1. Introduction
Monitoring and characterizing the interacting effects of land use and climate on land surface processes is a primary focus of land change science [
1], and of particular concern in arid environments where both landscapes and livelihoods can be impacted by short-term climate variability [
2,
3]. A considerable amount of research conducted in the western United States documents and describes a wide range of anthropogenic and climate-driven vegetation changes over the past several centuries [
4,
5,
6,
7]. Much of the focus in the southwest has been on: (1) the conversion of native grassland to shrubland and savanna through the process of woody plant encroachment [
8,
9], and (2) changes in the distribution and cover of species-rich riparian vegetation related to ground and surface water use [
10,
11]. Grasslands and riparian systems share ecological and hydrologic linkages that are important for sustaining watershed function: grassland species composition, cover, and disturbance history will influence run-off, erosion, and groundwater recharge, ultimately affecting water availability for riparian vegetation [
12,
13,
14]. Spatial models describing past landscape changes to upland and riverine systems can provide important baseline information for restoration of arid watersheds.
Desert grasslands are of major conservation concern in temperate regions as they provide a number of ecological services ranging from livestock production, carbon storage, nitrogen cycling, rainfall infiltration, and habitat for animal biodiversity [
15,
16]. Many grasslands around the world are experiencing change through the encroachment of woody shrubs and trees, and there is currently no consensus concerning the exact causes of woody plant expansion into grasslands. In western North America, several key drivers have been identified including overgrazing, wildfire suppression, climate change, and recent CO
2 and N enrichment of the environment [
2,
7,
8,
9,
10,
11]. Prior to Anglo-American settlement, woody plants in grasslands were suppressed by periodic wildfires that helped to maintain range productivity and cycle nutrients [
7,
16,
17,
18]. Livestock grazing effectively suppressed grassland wildfires by consuming and disrupting the continuity of fine fuels [
5] and browsing helped to disperse and establish leguminous seeds of woody plants like mesquite (
Prosopis velutina) [
19]. Broad climate fluctuations during the 20th century, particularly periods of unusually high winter precipitation, are implicated in the preferential establishment of C3 woody shrubs over warm-season-active C4 grasses [
7,
20].
In addition to woody invasion of rangelands, riparian areas, including both densely forested riverine riparian corridors and narrow xeroriparian woodlands (woody plants with dense cover or interlocking canopies adjacent to ephemeral, dry washes) along tributaries, have undergone considerable anthropogenic and climate-driven changes since Anglo-American settlement in the southwest [
10,
11]. Like grasslands, change in western riparian vegetation is a topic of debate; but it is the amount and location of change rather than the drivers of change that are contested [
10]. Riparian areas provide valuable cultural, regulating, and provisioning services like flood control, biodiversity, water quality, scenic values, habitat corridors, and pollution buffers [
21,
22]. More than a century of livestock grazing, urban development, surface-water development, groundwater over extraction, and climate-related hydrological changes have influenced the spatial extent and quality of riparian vegetation in southeastern Arizona and northern Mexico [
10,
11,
23,
24]. These ecological changes to grasslands and riparian areas of the western US due to climatic fluctuations, water development, and livestock-related disturbances have been compounded by increasing conversion to rural and exurban land uses during the past 30–50 years, as the economic value of rangeland shifted from a pastoral economy to real-estate developments [
25,
26].
Restoration of degraded lands and adaptation to changing climate and land uses requires identification of the rates, patterns and drivers of vegetation change in both the temporal and spatial dimensions. Currently, many adaptive management models are informed by studies conducted using small plots within experimental ranges and/or protected areas, and therefore may have limited applicability for management of large, heterogeneous landscapes [
12,
27]. In addition to non-representative locations, ecological field experiments are often carried out within a short academic time frame, and the results, interpretations, and implications of these studies can become distorted by anomalous or extreme climate events occurring within longer-term climate patterns [
28,
29,
30]. Perhaps the most confounding limitation of many early rangeland studies was the failure to incorporate multiple geographic scales, patterns and processes into the research design [
31]. Bestlemeyer
et al. [
32] recently reviewed a number of rangeland studies that implemented State and Transition Models (STMs) and concluded that the lack of information describing spatial heterogeneity over the landscape is a major factor that severely limits the use of STMs for management. They believe that land use history and spatial feedbacks between adjacent vegetation patches, which have been traditionally ignored in the models, may affect transition narratives [
32], and multi-scale approaches to STMs will provide additional evidence to uncover spatial processes in vegetation change [
31,
33].
The objective of this study was to assess historical rates and spatial patterns of grassland and riparian change within a topographically complex watershed that contains a heterogeneous and bi-national land use mosaic. More specifically, our goals were to develop an integrative, multi-observational approach to landscape-change analysis that combines change detection methods from quantitative multitemporal (satellite imagery) and semi-quantitative long-term (repeat ground photography) data that would allow us to contextualize variability in the spatial and temporal dynamics of vegetation change relative to climate and land-use.
1.1. Historical Data
By providing documentation of ecosystem conditions in the past, replicated historical photographs are an important source of information on long-term biological and ecological changes in various regions and landscapes throughout the globe [
34,
35]. Both qualitative and quantitative change detection techniques have been applied to repeat photography to answer ecological research questions ranging from assessments of forest canopy changes to desert-plant population dynamics [
36,
37,
38,
39,
40,
41,
42]. Repeat photography has proven to be a particularly powerful tool for identifying long-term vegetation and ecosystem trends in the desert southwest [
7,
10].
The use of repeat photography for change analysis has several weaknesses, particularly when compared to more systematically collected, multispectral imagery from space-based platforms. For example, the oblique angle of ground photographs makes precise estimation of area (and consequently, vegetation cover) difficult, especially when the time of day and seasonality of the image collection can influence interpretation (but see [
41]). New and novel approaches to change detection using ground photography have been pursued using digital image processing and analysis [
43]. Complex image-processing techniques may ultimately offer no real advantage over visual interpretation if the objective is to simply assess if a certain vegetation type has increased, decreased, or remained unchanged. In many cases, visual interpretation of the photograph is sufficient to provide sound estimates of change in vegetation community types or percent cover [
44,
45].
Another obstacle in the interpretation and application of repeat photography is the non-random, spatial bias of the samples: many locations were originally selected for cultural, aesthetic, or commercial reasons [
46]. Original photo sites were often located along travel routes with heavy use or close to settlements, and the vegetation conditions depicted often contain signs of human use or disturbances that may not reflect the broader landscape. This limits the interpretation of original photography as a record of vegetation in its “natural state” or “pre-settlement” conditions. There are cases of non-biased samples, like those photographed in 1892 by D.R. Payne as part of the USA/Mexico Boundary Commission survey; the establishment of the US/Mexico border led them through remote parts of the desert southwest ([
4,
7] note: several of these photographs were used in this research).
1.2. Fusion of Historical and Contemporary Data
While presenting many challenges, the linkage of repeat photography with remotely-sensed images can strengthen the overall observational power of remote sensing by offering multiple lines of evidence as well as yielding information at multiple temporal and geographic scales. For example, historical photography extends the temporal envelope of remote-sensing analysis and provides useful ground-verification data for historical satellite imagery [
47]. Conversely satellite-platform remote sensing expands the spatial coverage and inferences made from a small sample of repeat photographs, putting the location of these long-term monitoring data into a broader context of landscape change albeit over a shorter time span. To date, very few studies have attempted to analyze long-term changes with both repeat ground photography and multitemporal satellite data or use historical photography to validate land-cover changes [
42].
Interpretation and analysis of both ecosystem monitoring data from remotely-sensed imagery and long-term field data allow us to quantify temporal changes over entire watersheds that include a wide range of plant assemblages, soil types, land uses, and land use histories. Thus far, land-change analyses in the region have been accomplished using either long-term monitoring data or remote sensing, both of which, when used on their own, have inherent limitations that weaken spatial and temporal inferences. In this paper, we analyze medium-term land-cover change in conjunction with long-term repeat photography at discrete locations. The temporal and spatial-scale issues addressed in this research are particularly important given the uncertainties surrounding vegetation change under predicted future warming, and the potential intensification of changes related to interactions with changing land use. We address these issues by examining change patterns in the context of long-term climate and land management. This long-term, spatially explicit information describing vegetation changes can help land stewards better manage, restore, and adapt in light of predicted future climate and land use and urban growth scenarios.
1.3. Study Area
The upper Santa Cruz Watershed occupies 9,146 km
2 of southern Arizona, USA, and northern Sonora, Mexico, with most of the watershed in the United States. The watershed contains two major urbanized areas: the greater Tucson metropolitan area (2011 population: 989,569) and “
Ambos Nogales,” a contiguous urban area that contains both Nogales, Sonora (2010 population: 220,292) and Nogales, Arizona (2011 population: 20,948) divided by the USA/Mexico border (
Figure 1). The remaining landscape within the watershed is predominantly rural and sparsely populated.
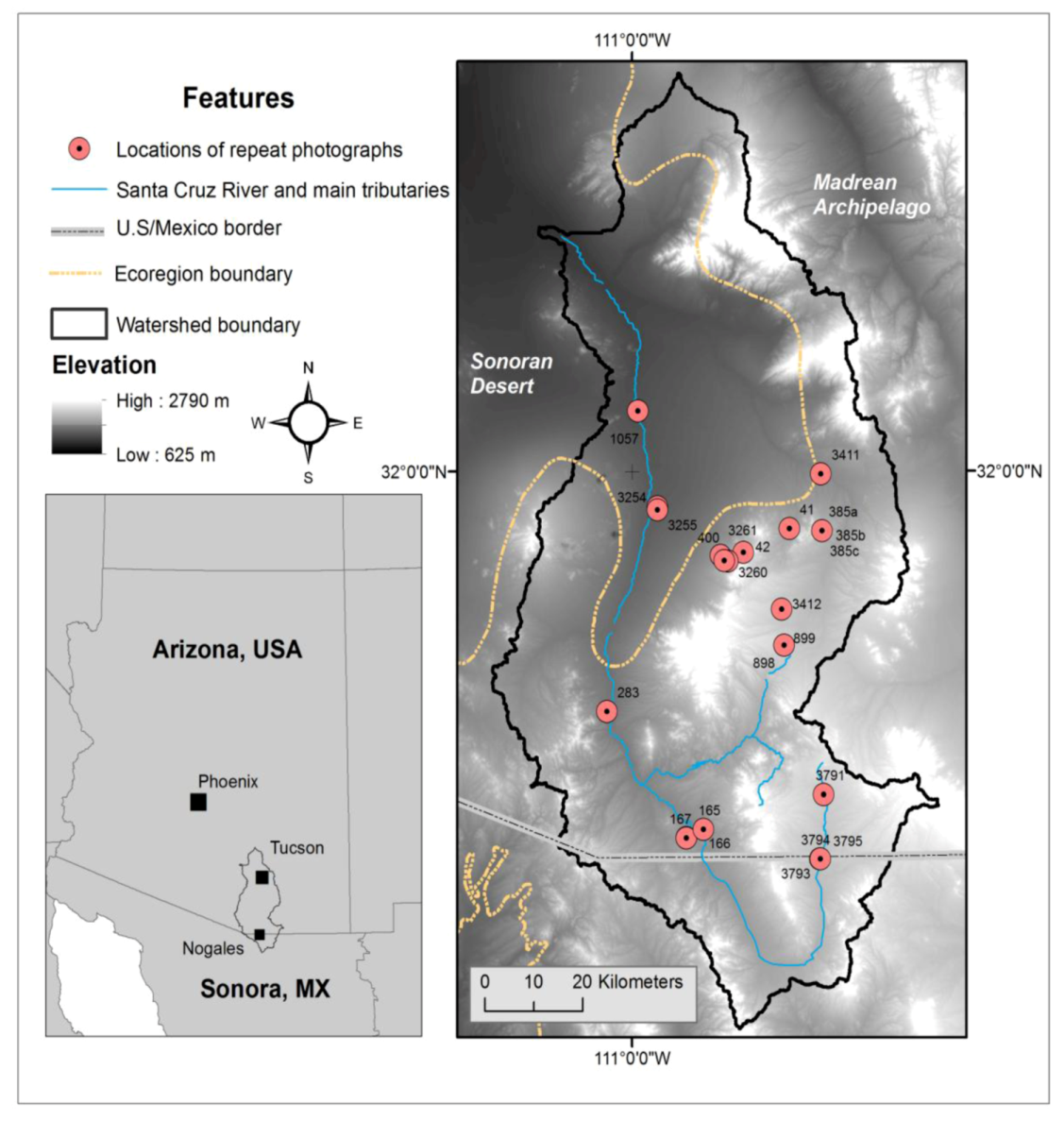
Figure 1.
Location of the Santa Cruz Watershed including Ecoregion boundaries, repeat photography camera stations, elevation, and the USA/Mexico boundary.
Figure 1.
Location of the Santa Cruz Watershed including Ecoregion boundaries, repeat photography camera stations, elevation, and the USA/Mexico boundary.
The topographically complex watershed ranges in elevation from 625 to 2,790 m and straddles the Sonoran Desert and the Madrean Archipelago Ecoregions [
48]. The greater Sonoran Desert Ecoregion, which covers arid portions of southeastern California and southwestern Arizona in the USA, and northeastern Baja California and northwestern Sonora in Mexico, is characterized by a dry subtropical desert climate and basin and range physiography, with elevations ranging from sea level to 1,400 m. The greater Madrean Archipelago Ecoregion spans southeast Arizona, southwest New Mexico in the USA, and northern Sonora in Mexico, and is characterized by a dry, subtropical to mid-latitude steppe climate and medium to high relief basin and range physiography with elevations ranging from 800–3,000 m. The separation between these two ecoregions in the Santa Cruz Watershed occurs at an elevation of approximately 1,000 m. Nearly 61% of the watershed is in the upper elevation Madrean Ecoregion; the lower basin, where the Tucson metropolitan area is situated, is primarily lower elevation Sonoran Desert. The watershed supports various plant associations and alliances ranging from Sonoran desert scrub to mixed-conifer forests.
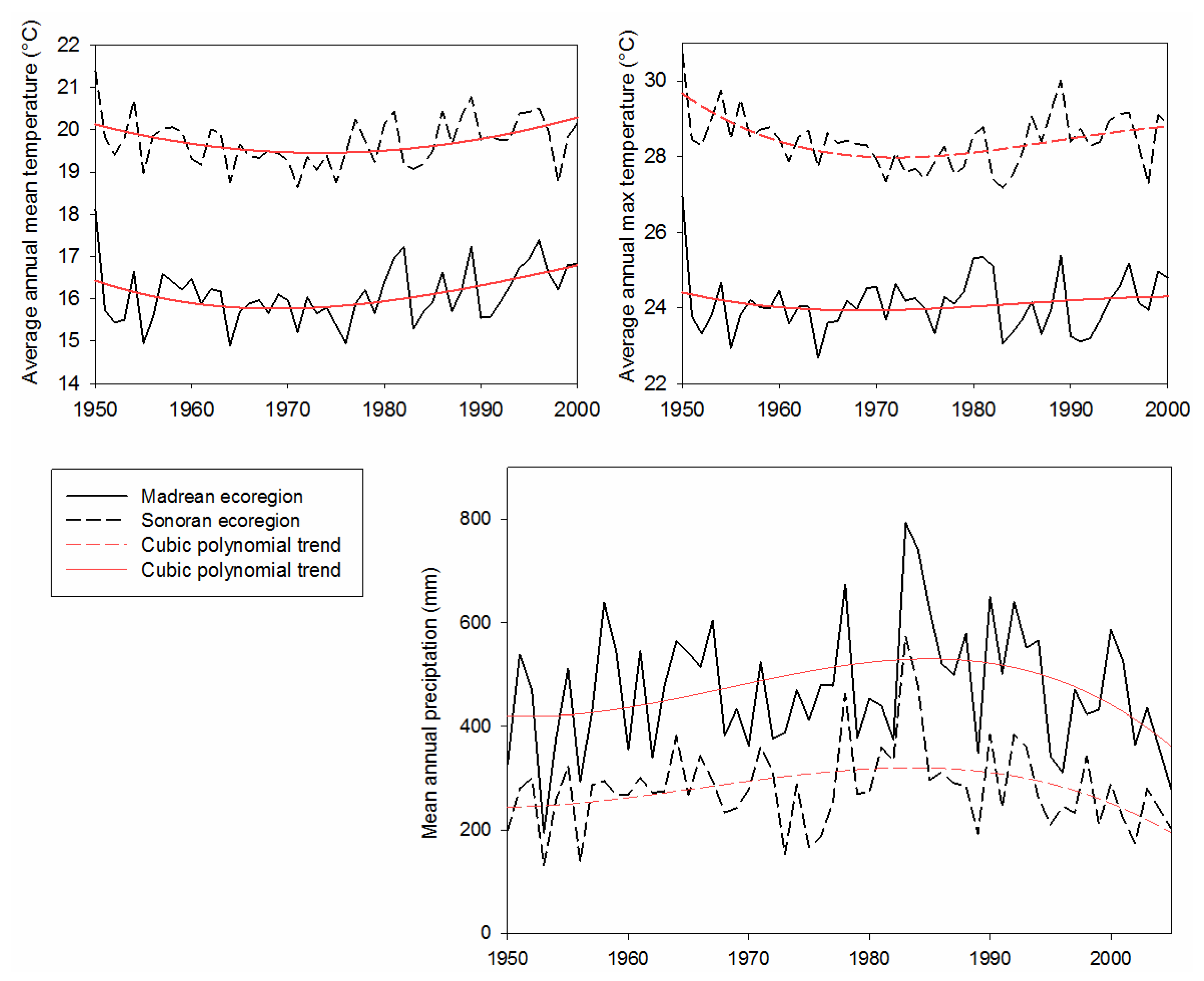
The climate of the watershed is characterized by mild winter and high summer temperatures and a bimodal precipitation pattern. Between 1950 and 2005, the minimum annual precipitation was 104 mm (measured at Anvil Ranch in 1956) and the maximum was 890 mm (measured at the Santa Rita Experimental Range in 1983). Average annual precipitation was 278 mm in the Sonoran Ecoregion and 470.5 mm in the Madrean. Mean temperature ranged from 16.09 °C in the Madrean to 19.71 °C in the Sonoran. Average monthly minimum temperature in the two Ecoregions ranged from 0.39 to −0.328 °C, and average maximum ranged from 32.44 °C to 37.83 °C.
1.4. Materials and Methods
We created a series of four (1979, 1989, 1999 and 2009) land use/land cover (LULC) maps to quantify and map land-cover change, acquired hardcopies and locations of historical, repeated ground photographs and rephotographed the locations to capture current day (2011–2012) conditions. The photographic time series were analyzed relative to landscape change patterns calculated from satellite data. Temporal and spatial landscape changes were then analyzed relative to broad historical climate patterns by ecoregion, and relative to fine-scale land ownership and management data. The following sections describe in detail the data used for analysis and the methods employed.
1.4.1. Multitemporal Land Cover
Primary spatial data were derived from four LULC maps with 14 cover classes mapped at decadal intervals (1979–2009; [
49]). The LULC classification scheme and classification techniques are based generally on methods used to develop the US Geological Survey (USGS) National Land Cover Database (NLCD) [
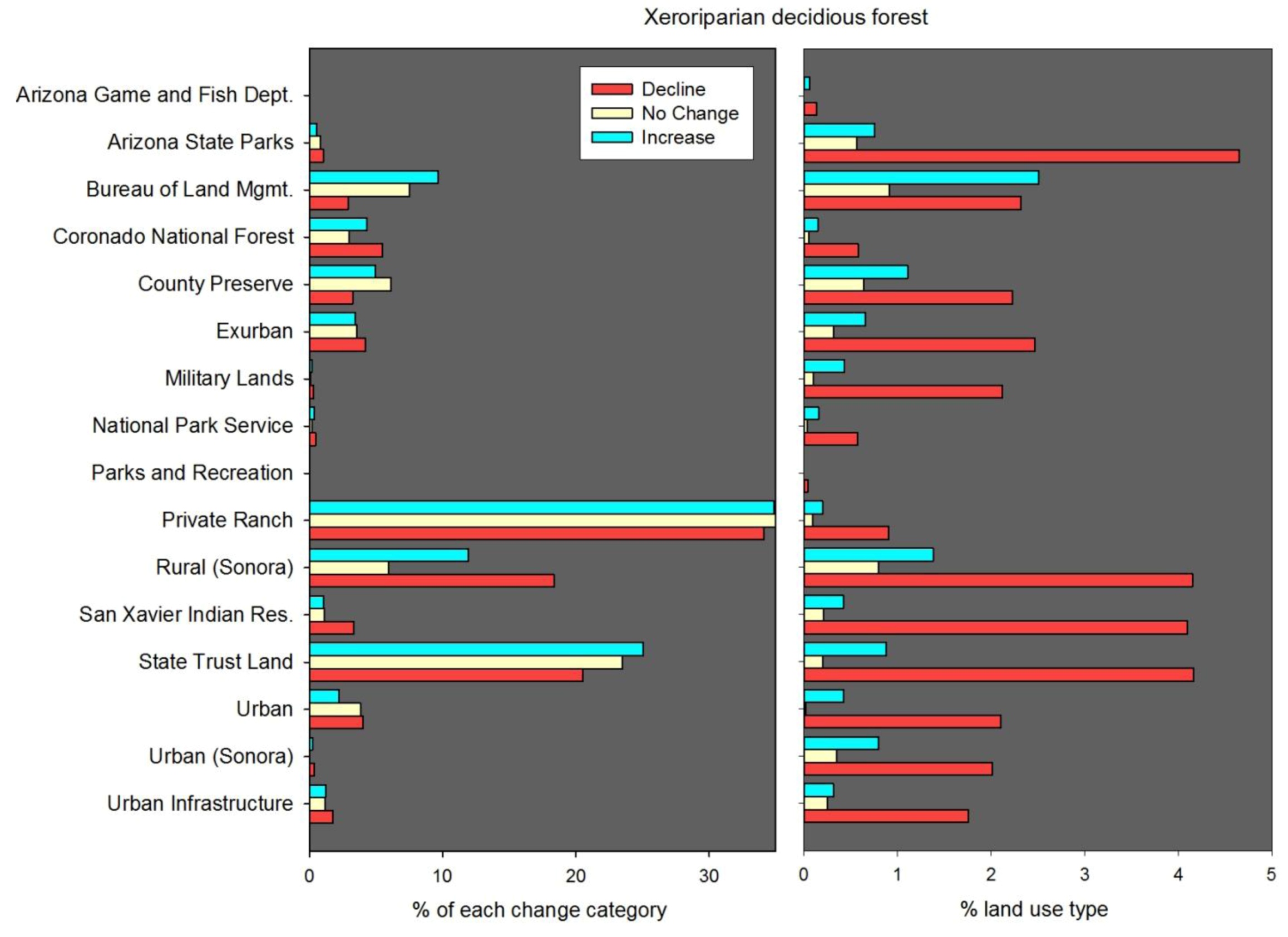
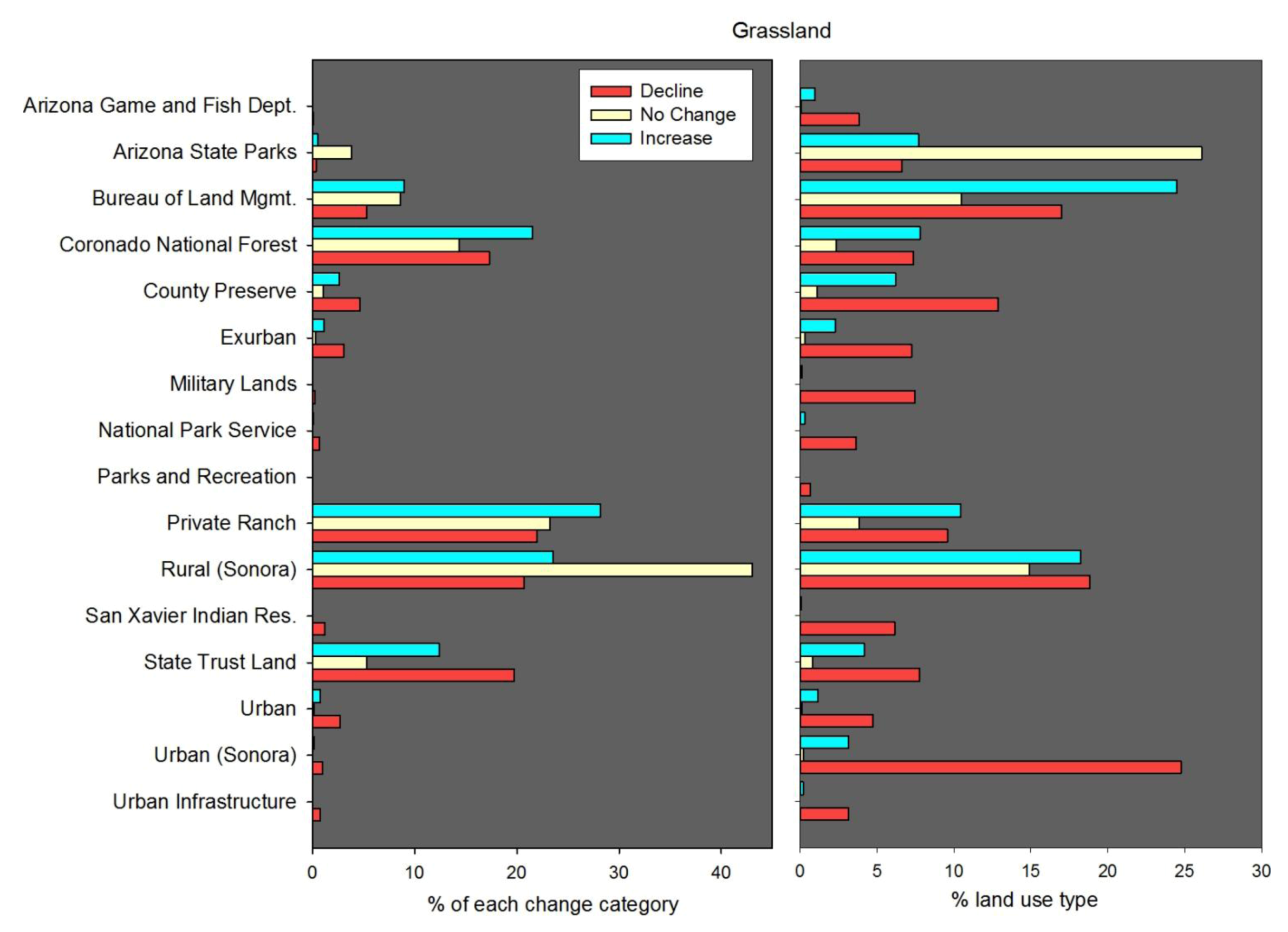
50]. There were some major, and deliberate, differences between our LULC dataset and existing NLCD maps. First, NLCD land cover was mapped for the conterminous United States only, whereas the LULC maps we developed of Santa Cruz Watershed included portions of Mexico. Second, the general NLCD classes we used were tailored to represent specific vegetation types that are of regional and local interest with an emphasis on accurately mapping riparian and xeroriparian classes, which were poorly characterized by NLCD in our study region. Although we mapped 14 classes of land cover, we prioritized our analysis on two classes of interest for rangeland conservation and restoration: grassland/herbaceous and xeroriparian deciduous forest (we classified xeroriparian and mesquite riparian
bosque (forest) communities as NLCD class “41 - Deciduous Forest”). Common woody plant species found in these xeroriparian communities include
Prosopis velutina, Acacia spp
., Parkinsonia florida, Ziziphus obtusifolia, and Lycium andersonii. Grasslands in the watershed are made up of mixed native grasses (
Bouteloua spp.,
Aristida spp.) and non-native grasses (e.g.,
Eragrostis lehmanniana), interspersed with large woody plants (e.g.,
Prosopis velutina,
Quercus spp.) desert shrubs (e.g.,
Acacia spp.,
Mimosa biuncifera), and cacti and succulents (e.g.,
Agave spp.
Opuntia spp.,
Yucca elata).
Landsat Multispectral Scanner (MSS) and Thematic Mapper (TM) bands, image-based transformations, and ancillary data sets were used to develop Classification and Regression Tree (CART) models. For each LULC classification we used two satellite images collected during seasonal extremes with the aim of differentiating land-cover classes based on changes in land surface phenology: one collected during the dry early-summer (May–June) and one after vegetation green-up following summer rains (August and September). The TM images were converted to reflectance values using the cosine of theta (COST) model [
51]. The 60 m MSS image was not converted to reflectance (however, this was not required as we made no direct spectral comparisons between years) but was scaled to 30 m. CART input variables included Landsat spectral bands, spectral transformation (including image variance and multitemporal Kauth Thomas [
52]) and ancillary environmental data (including slope and hydrology) (see [
49] for detailed methods).
CART is a non-parametric model that has proven useful in applications that involve both multispectral and ancillary data [
53]. The CART model recursively splits multidimensional data into a dichotomous classification tree based on statistical characteristics of the training data. In this case the CART classifications were trained using reference data from high-resolution aerial photographs collected on or near the date of the Landsat imagery. For example, training areas for the 1979 map were digitized from three dates of aerial photographs (1975 panchromatic, 1980 panchromatic, and 1984 color infrared imagery). Training classes were discriminated based on visual interpretation (
i.e., deciduous and evergreen tree cover is determined by color and texture of tree canopy structure). When possible the same training sites were used to classify all four dates of imagery (if the training sample locations experienced no vegetative change over time period). By developing decision-tree models for the different dates of satellite imagery using this invariant sampling approach, we reduce classification errors associated with potential spatial and temporal variability of land-cover training samples and spectral data.
Land-cover classes were assigned to a training area based on visual interpretation of percentage of cover types present in the sample area and identification of the dominant cover type. For example, if an area had a mixture of cover types, grassland was selected over a shrubland if the dominant cover type in the area was herbaceous, rather than tree, shrub, or bare soil. This cover-based approach to classifier training theoretically allows for hard classification of subtle spectral shifts in community composition over time. If, for example, at time t, a 5% per decade shrub encroachment occurred in a pixel that was at t
-1 dominated by 42% herbaceous cover, 40% bare soil, and 18% shrub cover, then that pixel could in theory be classified at t
-2 as bare soil or remain herbaceous, depending on which cover type the shrubs replaced. This approach makes the models more flexible, but also makes change interpretation more difficult. In this example, depending on what the output class was, we are not certain whether woody plant encroachment occurred or if the land had become desertified. Villarreal
et al. [
49] reports land-cover class accuracies, kappa statistics, and analysis of errors. A condensed version of this information, including overall map accuracies, kappas and accuracies of our two classes of interest are presented in
Table 1.
Table 1.
Classification accuracy of land cover data used in this study.
Table 1.
Classification accuracy of land cover data used in this study.
| Date | Land Cover Maps | Deciduous Forest | Grassland |
| Overall Accuracy (%) | Kappa | Users (%) | Producers (%) | Users (%) | Producers (%) |
| 1979 | 84.8 | 0.835 | 90 | 85 | 78 | 89 |
| 1989 | 84.3 | 0.829 | 92 | 80 | 74 | 65 |
| 1999 | 81.8 | 0.802 | 78 | 81 | 86 | 86 |
| 2009 | 86.5 | 0.854 | 86 | 84 | 83 | 80 |
1.4.2. Post-Classification Change Detection
Detecting temporal changes in land cover or vegetation using remotely sensed imagery is a challenging task and the focus of considerable research [
54,
55,
56]. There are numerous change detection techniques available, each with advantages and disadvantages [
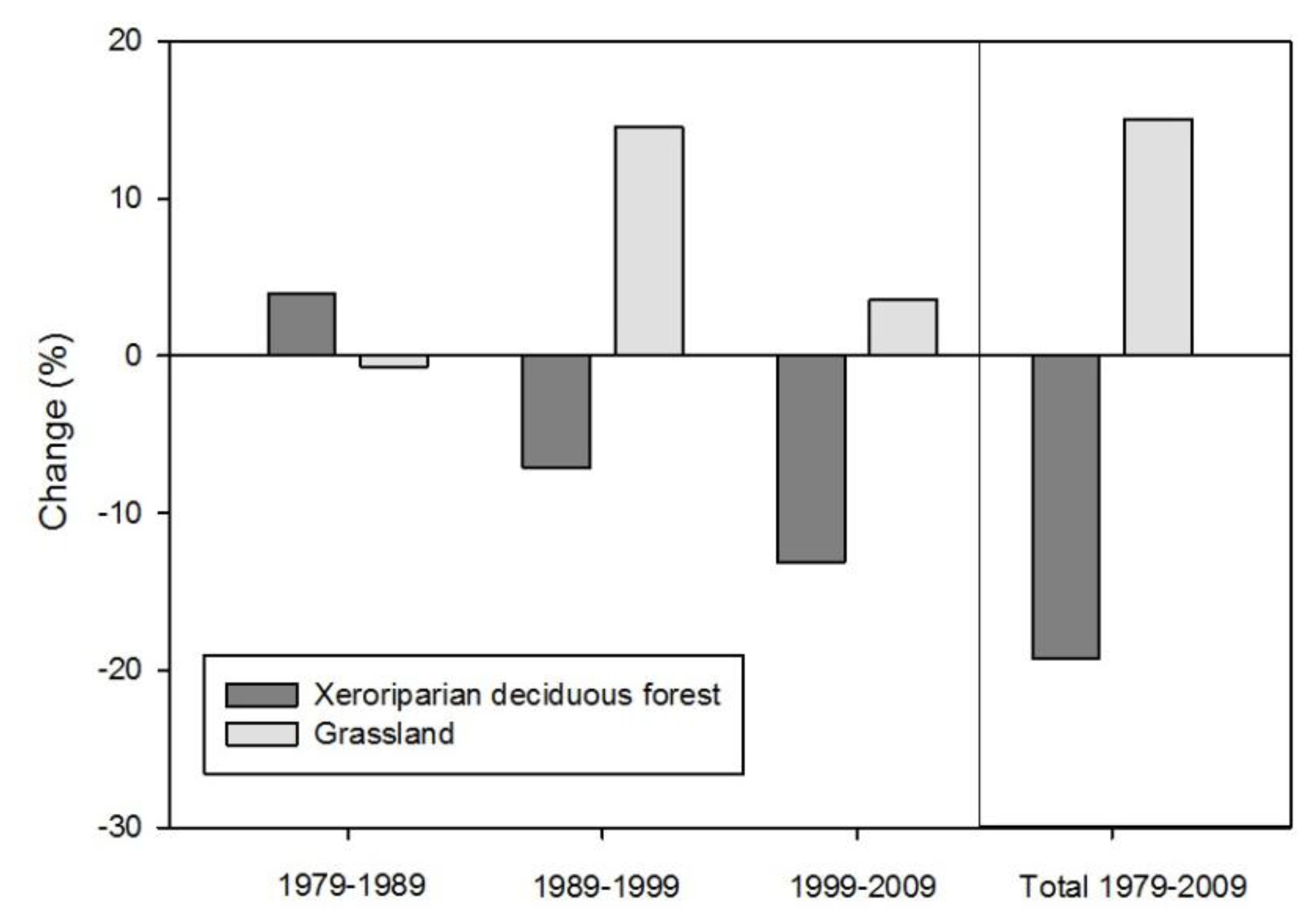
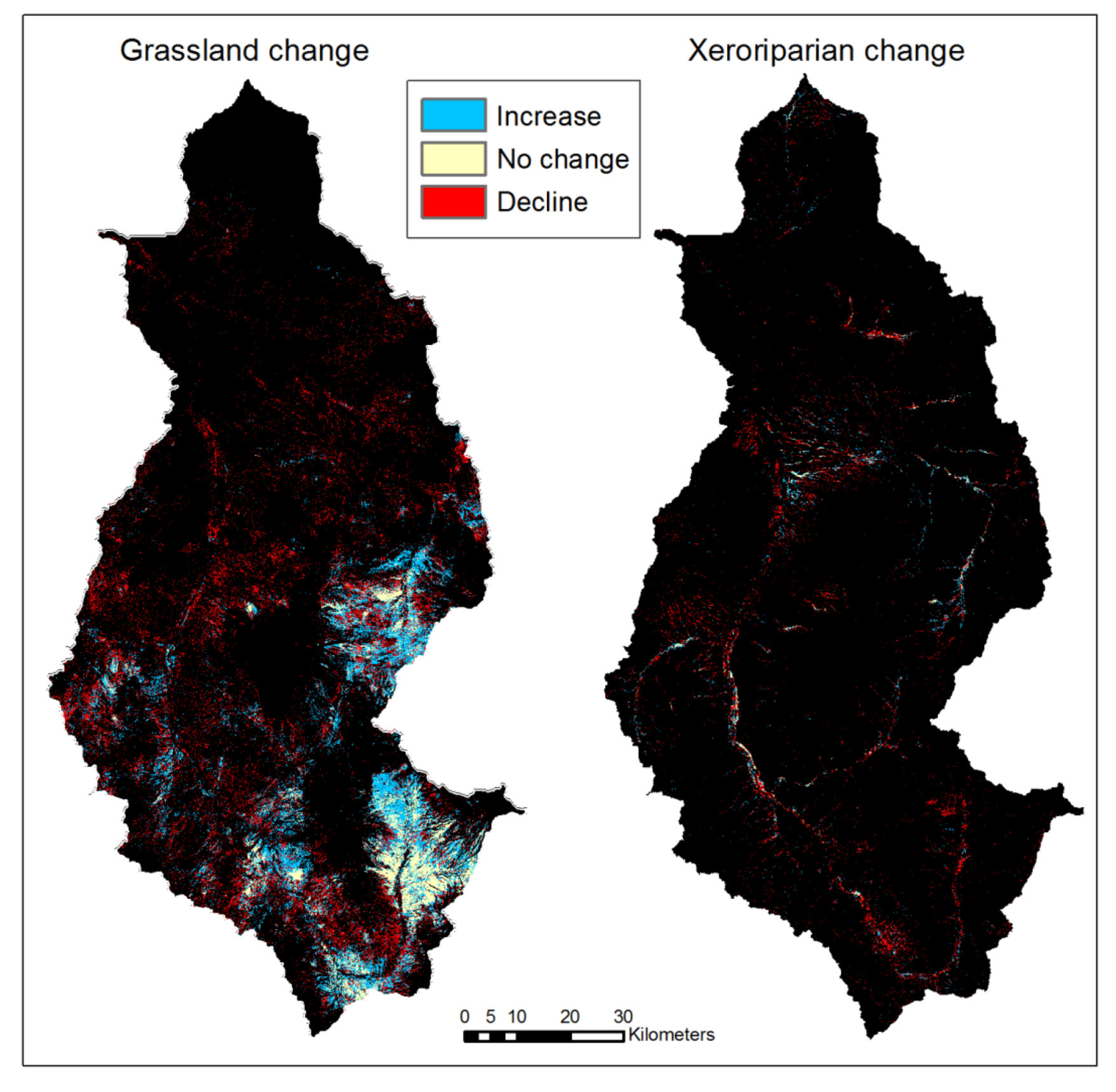
57]. The choice of change detection technique is typically informed by the specific research question, satellite data parameters, and temporal range of the analysis. For this study we created post-classification change detection maps by executing map algebra expression on the four dates of land cover. We chose post-classification change detection because we were interested in mapping and assessing changes in area and spatial distribution of specific land cover types. A major drawback of post-classification change detection is the number of potential change combinations resulting from the number of classes (14) and number of raster matrices (4). Because of the potentially large number of change combinations between the four dates we developed a set of criteria to categorize change classes as either “increase,” “no change,” or “decline.” For example, the “grassland increase” category included pixels mapped as grassland (1) only in 1999 or 2009, (2) in 1979 and again in 1999 or 2009, (3) in 1989 and again in 2009, and (4) in 1999 and 2009. The “grassland no change” category included pixels mapped as grassland in (1) all years, as well as (2) 1979, 1999, and 2009, and (3) 1989, 1999, and 2009. The “grassland decline” category included pixels mapped as grassland (1) only in 1979 but not after, (2) only in 1989 but not after, and (3) in both 1979 and 1989 but not in subsequent years. The xeroriparian deciduous forest change map was classified using the same criteria described above.
1.4.3. Repeat Photography
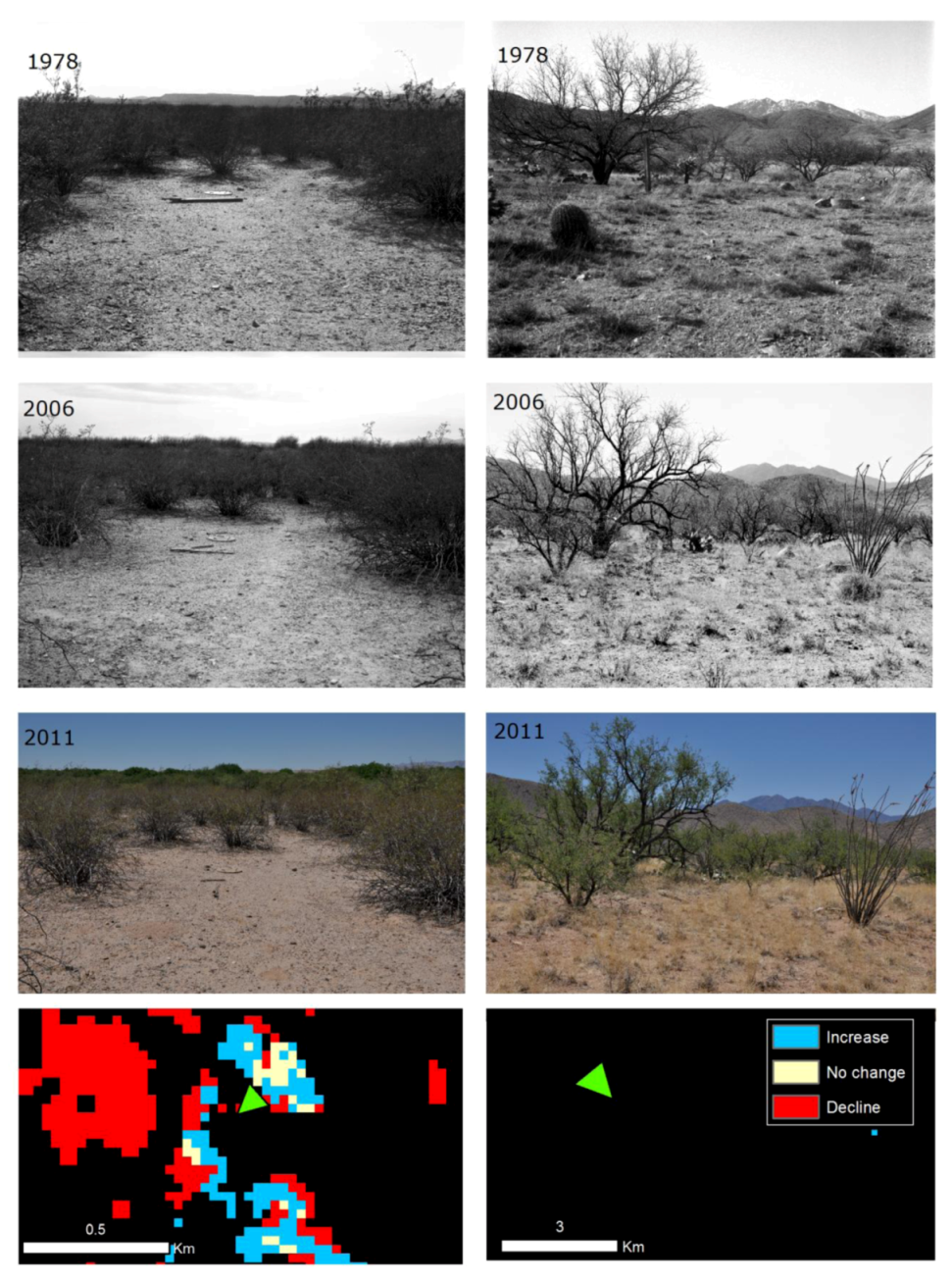
We identified georeferenced (Latitude/Longitude) locations of repeat ground photography in the Santa Cruz watershed from the US Geological Survey Desert Laboratory Collection of Repeat Photography [
58]. We identified photographic series with coordinates that overlapped mapped xeroriparian or grassland change classes. In addition, we identified and selected photographs located in areas that exhibited no change but depicted desert grassland and/or mesquite cover types. We revisited and rephotographed the sites if the most recent photograph was more than 5 years old. We ultimately rephotographed and qualitatively analyzed vegetation change at 24 camera stations (
Figure 1). Repeat photographs were taken using both a 4 × 5 film camera with black and white and color film, and a digital SLR. Film negatives were scanned and corrected and the digital files were matched with original photographs using Adobe Photoshop. We assigned change classes to each series of repeat photographs for both long-term historical trends and satellite-era trends (
i.e., [
59]). Geographic locations and camera direction were overlaid on Landsat-derived change maps and the changes were analyzed according to the landscape area and view angel depicted in the photographs.
1.4.4. Land Ownership and Ecoregion
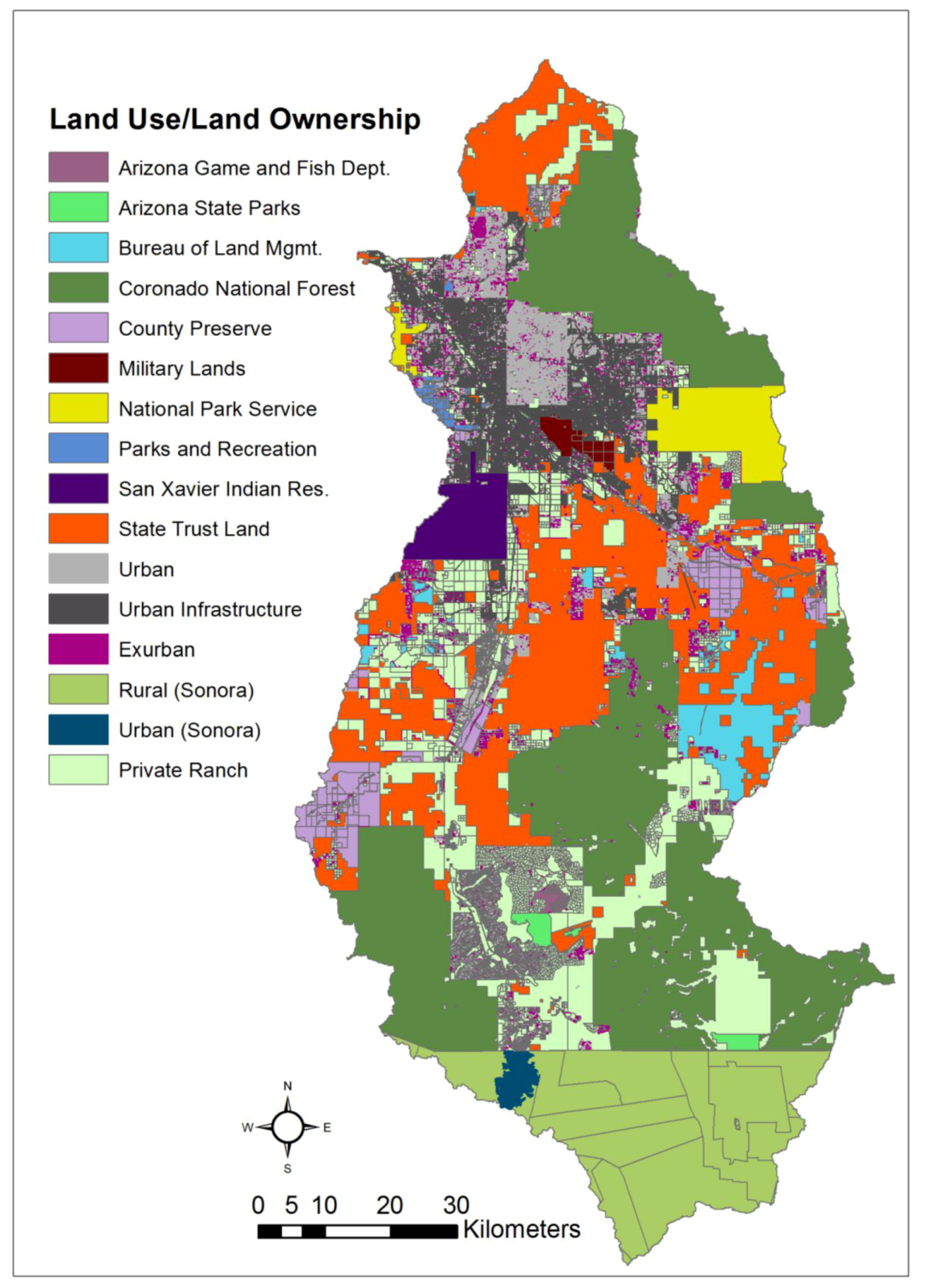
We analyzed landscape change and photography data within (1) units of land management and ownership, and by (2) ecoregions, that is, biogeographic units that contain similar long-term climate, vegetation assemblages, and landforms. We assembled a binational land-ownership dataset from multiple sources with varying degrees of categorical detail: Arizona State Land Department Public Land Ownership, Santa Cruz County Parcels, Pima County Preserves, Pima County Parcels, Nogales Mexico
Carta Urbana (Urban Map), and Integrated Mexican Basic Geo-Statistical Areas (Mexican Census boundaries). Privately-owned parcels from the United States part of the watershed were classified into 3 subgroups based on size: (1) <1.69 ha (Urban: high density urban residential to suburban ranch), (2) 1.7–6.7 ha (Exurban: rural homesteads – guest ranch and lodges), and (3) >6.8 ha (Ranch: low density rural -working ranches). These subgroups were based on Land Use Code zoning information for the City of Tucson, Pima County, and Santa Cruz County. Land in Sonora, Mexico, was divided into two categories: (1) urban (as defined in the
Carta Urbana), and (2) rural, from the Mexican Census. The rural-land boundaries roughly delineate large
ejidos, or communal lands used for ranching and agriculture. The main ownership types, as well as their descriptions and distributions, are reported in
Table 2 and
Figure 2.
Table 2.
Descriptive statistics of land use/land ownership parcel data.
Table 2.
Descriptive statistics of land use/land ownership parcel data.
| Land Use/Land Ownership | Description | Area (ha) | # of units | Mean (ha) |
|---|
| AZ Game and Fish Dept. | Managed by Arizona Game and Fish Dept. | 1,498 | 67 | 22.4 |
| AZ State Parks | Parklands managed by the State of Arizona. | 3,819 | 56 | 68.2 |
| Bureau of Land Mgmt. | Uses range from ranching to conservation. | 21,359 | 276 | 65.2 |
| Coronado National Forest | Managed by US National Forest Service. | 190,175 | 466 | 451.9 |
| County Preserve | Ranches and open space purchased for conservation. | 24,495 | 440 | 49.1 |
| Exurban | 1.7–6.7 ha. Uses include rural homesteads and guest ranches. | 33,882 | 11,754 | 2.9 |
| Military Lands | Department of Defense lands. | 4,351 | 59 | 65.9 |
| National Park Service | Managed by US National Park Service. | 23,332 | 3 | 507.4 |
| Parks and Recreation | Parklands managed by Pima and Santa Cruz counties. | 2,799 | 142 | 18.6 |
| Private Ranch | > 6.8 ha. Uses include cattle grazing and agriculture. | 175,475 | 3,987 | 39.5 |
| Rural (Sonora) | Communal ejido lands. Uses include grazing and agriculture. | 98,714 | 27 | 3,655.0 |
| San Xavier Indian Res. | Managed by the Tohono O'odham Nation. | 15,447 | 8 | 1,931.0 |
| State Trust Land | AZ undeveloped lands auctioned to provide Trust revenue. | 175,685 | 641 | 193.6 |
| Urban | < 1.7 ha. High density urban to suburban ranch uses. | 53,946 | 127,737 | 0.4 |
| Urban (Sonora) | High density urban lands in and around Nogales, Sonora. | 3,749 | 143 | 26.2 |
| Urban Infrastructure | Road networks, freeways, airports. | 32,406 | 16 | 3,150.7 |
Figure 2.
Distribution of land management and land-use units over the study area.
Figure 2.
Distribution of land management and land-use units over the study area.
We masked out land-cover classes that were not subject to change to reduce the likelihood of skewing change values relative to total watershed area and management-unit area. A mask was created from the initial 1979 land-cover map using only classes that had a low likelihood of transitioning to grassland or xeroriparian deciduous forests. These classes include Developed, Open Space; Developed, Low Intensity; Developed, Medium Intensity; Developed, High Intensity; Barren Land (Rock/Sand/Clay); and Evergreen Forest. The mask was overlain to block irrelevant data from analysis, allowing us to calculate a more realistic figure of total area subject to change within each land use/ownership type.
We analyzed long-term measurements from four climate stations that were split between the two ecoregions (Sonoran = Anvil Ranch, Station #0287; Tucson, Station #8820; Madrean = Santa Rita Experimental Range, Station #7593; and Canelo, Station #1231). Average-annual climate for each ecoregion was calculated from the two stations, which were selected to roughly represent the upper- and lower-elevational distribution of the grassland and xeroriparian vegetation classes. Climate variables include monthly mean precipitation (1950–2005), mean monthly temperature (1950–2000), and the average maximum monthly temperature (1950–2000). The non-parametric Mann-Whitney Rank Sum test was used to test for differences in precipitation between ecoregions, and t-tests for temperature variables. Land-cover and repeat-photography data were stratified by ecoregion to analyze and understand the potential influence of climate on observed vegetation changes.




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}











