
Phosphorus-Use-Efficiency Gene Identification in Fabaceae and RSL2 Expansion in Lupinus albus Is Associated with Low-Phosphorus Adaptation

Abstract
:1. Introduction
2. Materials and Methods
2.1. Downloading the Published Genomes of Fabaceae Crops
2.2. Analysis of Evolutionary Relationships of Fabaceae Crops
2.3. Identification of Phosphorus-Utilization Genes in Fabaceae Crops
2.4. Definition of the Amplification Modes of Homologous Genes
2.5. The Structure and Motif Prediction of Homologous Genes
2.6. Analysis of Cis-Acting Element of RSL2 Genes
2.7. Response of White Lupin to P Deficiency
3. Results
3.1. Identification of Phosphorus-Use-Efficiency Gene Families in 35 Fabaceae Genomes
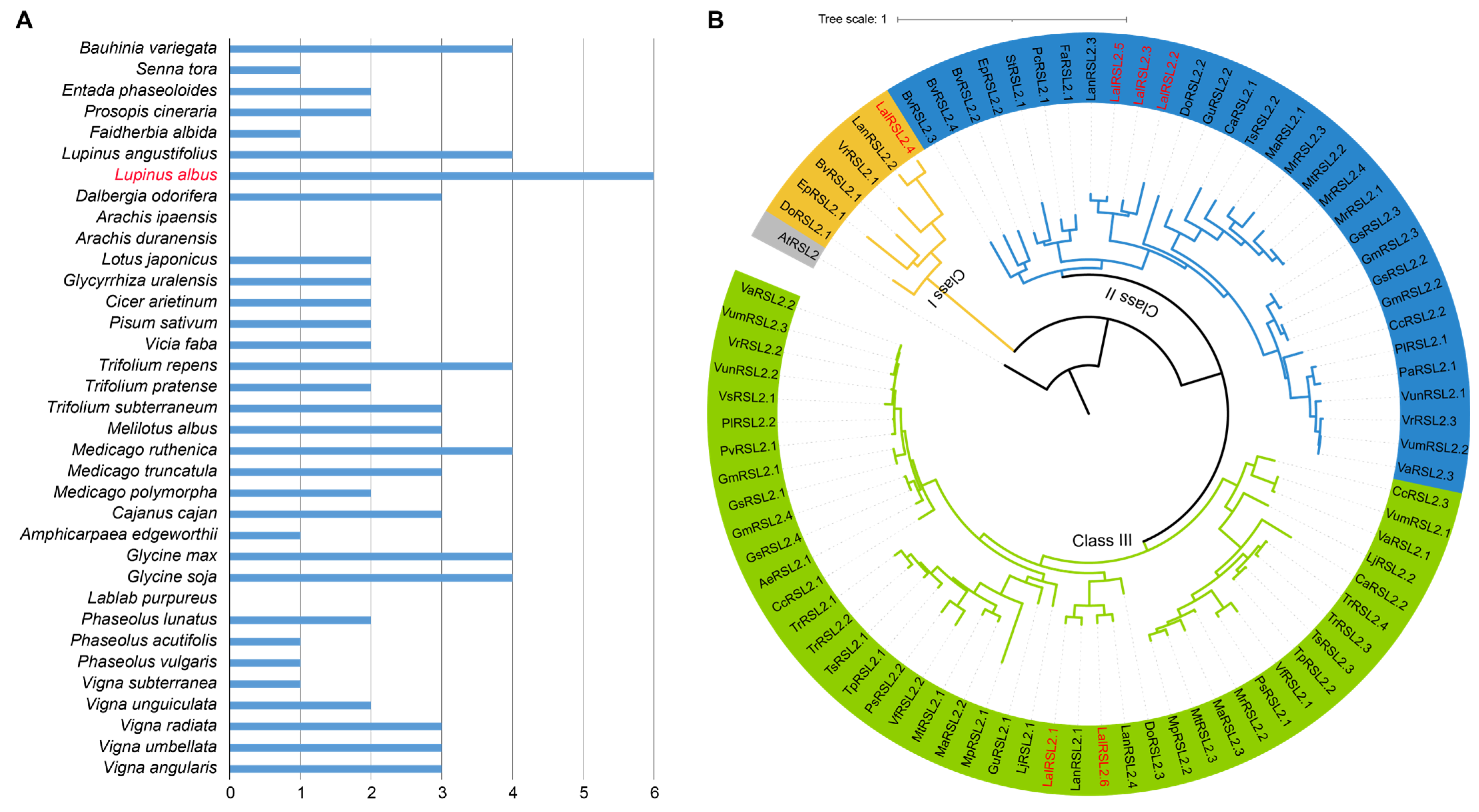
3.2. Over-Retention of RSL2 Genes in L. albus
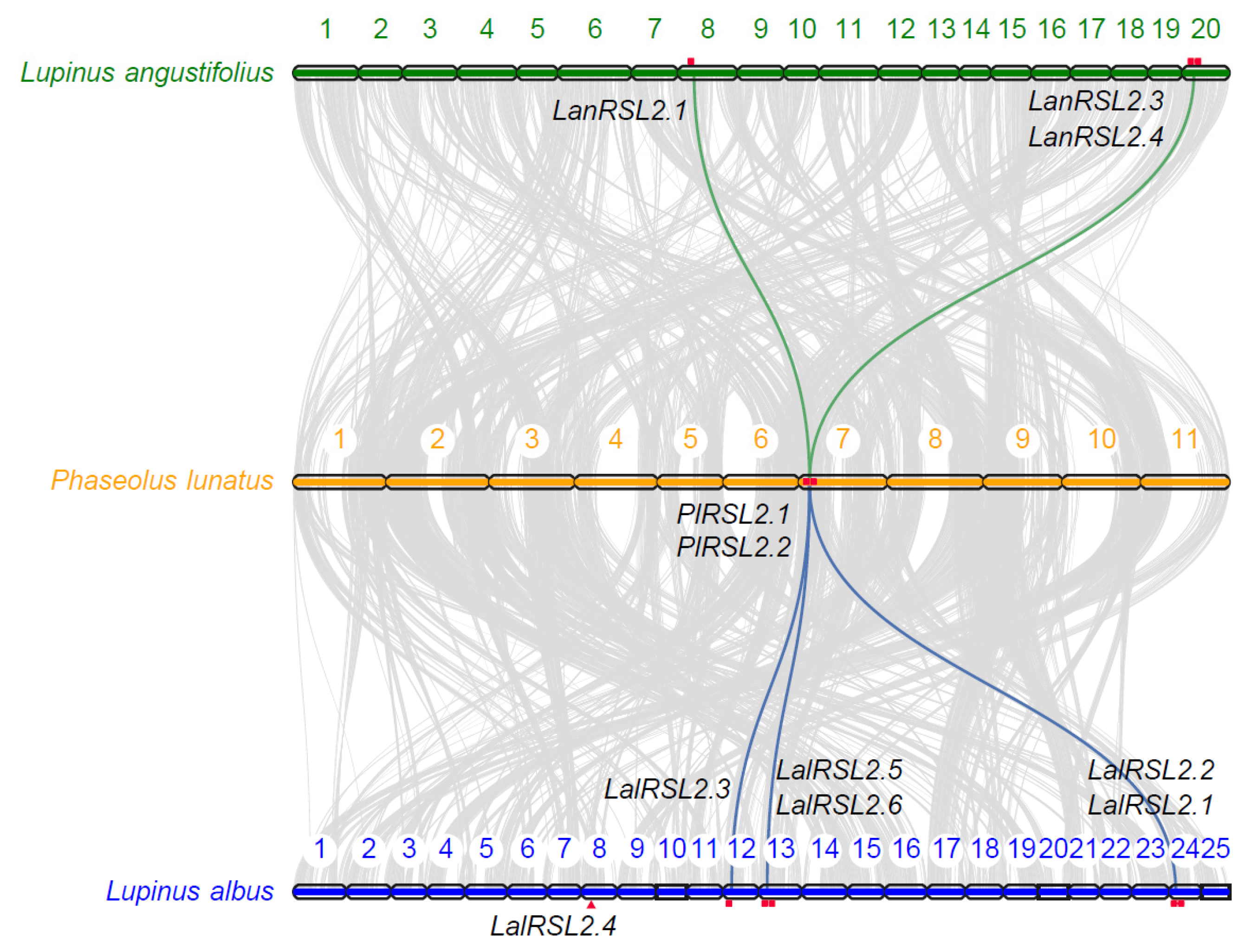
3.3. RSL2 Genes Expanded through WGT in L. albus
3.4. LalRSL2s Exhibit a Diverse Gene Structure and Motif Composition
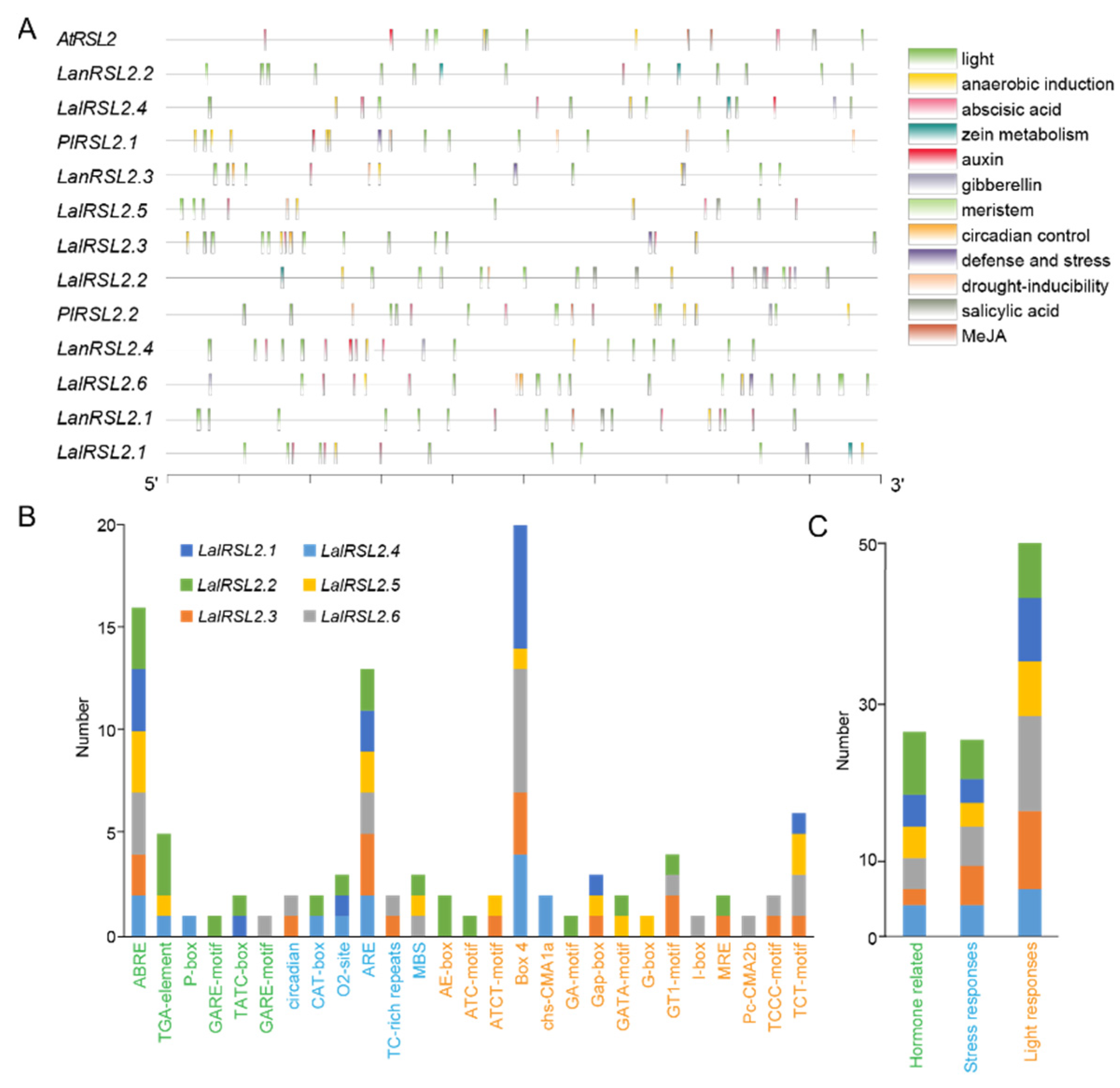
3.5. LalRSL2s Contain Various Cis-Acting Elements in Promoter Regions
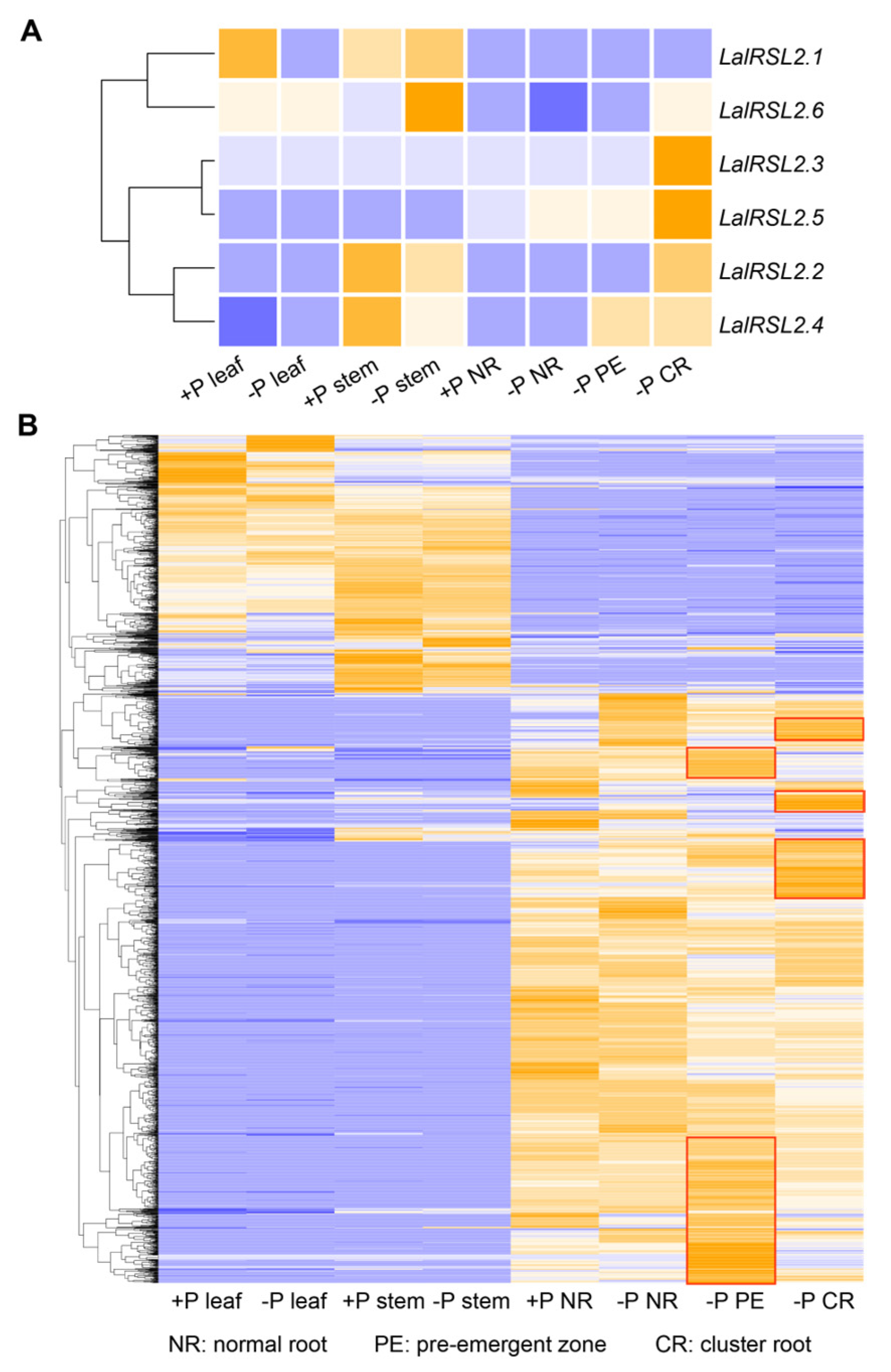
3.6. LalRSL2s Exhibit Tissue-Specific Expression under Low-Phosphorus Stresses
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Schrire, B.P.; Lewis, G.P.; Lavin, M.F.; Mackinder, B.; Lock, M. Legumes of the World; Royal Botanic Gardens: Kew, UK, 2005. [Google Scholar]
- Cannon, S.B.; McKain, M.R.; Harkess, A.; Nelson, M.N.; Dash, S.; Deyholos, M.K.; Peng, Y.H.; Joyce, B.; Stewart, C.N.; Rolf, M.; et al. Multiple Polyploidy Events in the Early Radiation of Nodulating and Nonnodulating Legumes. Mol. Biol. Evol. 2015, 32, 193–210. [Google Scholar] [CrossRef] [PubMed]
- Cui, L.Y.; Wall, P.K.; Leebens-Mack, J.H.; Lindsay, B.G.; Soltis, D.E.; Doyle, J.J.; Soltis, P.S.; Carlson, J.E.; Arumuganathan, K.; Barakat, A.; et al. Widespread genome duplications throughout the history of flowering plants. Genome Res. 2006, 16, 738–749. [Google Scholar] [CrossRef] [PubMed]
- Van De Peer, Y.; Mizrachi, E.; Marchal, K. The evolutionary significance of polyploidy. Nat. Rev. Genet. 2017, 18, 411–424. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.Y.; Zhang, R.; Jiang, K.W.; Qi, J.; Hu, Y.; Guo, J.; Zhu, R.B.; Zhang, T.K.; Egan, A.N.; Yi, T.S.; et al. Nuclear phylotranscriptomics and phylogenomics support numerous polyploidization events and hypotheses for the evolution of rhizobial nitrogen-fixing symbiosis in Fabaceae. Mol. Plant 2021, 14, 748–773. [Google Scholar] [CrossRef] [PubMed]
- Li, J.L.; Shen, J.T.; Wang, R.; Chen, Y.M.; Zhang, T.K.; Wang, H.F.; Guo, C.C.; Qi, J. The nearly complete assembly of the Cercis chinensis genome and Fabaceae phylogenomic studies provide insights into new gene evolution. Plant Commun. 2023, 4, 100422. [Google Scholar] [CrossRef] [PubMed]
- Bertioli, D.J.; Moretzsohn, M.C.; Madsen, L.H.; Sandal, N.; Leal-Bertioli, S.C.M.; Guimaraes, P.M.; Hougaard, B.K.; Fredslund, J.; Schauser, L.; Nielsen, A.M.; et al. An analysis of synteny of Arachis with Lotus and Medicago sheds new light on the structure, stability and evolution of legume genomes. BMC Genom. 2009, 10, 45. [Google Scholar] [CrossRef] [PubMed]
- Blanc, G.; Wolfe, K.H. Functional divergence of duplicated genes formed by polyploidy during Arabidopsis evolution. Plant Cell 2004, 16, 1679–1691. [Google Scholar] [CrossRef]
- Cannon, S.B.; Sterck, L.; Rombauts, S.; Sato, S.; Cheung, F.; Gouzy, J.; Wang, X.; Mudge, J.; Vasdewani, J.; Scheix, T.; et al. Legume genome evolution viewed through the Medicago truncatula and Lotus japonicus genomes. Proc. Natl. Acad. Sci. USA 2006, 103, 18026. [Google Scholar] [CrossRef] [PubMed]
- Li, Q.G.; Zhang, L.; Li, C.; Dunwell, J.M.; Zhang, Y.M. Comparative Genomics Suggests That an Ancestral Polyploidy Event Leads to Enhanced Root Nodule Symbiosis in the Papilionoideae. Mol. Biol. Evol. 2013, 30, 2602–2611. [Google Scholar] [CrossRef]
- Pfeil, B.E.; Schlueter, J.A.; Shoemaker, R.C.; Doyle, J.J. Placing paleopolyploidy in relation to taxon divergence: A phylogenetic analysis in legumes using 39 gene families. Syst. Biol. 2005, 54, 441–454. [Google Scholar] [CrossRef]
- Salman-Minkov, A.; Sabath, N.; Mayrose, I. Whole-genome duplication as a key factor in crop domestication. Nat. Plants 2016, 2, 16115. [Google Scholar] [CrossRef] [PubMed]
- Schlueter, J.A.; Dixon, P.; Granger, C.; Grant, D.; Clark, L.; Doyle, J.J.; Shoemaker, R.C. Mining EST databases to resolve evolutionary events in major crop species. Genome 2004, 47, 868–876. [Google Scholar] [CrossRef] [PubMed]
- Young, N.D.; Debellé, F.; Oldroyd, G.E.D.; Geurts, R.; Cannon, S.B.; Udvardi, M.K.; Benedito, V.A.; Mayer, K.F.X.; Gouzy, J.; Schoof, H.; et al. The Medicago genome provides insight into the evolution of rhizobial symbioses. Nature 2011, 480, 520–524. [Google Scholar] [CrossRef]
- Cordell, D.; Drangert, J.O.; White, S. The story of phosphorus: Global food security and food for thought. Glob. Environ. Change 2009, 19, 292–305. [Google Scholar] [CrossRef]
- Zhang, C.Y.; Simpson, R.J.; Kim, C.M.; Warthmann, N.; Delhaize, E.; Dolan, L.; Byrne, M.E.; Wu, Y.; Ryan, P.R. Do longer root hairs improve phosphorus uptake? Testing the hypothesis with transgenic Brachypodium distachyon lines over-expressing endogenous RSL genes. New Phytol. 2018, 217, 1654–1666. [Google Scholar] [CrossRef]
- Xu, W.F.; Shi, W.M.; Jia, L.G.; Liang, J.S.; Zhang, J.H. TFT6 and TFT7, two different members of tomato 14-3-3 gene family, play distinct roles in plant adaption to low phosphorus stress. Plant Cell Environ. 2012, 35, 1393–1406. [Google Scholar] [CrossRef] [PubMed]
- Bhosale, R.; Giri, J.; Pandey, B.K.; Giehl, R.F.H.; Hartmann, A.; Trainil, R.; Truskina, J.; Leftley, N.; Hanlon, M.; Swarup, K.; et al. A mechanistic framework for auxin dependent Arabidopsis root hair elongation to low external phosphate. Nat. Commun. 2018, 9, 1409. [Google Scholar] [CrossRef] [PubMed]
- Vance, C.P.; Uhde-Stone, C.; Allan, D.L. Phosphorus acquisition and use: Critical adaptations by plants for securing a nonrenewable resource. New Phytol. 2003, 157, 423–447. [Google Scholar] [CrossRef] [PubMed]
- Williamson, L.C.; Ribrioux, S.P.C.P.; Fitter, A.H.; Leyser, H.M.O. Phosphate availability regulates root system architecture in Arabidopsis. Plant Physiol. 2001, 126, 875–882. [Google Scholar] [CrossRef]
- Nacry, P.; Canivenc, G.; Muller, B.; Azmi, A.; Van Onckelen, H.; Rossignol, M.; Doumas, P. A role for auxin redistribution in the responses of the root system architecture to phosphate starvation in Arabidopsis. Plant Physiol. 2005, 138, 2061–2074. [Google Scholar] [CrossRef]
- Xu, W.F.; Zhang, Q.; Yuan, W.; Xu, F.Y.; Aslam, M.M.; Miao, R.; Li, Y.; Wang, Q.W.; Li, X.; Zhang, X.; et al. The genome evolution and low-phosphorus adaptation in white lupin. Nat. Commun. 2020, 11, 1069. [Google Scholar] [CrossRef] [PubMed]
- Cheng, L.Y.; Tang, X.Y.; Vance, C.P.; White, P.J.; Zhang, F.S.; Shen, J.B. Interactions between light intensity and phosphorus nutrition affect the phosphate-mining capacity of white lupin (Lupinus albus L.). J. Exp. Bot. 2014, 65, 2995–3003. [Google Scholar] [CrossRef] [PubMed]
- Yi, K.; Menand, B.; Bell, E.; Dolan, L. A basic helix-loop-helix transcription factor controls cell growth and size in root hairs. Nat. Genet. 2010, 42, 264–267. [Google Scholar] [CrossRef] [PubMed]
- Jang, G.P.; Dolan, L. Auxin promotes the transition from chloronema to caulonema in moss protonema by positively regulating PpRSL1 and PpRSL2 in Physcomitrella patens. New Phytol. 2011, 192, 319–327. [Google Scholar] [CrossRef]
- Jang, G.; Yi, K.K.; Pires, N.D.; Menand, B.; Dolan, L. RSL genes are sufficient for rhizoid system development in early diverging land plants. Development 2011, 138, 2273–2281. [Google Scholar] [CrossRef]
- Proust, H.; Honkanen, S.; Jones, V.A.S.; Morieri, G.; Prescott, H.; Kelly, S.; Ishizaki, K.; Kohchi, T.; Dolan, L. RSL Class I Genes Controlled the Development of Epidermal Structures in the Common Ancestor of Land Plants. Curr. Biol. 2016, 26, 93–99. [Google Scholar] [CrossRef] [PubMed]
- Vijayakumar, P.; Datta, S.; Dolan, L. ROOT HAIR DEFECTIVE SIX-LIKE4 (RSL4) promotes root hair elongation by transcriptionally regulating the expression of genes required for cell growth. New Phytol. 2016, 212, 944–953. [Google Scholar] [CrossRef] [PubMed]
- Kim, C.M.; Han, C.D.; Dolan, L. RSL class I genes positively regulate root hair development in Oryza sativa. New Phytol. 2017, 213, 314–323. [Google Scholar] [CrossRef]
- Han, Y.; Xin, M.M.; Huang, K.; Xu, Y.Y.; Liu, Z.S.; Hu, Z.R.; Yao, Y.Y.; Peng, H.R.; Ni, Z.F.; Sun, Q.X. Altered expression of TaRSL4 gene by genome interplay shapes root hair length in allopolyploid wheat. New Phytol. 2016, 209, 721–732. [Google Scholar] [CrossRef]
- Kim, C.M.; Dolan, L. ROOT HAIR DEFECTIVE SIX-LIKE Class I Genes Promote Root Hair Development in the Grass. PLoS Genet. 2016, 12, e1006211. [Google Scholar] [CrossRef]
- Emms, D.M.; Kelly, S. OrthoFinder: Phylogenetic orthology inference for comparative genomics. Genome Biol. 2019, 20, 238. [Google Scholar] [CrossRef] [PubMed]
- Yan, F.; Zhu, Y.Y.; Müller, C.; Zörb, C.; Schubert, S. Adaptation of H+ -pumping and plasma membrane H+ ATPase activity in proteoid roots of white lupin under phosphate deficiency. Plant Physiol. 2002, 129, 50–63. [Google Scholar] [CrossRef]
- Wang, L.K.; Feng, Z.X.; Wang, X.; Wang, X.W.; Zhang, X.G. DEGseq: An R package for identifying differentially expressed genes from RNA-seq data. Bioinformatics 2010, 26, 136–138. [Google Scholar] [CrossRef] [PubMed]
- Gu, Z.G.; Eils, R.; Schlesner, M. Complex heatmaps reveal patterns and correlations in multidimensional genomic data. Bioinformatics 2016, 32, 2847–2849. [Google Scholar] [CrossRef]
- Schmutz, J.; Cannon, S.B.; Schlueter, J.; Ma, J.X.; Mitros, T.; Nelson, W.; Hyten, D.L.; Song, Q.J.; Thelen, J.J.; Cheng, J.L.; et al. Genome sequence of the palaeopolyploid soybean. Nature 2010, 463, 178–183. [Google Scholar] [CrossRef] [PubMed]
- Stai, J.S.; Yadav, A.; Sinou, C.; Bruneau, A.; Doyle, J.J.; Fernández-Baca, D.; Cannon, S.B. Cercis: A Non-polyploid Genomic Relic Within the Generally Polyploid Legume Family. Front. Plant Sci. 2019, 10, 345. [Google Scholar] [CrossRef]
- Zhuang, Y.B.; Wang, X.T.; Li, X.C.; Hu, J.M.; Fan, L.C.; Landis, J.B.; Cannon, S.B.; Grimwood, J.; Schmutz, J.; Jackson, S.A.; et al. Phylogenomics of the genus Glycine sheds light on polyploid evolution and life-strategy transition. Nat. Plants 2022, 8, 233. [Google Scholar] [CrossRef]
- Wang, L.F.; Jia, G.H.; Jiang, X.Y.; Cao, S.; Chen, Z.J.; Song, Q.X. Altered chromatin architecture and gene expression during polyploidization and domestication of soybean. Plant Cell 2021, 33, 1430–1446. [Google Scholar] [CrossRef]
- Fang, C.; Yang, M.Y.; Tang, Y.C.; Zhang, L.; Zhao, H.A.; Ni, H.J.; Chen, Q.S.; Meng, F.L.; Ae, J.J. Dynamics of cis-regulatory sequences and transcriptional divergence of duplicated genes in soybean. Proc. Natl. Acad. Sci. USA 2023, 120, e2303836120. [Google Scholar] [CrossRef]
- Lan, P.; Li, W.F.; Lin, W.D.; Santi, S.; Schmidt, W. Mapping gene activity of Arabidopsis root hairs. Genome Biol. 2013, 14, R67. [Google Scholar] [CrossRef]
- Deal, R.B.; Henikoff, S. A Simple Method for Gene Expression and Chromatin Profiling of Individual Cell Types within a Tissue. Dev. Cell 2010, 18, 1030–1040. [Google Scholar] [CrossRef] [PubMed]
- Stetter, M.G.; Schmid, K.; Ludewig, U. Uncovering Genes and Ploidy Involved in the High Diversity in Root Hair Density, Length and Response to Local Scarce Phosphate in Arabidopsis thaliana. PLoS ONE 2015, 10, e0120604. [Google Scholar] [CrossRef] [PubMed]
- Salazar-Henao, J.E.; Vélez-Bermúdez, I.C.; Schmidt, W. The regulation and plasticity of root hair patterning and morphogenesis. Development 2016, 143, 1848–1858. [Google Scholar] [CrossRef] [PubMed]
- Datta, S.; Prescott, H.; Dolan, L. Intensity of a pulse of RSL4 transcription factor synthesis determines Arabidopsis root hair cell size. Nat. Plants 2015, 1, 15138. [Google Scholar] [CrossRef] [PubMed]
- Lan, P.; Li, W.F.; Wen, T.N.; Schmidt, W. Quantitative Phosphoproteome Profiling of Iron-Deficient Arabidopsis Roots. Plant Physiol. 2012, 159, 403–417. [Google Scholar] [CrossRef] [PubMed]
- Menand, B.; Yi, K.K.; Jouannic, S.; Hoffmann, L.; Ryan, E.; Linstead, P.; Schaefer, D.G.; Dolan, L. An ancient mechanism controls the development of cells with a rooting function in land plants. Science 2007, 316, 1477–1480. [Google Scholar] [CrossRef]
- Mangano, S.; Denita-Juarez, S.P.; Marzol, E.; Borassi, C.; Estevez, J.M. High Auxin and High Phosphate Impact on RSL2 Expression and ROS-Homeostasis Linked to Root Hair Growth in Arabidopsis thaliana. Front. Plant Sci. 2018, 9, 1164. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| PUE Pathways | Arabidopsis | Lupinus | Pisum | Trifolium | Medicago | Glycine | Phaseolus | Vigna | Fabaceae |
|---|---|---|---|---|---|---|---|---|---|
| Glycolysis | 82 | 145 | 102 | 91 | 97 | 194 | 97 | 96 | 109 |
| Calvin cycle | 16 | 33 | 33 | 19 | 28 | 45 | 23 | 21 | 25 |
| TCA cycle for organic acid | 34 | 45 | 23 | 30 | 27 | 44 | 29 | 26 | 30 |
| CO2 fixation in non-photosynthetic | 20 | 50 | 42 | 31 | 42 | 64 | 36 | 39 | 40 |
| Lipid remodeling | 73 | 112 | 75 | 63 | 70 | 134 | 75 | 71 | 81 |
| Primary root development | 16 | 46 | 22 | 26 | 30 | 50 | 25 | 26 | 29 |
| Lateral root development | 21 | 83 | 65 | 65 | 65 | 139 | 66 | 71 | 73 |
| Root hair development | 50 | 264 | 188 | 182 | 211 | 371 | 218 | 212 | 226 |
| Acid phosphatase | 26 | 29 | 23 | 22 | 25 | 35 | 21 | 19 | 23 |
| Secreted Acid phosphatase | 2 | 8 | 3 | 4 | 3 | 5 | 3 | 3 | 4 |
| Secreted ribonuclease | 5 | 7 | 5 | 5 | 9 | 7 | 4 | 4 | 5 |
| Phosphate uptake | 9 | 12 | 10 | 7 | 11 | 14 | 9 | 8 | 10 |
| Phosphate transport | 25 | 35 | 30 | 23 | 25 | 48 | 24 | 25 | 28 |
| Mitochondrion electron transport | 10 | 23 | 19 | 16 | 19 | 31 | 19 | 18 | 19 |
| Proton transport | 24 | 115 | 78 | 103 | 120 | 111 | 94 | 89 | 90 |
| Sugar transport | 38 | 99 | 68 | 64 | 71 | 142 | 79 | 77 | 80 |
| Organic acid transporter | 70 | 73 | 78 | 59 | 75 | 129 | 69 | 70 | 76 |
| Other PUE genes | 164 | 1041 | 927 | 841 | 985 | 1453 | 902 | 853 | 940 |
| Total number | 685 | 2220 | 1791 | 1651 | 1913 | 3016 | 1793 | 1728 | 1888 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, X.; Yang, J.; Zhang, Q.; Zhang, L.; Cheng, F.; Xu, W. Phosphorus-Use-Efficiency Gene Identification in Fabaceae and RSL2 Expansion in Lupinus albus Is Associated with Low-Phosphorus Adaptation. Genes 2024, 15, 1049. https://doi.org/10.3390/genes15081049
Li X, Yang J, Zhang Q, Zhang L, Cheng F, Xu W. Phosphorus-Use-Efficiency Gene Identification in Fabaceae and RSL2 Expansion in Lupinus albus Is Associated with Low-Phosphorus Adaptation. Genes. 2024; 15(8):1049. https://doi.org/10.3390/genes15081049
Chicago/Turabian StyleLi, Xing, Jinyong Yang, Qian Zhang, Lingkui Zhang, Feng Cheng, and Weifeng Xu. 2024. "Phosphorus-Use-Efficiency Gene Identification in Fabaceae and RSL2 Expansion in Lupinus albus Is Associated with Low-Phosphorus Adaptation" Genes 15, no. 8: 1049. https://doi.org/10.3390/genes15081049