
2 is scavenged by APX, CAT, and POD. It is reported that a higher SOD amount is found in purple leaves, which contain more anthocyanins, and H
2O
2 can induce the signal for anthocyanin biosynthesis [
79,
81]. In addition, APX, CAT, and POD act as the hemeproteins and their activities are affected both by the Fe content and ROS [
82]. It has been reported that POD is more sensitive to H
2O
2 than CAT [
83]; therefore, in this study, the activity of POD increased under pH 5.70, and similar results were observed with the activity of APX, it is speculated that the APX has a higher sensitivity to H
2O
2. In brief, as shown in
Figure 10, Fe deficiency can directly reduce chlorophyll synthesis and affect plant growth. Furthermore, Fe deficiency increases the ROS content, which induces the antioxidant system, while insufficient antioxidant hemeproteins exacerbate the damage to plantlets.
The control of the Fe-homeostasis is handled at the genetic level. First, the reduction of the ferric chelates by ferric reductase is an obligatory step in Strategy Ι plants. Overexpression of
FRO2 in
Arabidopsis thaliana was observed to correspond with a significant increase in the tissue content of Fe [
84]. Meanwhile, the
AtFRO2 is controlled by a transcriptional factor
FER, reported to be the spontaneous mutant
T3238fer, which is unable to activate Fe-deficiency responses under low Fe conditions, which includes but is not limited to the expression of
FRO and the activity of ferric chelate reductase [
85]. In this study, Fe deficiency upregulated the expression of
Hm-AtFRO2 and
Hm-AtFER1, whereas the
Hm-AtFER1 in the control at pH 5.70 was significantly higher than that in the control group at the other two pH levels, which is speculated to be related with the underlying FER regulatory pathway in strategy Ι. The detailed mechanisms involved remain to be revealed. Second, once inside the roots, Fe is transported over long distances via the veins to the tissues by transpiration. Previous researchers have found that aerial part of the plant has a key role in the distribution and regulation of Fe, in both strategy I and strategy II plants, intracellular delivery of Fe and trafficking within the plant is dependent on NA, which chelates and transports micronutrient metal ions. The lack of NA would decrease the mobility and availability of Fe, as well as form precipitation in the chloroplast and phloem sap [
22]. The Fe-NA complexes are transferred from the xylem to the neighboring cells through the YSL proteins in
Arabidopsis [
86]. These are regulated by
NAS and
YSL. Analysis of the Fe-sufficient and Fe-deficient transcriptome in
Arabidopsis revealed that NAS4 was strongly Fe-regulated in leaves [
22]. Differently, in this study,
Hm-AtNAS3 and
Hm-AtNAS5 were respectively upregulated by Fe deficiency and supplementary Fe. It is speculated that these genes are also regulated by Cu and Zn [
87], as the YSL protein is able to transport metal-NA complexes and NA has a high affinity for the two metals. The YSL transporters belong to the OPT family. Under Fe deficiency, the
YSL5 was upregulated nearly twice in order to improve the Fe nutrient condition in apple [
88]. Similar results were found with
YSL7 in peanut. The
OPT3, which also belongs to the OPT family, is preferentially expressed in the phloem cells during Fe deficiency [
89]. The
OPT3 functions in the phloem-mediated Fe redistribution, suggesting that
OPT3 transfers the Fe out of the leaves [
90]. Therefore, the increased expression of
Hm-AtOPT3 in the control group exacerbated the Fe deficiency in the leaves. It is speculated that the shoots feedback the Fe deficiency signals to the roots through phloem. Similar reports found after foliar spray Fe, leaves of
A. thaliana transmit signals to roots via the Fe-peptide transport regulated. Besides, Fe content of the phloem in Fe deficiency leaves was lower than in Fe supplemental leaves in castor oil (
Ricinus communis L.) and dwarf bean (
Phaseolus vulgris L.) [
91,
92], and the unallocated Fe is stored by the leaves [
93]. The chlorosis did the most damage to the PSII, and it has been documented that PSII is more sensitive to Fe deficiency [
26,
94,
95]. Another view is PSI is a prime target for Fe deficiency due to its high Fe demand [
96]. In this study, the results showed that there was the huge upregulated expression of
Hm-AtPsAN and
Hm-AtPsAF with supplementary Fe. PsAN and PsAF proteins are connected with PSI and plastocyanin (Pc), where the function of the latter is the transfer of electrons from Cyt b6/
f to the PSI complex in the thylakoid lumen [
97,
98]. Interestingly, reported found in the thylakoid membranes of Fe deficiency leaves, core proteins of PSI (PsaA) declined drastically, while the core proteins of PSII (PsbA) remained stable relatively [
99,
100]. Furthermore, the IsiA protein, which is homologous to the inner antenna protein of PSII (PsbC), and expressed specifically under Fe deficiency. It is involved in the state transitions of light sharing between two photosystems, which needs further study.















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}