
Cloning and Expression of Pigeon-Derived Escherichia coli Type 1 Pilus Clusters and Analysis of Amino Acid Sequence Characteristics of Functional Proteins


Abstract
:1. Introduction
2. Materials and Methods
2.1. Strains
2.2. Primer Design
2.3. DNA Extraction
2.4. PCR Amplification, Cloning, and DNA Purification
2.5. Construction of the Recombinant Bacteria
2.6. Transmission Electron Microscopy (TEM)
2.7. Hemagglutination Assay and D-Mannose Sensitive Hemagglutination Assay (MSHA)
2.8. Complete Gene Sequence Determination
2.9. Amino Acid Sequence Analysis
3. Results
3.1. Construction of Recombinant Bacteria and Expression of Pili
3.1.1. Results of PCR Amplification of fimBH
3.1.2. Results of PCR Amplification of fimBG
3.1.3. Hemagglutination and MSHA
3.1.4. Determination of the Complete Gene Sequence of Type 1 Pili
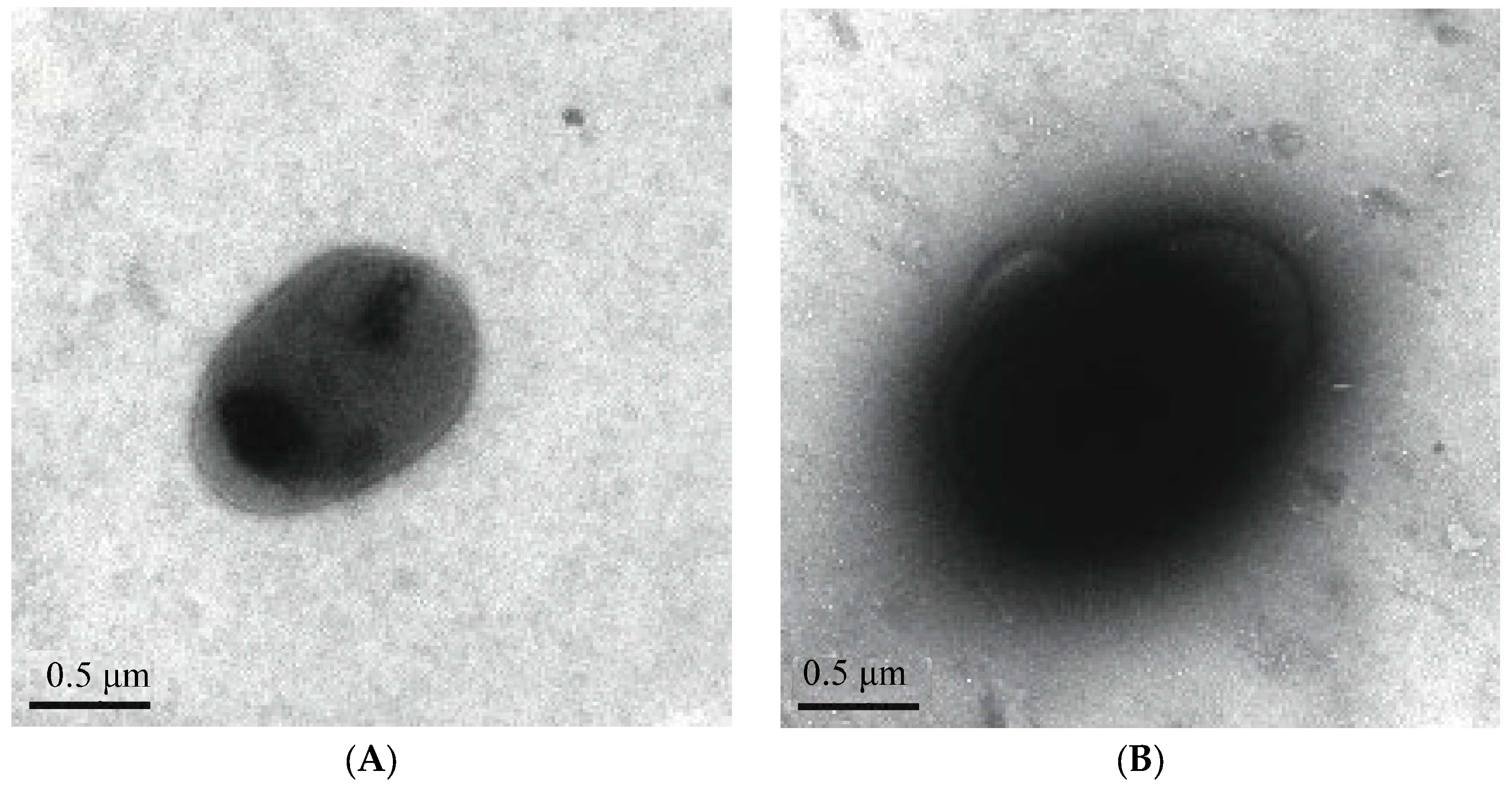
3.1.5. TEM Observation of Type 1 Pili
3.2. Comparison of Amino Acids of Pilus Rod FimA Proteins from Different Sources
3.3. Comparative Analysis of Amino Acid Sequences of Regulatory Proteins fimB and fimE
3.4. Comparative Amino Acid Sequence Analysis of FimC Chaperone Proteins
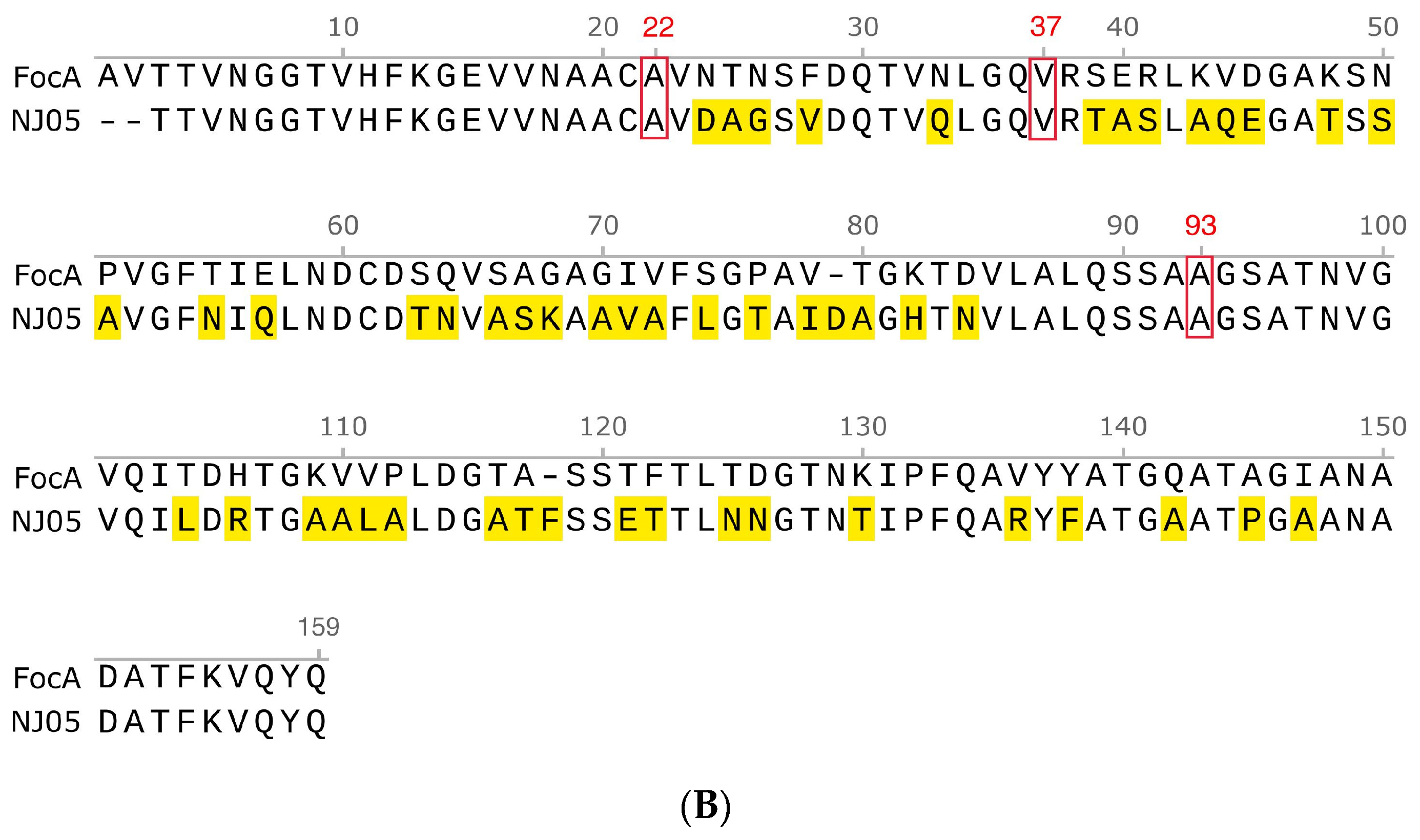
3.5. Amino Acid Sequence Analysis of FimD Protein
3.6. Amino Acid Sequence Analysis of FimH Protein
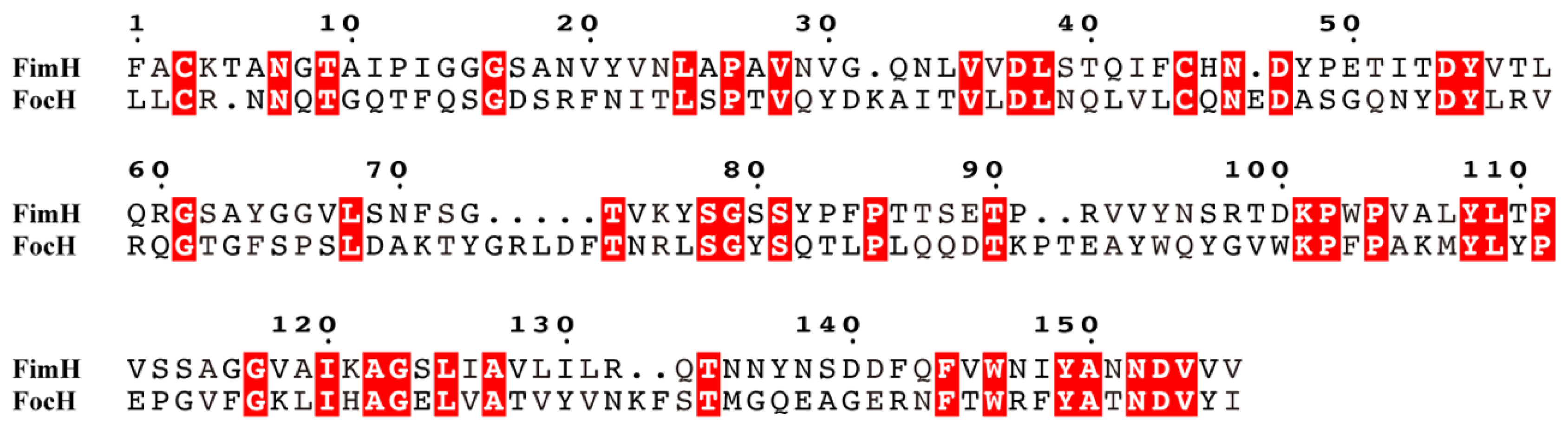
3.7. Comparison of Amino Acid Sequences of Receptor Binding Domains of FimH and FocH Proteins
3.8. Comparison of Amino Acid Sequences of the Pilus Binding Domains of FimH and FocH Proteins
3.9. Comparison of Amino Acid Sequences of FimH, FimA, FimF, and FimG Proteins
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Dozois, C.M.; Chanteloup, N.; Dho-Moulin, M.; Brée, A.; Desautels, C.; Fairbrother, J.M. Bacterial colonization and in vivo expression of F1 (type 1) fimbrial antigens in chickens experimentally infected with pathogenic Escherichia coli. Avian Dis. 1994, 38, 231–239. [Google Scholar] [CrossRef] [PubMed]
- Gyimah, J.E.; Panigrahy, B. Adhesin-receptor interactions mediating the attachment of pathogenic Escherichia coli to chicken tracheal epithelium. Avian Dis. 1988, 32, 74–78. [Google Scholar] [CrossRef] [PubMed]
- Dozois, C.M.; Pourbakhsh, S.A.; Fairbrother, J.M. Expression of P and type 1 (F1) fimbriae in pathogenic Escherichia coli from poultry. Vet. Microbiol. 1995, 45, 297–309. [Google Scholar] [CrossRef] [PubMed]
- Spaulding, C.N.; Klein, R.D.; Ruer, S.; Kau, A.L.; Schreiber, H.L.; Cusumano, Z.T.; Dodson, K.W.; Pinkner, J.S.; Fremont, D.H.; Janetka, J.W.; et al. Selective depletion of uropathogenic E. coli from the gut by a FimH antagonist. Nature 2017, 546, 528–532. [Google Scholar] [CrossRef] [PubMed]
- Tomasek, K.; Leithner, A.; Glatzova, I.; Lukesch, M.S.; Guet, C.C.; Sixt, M.; Zanoni, I. Type 1 piliated uropathogenic Escherichia coli hijack the host immune response by binding to CD14. eLife 2022, 11, e78995. [Google Scholar] [CrossRef]
- Guiton, P.S.; Cusumano, C.K.; Kline, K.A.; Dodson, K.W.; Han, Z.; Janetka, J.W.; Henderson, J.P.; Caparon, M.G.; Hultgren, S.J. Combinatorial small-molecule therapy prevents uropathogenic Escherichia coli catheter-associated urinary tract infections in mice. Antimicrob. Agents Chemother. 2012, 56, 4738–4745. [Google Scholar] [CrossRef]
- Svensson, A.; Larsson, A.; Emtenas, H.; Hedenstrom, M.; Fex, T.; Hultgren, S.J.; Pinkner, J.S.; Almqvist, F.; Kihlberg, J. Design and evaluation of pilicides: Potential novel antibacterial agents directed against uropathogenic Escherichia coli. Chembiochem 2001, 2, 915–918. [Google Scholar] [CrossRef]
- Orndorff, P.E.; Falkow, S. Nucleotide sequence of pilA, the gene encoding the structural component of type 1 pili in Escherichia coli. J. Bacteriol. 1985, 162, 454–457. [Google Scholar] [CrossRef]
- Schwan, W.R.; Beck, M.T.; Hung, C.S.; Hultgren, S.J. Differential Regulation of Escherichia coli fim Genes following Binding to Mannose Receptors. J. Pathog. 2018, 2018, 2897581. [Google Scholar] [CrossRef]
- Holden, N.; Blomfield, I.C.; Uhlin, B.-E.; Totsika, M.; Kulasekara, D.H.; Gally, D.L. Comparative analysis of FimB and FimE recombinase activity. Microbiology 2007, 153, 4138–4149. [Google Scholar] [CrossRef]
- Klemm, P. FimC, a chaperone-like periplasmic protein of Escherichia coli involved in biogenesis of type 1 fimbriae. Res. Microbiol. 1992, 143, 831–838. [Google Scholar] [CrossRef] [PubMed]
- Klemm, P.; Christiansen, G. The fimD gene required for cell surface localization of Escherichia coli type 1 fimbriae. Mol. Gen. Genet. 1990, 220, 334–338. [Google Scholar] [CrossRef] [PubMed]
- Klemm, P.; Christiansen, G. Three fim genes required for the regulation of length and mediation of adhesion of Escherichia coli type 1 fimbriae. Mol. Gen. Genet. 1987, 208, 439–445. [Google Scholar] [CrossRef] [PubMed]
- Schembri, M.A.; Kjaergaard, K.; Sokurenko, E.V.; Klemm, P. Molecular characterization of the Escherichia coli FimH adhesin. J. Infect. Dis. 2001, 183 (Suppl. S1), S28–S31. [Google Scholar] [CrossRef] [PubMed]
- Rabbani, S.; Fiege, B.; Eris, D.; Silbermann, M.; Jakob, R.P.; Navarra, G.; Maier, T.; Ernst, B. Conformational switch of the bacterial adhesin FimH in the absence of the regulatory domain: Engineering a minimalistic allosteric system. J. Biol. Chem. 2018, 293, 1835–1849. [Google Scholar] [CrossRef]
- Tchesnokova, V.; Aprikian, P.; Kisiela, D.; Gowey, S.; Korotkova, N.; Thomas, W.; Sokurenko, E. Type 1 fimbrial adhesin FimH elicits an immune response that enhances cell adhesion of Escherichia coli. Infect. Immun. 2011, 79, 3895–3904. [Google Scholar] [CrossRef]
- Eldridge, G.R.; Hughey, H.; Rosenberger, L.; Martin, S.M.; Shapiro, A.M.; D’Antonio, E.; Krejci, K.G.; Shore, N.; Peterson, J.; Lukes, A.S.; et al. Safety and immunogenicity of an adjuvanted Escherichia coli adhesin vaccine in healthy women with and without histories of recurrent urinary tract infections: Results from a first-in-human phase 1 study. Hum. Vaccin Immunother. 2021, 17, 1262–1270. [Google Scholar] [CrossRef]
- Mian, M.F.; Lauzon, N.M.; Andrews, D.W.; Lichty, B.D.; Ashkar, A.A. FimH can directly activate human and murine natural killer cells via TLR4. Mol. Ther. 2010, 18, 1379–1388. [Google Scholar] [CrossRef]
- Liu, R.; Wu, C.; Li, L.; Chi, F.; Zhang, T.; Xu, Y.; Ji, L.; Chen, Z.; Hu, H.; Zhang, X.; et al. CD48 and α7 Nicotinic Acetylcholine Receptor Synergistically Regulate FimH-Mediated Escherichia coli K1 Penetration and Neutrophil Transmigration Across Human Brain Microvascular Endothelial Cells. J. Infect. Dis. 2019, 219, 470–479. [Google Scholar] [CrossRef]
- Zhang, W.; Xu, L.; Park, H.-B.; Hwang, J.; Kwak, M.; Lee, P.C.W.; Liang, G.; Zhang, X.; Xu, J.; Jin, J.-O. Escherichia coli adhesion portion FimH functions as an adjuvant for cancer immunotherapy. Nat. Commun. 2020, 11, 1187. [Google Scholar] [CrossRef]
- Spaulding, C.N.; Schreiber, H.L.; Zheng, W.; Dodson, K.W.; Hazen, J.E.; Conover, M.S.; Wang, F.; Svenmarker, P.; Luna-Rico, A.; Francetic, O.; et al. Functional role of the type 1 pilus rod structure in mediating host-pathogen interactions. eLife 2018, 7, e31662. [Google Scholar] [CrossRef] [PubMed]
- van Die, I.; van Geffen, B.; Hoekstra, W.; Bergmans, H. Type 1C fimbriae of a uropathogenic Escherichia coli strain: Cloning and characterization of the genes involved in the expression of the 1C antigen and nucleotide sequence of the subunit gene. Gene 1985, 34, 187–196. [Google Scholar] [CrossRef]
- Sokurenko, E.V.; Schembri, M.A.; Trintchina, E.; Kjaergaard, K.; Hasty, D.L.; Klemm, P. Valency conversion in the type 1 fimbrial adhesin of Escherichia coli. Mol. Microbiol. 2001, 41, 675–686. [Google Scholar] [CrossRef] [PubMed]
- Lillington, J.; Geibel, S.; Waksman, G. Biogenesis and adhesion of type 1 and P pili. Biochim. Biophys. Acta 2014, 1840, 2783–2793. [Google Scholar] [CrossRef]
- Pourbakhsh, S.A.; Dho-Moulin, M.; Brée, A.; Desautels, C.; Martineau-Doize, B.; Fairbrother, J.M. Localization of the in vivo expression of P and F1 fimbriae in chickens experimentally inoculated with pathogenic Escherichia coli. Microb. Pathog. 1997, 22, 331–341. [Google Scholar] [CrossRef] [PubMed]
- Kariyawasam, S.; Nolan, L.K. papA gene of avian pathogenic Escherichia coli. Avian Dis. 2011, 55, 532–538. [Google Scholar] [CrossRef] [PubMed]
- Kurabayashi, K.; Agata, T.; Asano, H.; Tomita, H.; Hirakawa, H. Fur Represses Adhesion to, Invasion of, and Intracellular Bacterial Community Formation within Bladder Epithelial Cells and Motility in Uropathogenic Escherichia coli. Infect. Immun. 2016, 84, 3220–3231. [Google Scholar] [CrossRef]
- Uchiya, K.-I.; Kamimura, Y.; Jusakon, A.; Nikai, T. Salmonella Fimbrial Protein FimH Is Involved in Expression of Proinflammatory Cytokines in a Toll-Like Receptor 4-Dependent Manner. Infect. Immun. 2019, 87, e00881-18. [Google Scholar] [CrossRef]
- Walczak, M.J.; Puorger, C.; Glockshuber, R.; Wider, G. Intramolecular donor strand complementation in the E. coli type 1 pilus subunit FimA explains the existence of FimA monomers as off-pathway products of pilus assembly that inhibit host cell apoptosis. J. Mol. Biol. 2014, 426, 542–549. [Google Scholar] [CrossRef]
- Chen, J.; Dai, W.; Cui, S.; Lei, W.; Dai, D. Screening of antigenic epitopes related to the adhesion of the avian Escherichia coli Type 1 Fimbrial Agglutinin Domain. BMC Vet. Res. 2023, 19, 187. [Google Scholar] [CrossRef]
- Wellens, A.; Lahmann, M.; Touaibia, M.; Vaucher, J.; Oscarson, S.; Roy, R.; Remaut, H.; Bouckaert, J. The tyrosine gate as a potential entropic lever in the receptor-binding site of the bacterial adhesin FimH. Biochemistry 2012, 51, 4790–4799. [Google Scholar] [CrossRef] [PubMed]














{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name | Forward (5′-3′) | Size | |
|---|---|---|---|
| Primer 1 | fimBH-F | GCTATGACCATGATTACGCCAAGCTTCCAGCACAGCTAGTGCGCGTCTG | 8924 bp |
| fimBH-R | GTAAAACGACGGCCAGTGAATTCTGGCCTACAAAGGGCTAACGTG | ||
| Primer 2 | fimBG-F | GCTATGACCATGATTACGCCAAGCTTCCAGCACAGCTAGTGCGCGTCTG | 7927 bp |
| fimBG-R | GTAAAACGACGGCCAGTGAATTCCGGGTTCAGCTGTAGGTATAGGTG | ||
| Primer M13 | M13 rev | CAGGAAACAGCTATGAC | 8934 bp |
| M13 fwd | TGTAAAACGACGGCCAGT | ||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chen, J.; Dai, W.; Wang, H.; Lei, W.; Fang, G.; Dai, D. Cloning and Expression of Pigeon-Derived Escherichia coli Type 1 Pilus Clusters and Analysis of Amino Acid Sequence Characteristics of Functional Proteins. Genes 2024, 15, 1253. https://doi.org/10.3390/genes15101253
Chen J, Dai W, Wang H, Lei W, Fang G, Dai D. Cloning and Expression of Pigeon-Derived Escherichia coli Type 1 Pilus Clusters and Analysis of Amino Acid Sequence Characteristics of Functional Proteins. Genes. 2024; 15(10):1253. https://doi.org/10.3390/genes15101253
Chicago/Turabian StyleChen, Junhong, Wei Dai, Hang Wang, Weiqiang Lei, Guangyuan Fang, and Dingzhen Dai. 2024. "Cloning and Expression of Pigeon-Derived Escherichia coli Type 1 Pilus Clusters and Analysis of Amino Acid Sequence Characteristics of Functional Proteins" Genes 15, no. 10: 1253. https://doi.org/10.3390/genes15101253