1. Introduction
Heteroptera, or true bugs, are a widely distributed, highly diverse insect taxon with approximately 45,300 [
1] species worldwide. They populate every climate zone from arctic to desert [
2,
3,
4], showing great ability to cope with a variety of environmental factors. This leads to a substantial potential for invasiveness, and indeed many bug species are known to be highly invasive. For example,
Halyomorpha halys, an Asian stink bug invading Europe and North America, made it to the top four “most wanted” species list in the USA, inflicting economic damage of USD 37 million to tree fruit production in 2010 [
5]. However, although some species thrive over wide latitudinal ranges, other closely related species are more restricted in their distribution range (e.g.,
Orsillus depressus: Southern and Central Europe, northwards until Scandinavia;
O. maculatus: Southern Europe, around the Mediterranean and Black Sea) [
6]. Species such as
Oxycarenus lavaterae are known to have spread from their Mediterranean origins rather recently and are on their way north- and eastwards [
7,
8,
9,
10,
11]. They seem to benefit from climate change, with its higher annual mean temperatures, as well as human behavior, which facilitates dispersion (often along traffic routes) and settlement (e.g., through extensive plantings of
Tilia trees, a host plant of
Oxycarenus in urban environments).
Temperature is a main parameter for thriving and spread of ectothermic insects [
12,
13,
14]. Within the insects’ favorable temperature zone, it determines development and reproduction. Temperatures outside of this favorable zone hinder and become lethal at a certain point [
15]. In temperate climate regions, the change from favorable to unfavorable is mainly a seasonal occurrence, with low temperatures during the winter and possible high temperature extremes during summer. Adaptations such as hibernation or winter diapause, and aestivation lets the organisms cope with these adverse environmental conditions.
The climate variability hypothesis states that organisms exposed to higher variations in their thermal environment show more tolerance to extreme temperatures [
16]. In this regard, the organisms’ critical thermal maxima show less variation than their critical thermal minima [
17,
18,
19]. Geographical latitude, seasonality in temperature, and also elevation above sea level are thought to correlate with the animals’ temperature tolerance breadth and be a valid measurement for climate variability (see for instance [
18,
20,
21,
22,
23]).
Several attempts have been made to assess and model the distribution of ectotherms and endotherms in connection with thermal tolerance and macro-ecological scales such as climate parameters and latitude [
18,
20,
24,
25,
26,
27,
28]. One problem correlating thermal traits of animals with climatic measurements are the differences in scale (climate data point = animal size × 10,000 [
29]). Contradictory hypotheses regarding the species’ susceptibility to climate warming (e.g., [
30] vs. [
31]) show that large-scale climate-based studies might be insufficient to describe and predict biological effects. The assessment of microclimatic habitats as buffers of environmental conditions seems to be more suitable. However, even microclimate measurements do not always show the temperatures the animals are exposed to [
32,
33], because in reaction to uncomfortable conditions the animals show various physiological (e.g., evaporation [
34,
35,
36]) and/or behavioral reactions (e.g., by burrowing or simply moving away [
37,
38]). Such micro-scale measurements, however, are often not feasible, or data are not accessible. For medium to large scale assessment of physiological and behavioral factors driving insect distribution and dispersion, therefore, standard meteorological data and their bioclimatic classifications are often the only accessible data source to study physiological and behavioral responses in order to gain a deeper understanding of the underlying mechanisms of animal survival under extreme environmental conditions [
39]. Critical thermal limits are commonly used physiological parameters to assess factors driving insect dispersion and distribution. One way to describe upper and lower thermal limits of organisms is the definition of temperature when voluntary, coordinated movement stops [
40,
41,
42], rendering the animal finally unable to react to an adverse environment via locomotion (i.e., moving away to another, more habitable part of its surroundings).
The aim of this study was the evaluation of thermal tolerance traits (critical thermal maximum and minimum) of some common bugs from the temperate central Europe and the neighboring Mediterranean climate zones. The data were analyzed in relation to climatological and geographical parameters, as well as phylogenetic traits. With this knowledge, we wanted to test whether the climate variability hypothesis is applicable for this insect group, and whether their thermal traits are the basics for their high potential of invasiveness.
4. Discussion
True bugs are an abundant and successful insect group, capable of populating a wide variety of habitats. The species covered in this paper are closely related, of the families Lygaeidae and Pyrrhocoridae, and colonized four Köppen–Geiger climate regions over a latitudinal range of almost 6°.
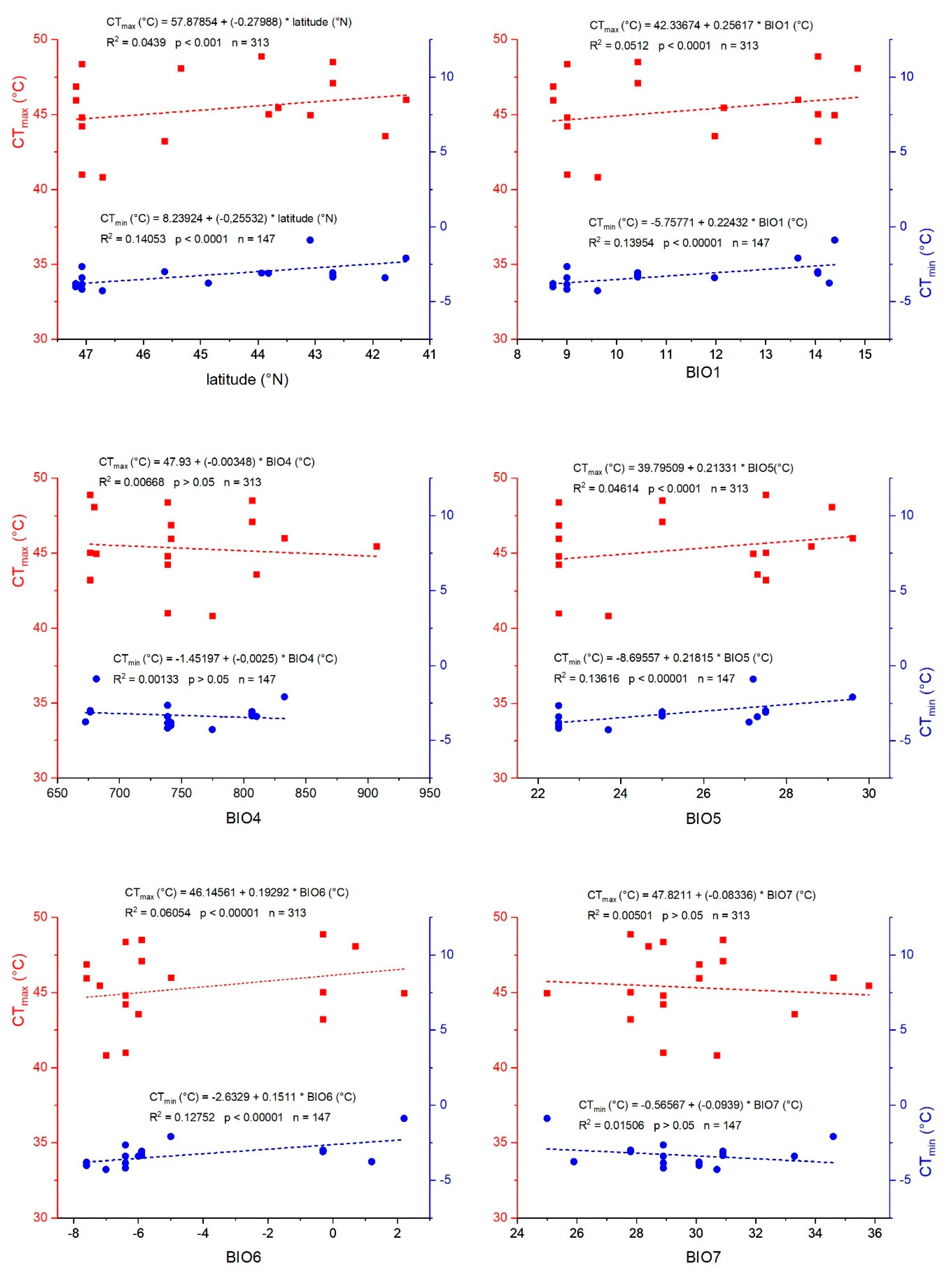
Upper thermal limits in the tested bug species varied stronger (SD = 2.5 °C, range = 39.4–49.5 °C; total mean CT
max = 45.3 °C) than lower limits (SD = 1.4 °C, range = −4.6–2.1 °C; mean CT
min = −3.3 °C) (
p < 0.001,
F-test; see
Table 2). Regarding the upper limits, this concurs with the findings of Kellermann et al. ([
57], on fruit flies). However, the ability to withstand high temperatures (CT
max) declines with an improved tolerance to low temperatures (CT
min) in dung beetles [
19]. In our seed bugs, CT
max correlated significantly with all studied climatic parameters (
Table 3), except BIO7 (a derived bioclimatic parameter: BIO7 = BIO5 − BIO6), despite the moderate variation in latitude (6°) and thus climate variability. This finding demonstrates that climate has a strong impact on seed bug heat tolerance. Only temperature seasonality (BIO4) and minimum temperature of the coldest month (BIO6) had a less pronounced (though significant) effect on CT
max. Besides bioclimatic parameters, mass showed a strong positive effect on CT
max (
Table 3,
Figure S3). This coincides with findings in beetles [
58,
59]. In other insects, however, results are not consistent. In ants, a positive correlation was reported [
60,
61] but also a decrease with mass [
62,
63]. In termites, a positive [
64] or no correlation was reported [
65].
Concerning the ability to tolerate cold, by contrast, the results are more diverse (
Table 3). Although CT
min is influenced by bioclimatic variables in general, only some bioclimatic variables seemed to have a direct effect, that is, the minimum temperature of the coldest month (BIO6) and associated parameters (mean diurnal range, BIO2; isothermality, BIO3). This seems plausible, because minimum environmental temperatures strongly determine survival during cold seasons, when behavioral avoidance is restricted, and thus the evolutionary drive to withstand them by physiological adaptation is high. If one accepts this interpretation, it seems intelligible that the annual mean temperature (BIO1) has only a weak effect on CT
min. Mass had no effect on the CT
min of seed bugs (
Table 3,
Figure S3). By contrast, in ants, a decrease with mass was reported [
63]. In a seasonal comparison of two termite species, no effect of mass on CT
min was reported [
65]. Raschmanová et al. [
66] observed a significant decrease of cold resistance with increasing body length in Collembola species inhabiting soil and subterranean habitats.
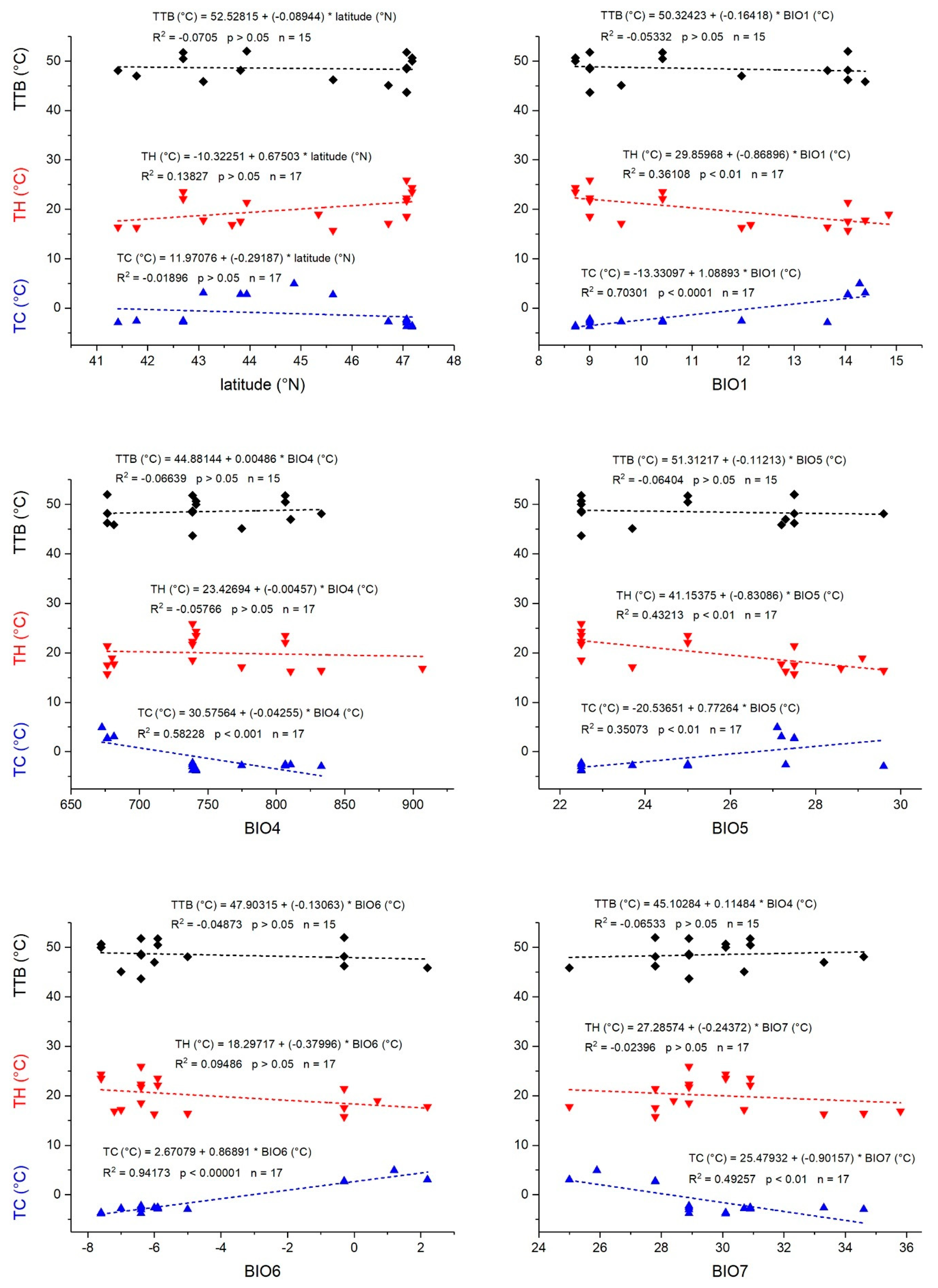
Responses to temperatures below the favorable temperature range (i.e., cold stress) often rely on different physiological mechanisms than those to heat stress. They are decoupled evolutionarily, and therefore may change differently in the course of the species’ colonization of new areas [
67]. Our findings support the hypothesis of decoupled thermal limits. Although cold tolerance (TC) correlated significantly with several bioclimatic parameters in a GLM analysis, heat tolerance (TH) did not (
Table 4). This is in good accordance with a study of Addo-Bediako et al. [
20], who found that upper thermal limits show little geographical variation, but the lower bounds of supercooling points and lower lethal temperatures do decline with latitude (and thus minimum temperature). In individual regression analyses (where other bioclimatic parameters were not included as covariates), cold tolerance (TC) depended on all tested bioclimatic parameters (
Figure 3). On the one hand, it is plausible that with decreasing annual mean temperature (BIO1) and the minimum temperature of the coldest month (BIO6), the bugs adapted to endure lower temperatures (have a lower CT
min;
Figure 2). On the other hand, the observed negative values of cold tolerance (TC) under cold conditions (minimum temperature of the coldest month, BIO6;
Figure 3) would indicate that the bugs were below their safe thermal conditions. This suggests that they seek out more favorable microclimatic environments in good time, that is, at higher ambient temperatures where unhindered mobility is guaranteed. The temperature in these microclimatic environments may well be higher than indicated by the macroclimatic variables [
68]. By contrast, heat tolerance TH, which represents the difference of CT
max to the maximum temperature of the warmest month (BIO5), depended only on the annual mean temperature (BIO1) and the maximum temperature of the warmest month (BIO5) (
Figure 3,
Table S2). A decreasing heat tolerance with these bioclimatic variables (BIO1, BIO5) indicates a decrease of the upper thermal safety margin (compare
Figure 4). It remains unclear, however, whether this occurs because of physiological restraints (i.e., inability for a higher CT
max) or a lack of need to increase CT
max further (because the thermal safety range is already sufficient).
Both cold and heat tolerance (TC, TH) did not correlate with latitude (
Figure 3). This could have been due to the narrow latitudinal range of 6°, as already mentioned above. Our results demonstrated that the investigation of variables directly affecting the studied organisms, instead of surrogate variables such as latitude or elevation, should increase the ability to understand the mechanisms driving animal distribution and biodiversity (see also [
69]).
It has to be kept in mind, however, that our analysis does not include all ecological factors relevant for survival of unfavorable conditions. Mean minimum temperatures of the coldest month (BIO6) in Austria and Bulgaria were lower than the species’ CT
min, resulting in negative values of cold tolerance (TC) (
Figure 4). Here, the overwintering individuals have to seek out microhabitats that provide a more benign microclimate, wherein they can endure cold periods [
68,
70,
71]. However, if endured winter temperatures sink under the lethal temperatures of the bug species, colonization of such regions is not possible. This might explain the absence of, for instance, the Mediterranean
Orsillus maculatus in colder regions of Europe, whereas
Orsillus depressus thrives in these regions [
43].
Heat tolerance (TH) in all species exceeded mean maximum temperatures of the warmest month (BIO5) in all locations (
Table 2; by up to 25.9 °C in
Oxycarenus lavaterae in Graz/AT), which coincides with the findings of Sunday et al. [
24] in several ectotherms. As all individuals were sampled during the cold season (winter), an immediate acclimation to high temperatures is unlikely. However, as bioclimatic parameters were the climatological normal from 1970 to 2000, absolute maximum temperatures were likely to be higher at some times, but not to the extent that they make permanent populations impossible.
The climate variability hypothesis states that a positive relationship exists between the breadth of thermal tolerance (TTB, the degree of eurythermy) and the level of climatic variability experienced by taxa with increasing latitude, especially in terrestrial ectotherms [
16,
72]. However, on the basis of our results, we could not make a clear decision because there was no correlation with BIO7 (BIO5 − BIO6), the bioclimatic variable we assumed to characterize best the climatic variability, but there was a weak correlation with BIO4 (temperature seasonality) (
Table 4). One reason for the ambiguity could be the small latitudinal range covered in this study and the resulting small climatic variability.
The presence of a phylogenetic signal, that is, when related species share similar thermal responses, may indicate constraints of their thermal niches, which dictate the environments in which they can persist [
73], or it may indicate similar selection pressures and similar environmental effects [
73,
74,
75]. In our assessed seed bug species, kinship seems to have played a lesser, or at least no unambiguous, role in the adaptation to high environmental temperatures, and no role in low temperature adaptation (
Table 5). This concurs with the findings of Teets and Hahn [
76] on CT
min of
Drosophila. Concerning CT
max, it concurs with findings of Ayrinhac et al. [
77] and Hoffmann et al. [
78] regarding the upper thermal limits in
Drosophila, of Terblanche et al. [
79] in
Glossina pallidipes, of Vorhees et al. [
39] in
Culex tarsalis, and of Hamblin et al. [
80] in a variety of bee species. In other words, our results suggest that despite phylogenetic relatedness, the seed bugs differ in their CT
max because of adaptation to differing thermal environments, indicated by the comprehensive significant effects of bioclimatic variables on CT
max (
Figure 2,
Table 3 and
Table 4).
In the face of global warming and the current “Mediterranaziation” of the Heteroptera fauna [
81,
82] in regions of previously unsuitable climatological (i.e., “too cold”) conditions, we suggest that seed bugs will not experience troubles to disperse towards the north. Temperature-induced longer (reproductive) seasons and shortened diapause may result in an increase of generations per year or/and increased overwintering success [
83,
84,
85]. Although winter mortality may vary depending on parameters such as gender, microhabitat choice, as well as size and coloration [
86,
87,
88], temperature seems to be the main factor. That which might hinder the true bugs’ dispersion could be the minimum environmental temperatures, which still can reach quite low values in Europe despite higher annual temperatures. Yukawa et al. [
89] suggested the northward range expansion of
Nezara viridula in Japan to be due to global warming, because the monthly mean temperature for January in the newly invaded areas exceeded the limit temperature of 5 °C, below which winter mortality increases by 15%–16.5% per 1 °C decrease in mean winter temperature [
87,
90,
91]. Concerning the ability to invade colder areas in reaction to global temperature increase, we suggest a similar dispersion capacity of European seed bugs than predicted for
Nezara viridula in Kyushu in Southern Japan [
89].

 ,
, 




{kind=link}
{kind=link}
{kind=link}
{kind=link}