1. Introduction
Ludwigia L. is a large monophyletic genus of wetland plant species within the Onagraceae family, currently classified into 23 sections with 88 taxa, including 83 species [
1,
2,
3]. Most species (80%) are native to the New World, although the genus is pantropical with some (largely naturalized) representation in temperate Europe, Africa, and Eurasia [
1]. Of particular interest is a group of
Ludwigia spp. from the largely aquatic
Ludwigia section
Jussiaea [
4], native to South America [
1,
3]. They have invaded both aquatic and riparian ecosystems in many regions worldwide [
5,
6] and are now considered among the most aggressive weeds in the world [
7]. In the USA, four
Ludwigia section
Jussiaea taxa have naturalized in aquatic systems of the South Atlantic, Gulf, and/or Pacific coastal states [
8]:
Ludwigia hexapetala (Hook. & Arn.) Zardini, Gu & P. H. Raven;
Ludwigia peploides (Kunth) P. H. Raven subsp.
peploides;
Ludwigia peploides (Kunth) P. H. Raven subsp.
montevidensis (Spreng.) P. H. Raven; and
Ludwigia grandiflora (Michx.) Greuter & Burdet. Rapid biomass production of these species impacts ecological processes in aquatic ecosystems, displacing desired native wildlife and vegetation [
5,
9,
10,
11,
12]. Characteristic dense mats over the water’s surface also impede navigation and interfere with recreational activities, irrigation, drainage, and agricultural production [
5,
8].
Management of invasive
Ludwigia spp. in the USA has to date relied on physical and chemical methods [
5]. These options only provide short-term control and require repeated annual treatments, and some can be limited by regulatory restrictions in environmentally sensitive systems [
8,
13,
14].
Ludwigia hexapetala and
L. peploides also produce viable seeds with a high capacity for germination under a wide range of temperatures [
15,
16], resulting in persistent seedbanks that require long-term management programs [
10]. Classical biological control provides a sustainable alternative to these conventional techniques as it can be used alone or as a tool in integrated management programs. Unlike other control methods, successful biological control might provide permanent, landscape-level weed suppression if exotic natural enemies are carefully selected and establish upon introduction [
17].
Recently, a biological control program targeting exotic
Ludwigia spp. was initiated by the United States Department of Agriculture’s Agricultural Research Service (USDA-ARS) at the request of water management stakeholders [
18]. However, interest in biological control of exotic
Ludwigia spp. is not new. The first foreign explorations and field host range evaluations of insect herbivores associated with
Ludwigia spp. were conducted in the 1970s in Argentina by Cordo and DeLoach [
19,
20]. More recently, an expanded comprehensive survey conducted by scientists from the Fundación para el Estudio de Especies Invasivas (FuEDEI) identified 19 insect species across 6 feeding guilds that feed on
L. hexapetala in Argentina [
21]. An insect commonly observed on
Ludwigia spp. in these surveys includes the bronze-colored, flea beetle
Lysathia flavipes (Boheman) (Coleoptera: Chrysomelidae). Adults feed and oviposit eggs singly or in masses on leaves of their host plant [
20]. All larval stages feed, develop, and pupate on leaves. Feeding by both adults and larvae can cause significant damage to host plants under high population densities [
20]. Moreover, Cordo and DeLoach [
20] conducted preliminary studies on the biology and host range of
L. flavipes and concluded that the flea beetle may be suitable for introduction into the USA, but they emphasized the need for additional research as their work focused solely on adult feeding in multiple-choice tests. This, coupled with the renewed interest in
Ludwigia biological control, led to surveys in Argentina and Uruguay during March of 2019, with a specific focus on collecting and colonizing
L. flavipes for further study.
The primary objective of this research was to investigate the host range of
L. flavipes in relationship to exotic and native
Ludwigia spp. in the USA. To accomplish this goal, both no-choice and multiple-choice host range tests were conducted with an initial suite of 11 plant species that represent the target weeds (
n = 3), as well as selecting closely related natives (
n = 7) and one exotic species. Additionally, the life-history parameters (i.e., survival, development, and fecundity) of
L. flavipes were collected to aid in interpreting herbivore performance across host plants. Herein we test the hypothesis that
L. flavipes is host specific to species within the
Jussiaea section of the genus
Ludwigia given that there are no native representatives of the
Jussiaea in the USA [
18].
2. Materials and Methods
2.1. Origin and Rearing of Lysathia flavipes
Lysathia flavipes adults were collected from L. hexapetala plants near Punta Del Diablo (34°00′47.0″ S, 53°35′53.9″ W) and Lascano (33°44′53.9″ S, 54°07′55.4″ W) in Uruguay during March 2019, and mixed across sites. The nascent L. flavipes colony was exported from Uruguay under the scientific collection permit N° 9/2019 supplied by the Dirección Nacional de Medio Ambiente (DINAMA) and imported under USDA APHIS-PPQ permit # P526P-19-03070 to a USDA-ARS containment facility in Albany, California. Species identity was confirmed by the USDA-ARS Systematic Entomology Laboratory at the Smithsonian Institution, National Museum of Natural History, Washington, DC, USA. The colony was maintained on a laboratory benchtop in the Albany containment facility under ambient temperature conditions (20–25 °C). Adults were kept in 946 mL plastic containers with a piece of fine mesh cloth integrated into the lid to allow air circulation and prevent condensation. Approximately 15 adults per container fed and reproduced on a bouquet composed of three 15 cm-long excised L. hexapetala stems inserted into a plastic floral water tube. Stems were changed weekly. In-between feedings, water was added to the floral tubes as needed to maintain plant turgor. Periodically, older bouquets harboring eggs were retained and reared to augment colony numbers. Colonies were reared exclusively on L. hexapetala, originally collected from the Sacramento–San Joaquin River Delta in northern California (38°00′08.8″ N, 121°34′06.9″ W). All subsequent biology and host range studies were conducted in an environmental chamber set to a constant 25 °C (±1 °C), with a 14:10 h (L:D) photoperiod.
2.2. Egg Development of Lysathia flavipes
Fresh bouquets of L. hexapetala stems were provided to all adult colony containers. Plant material was removed after 24 h and all eggs were collected. Eggs from different clusters were mixed and spread across 15 replicate Petri dishes (90 mm diameter). Each Petri dish contained 10 eggs that were carefully placed on sterile filter paper (Whatman No. 2) moistened with water prior to sealing the dish with Parafilm® to avoid desiccation. Replicated Petri dishes were arranged in a completely randomized design in a chamber and their position was rotated daily when egg hatching was monitored. Water was added as needed to keep the filter paper moist. The mean development time (days) and hatching rate (proportion) per Petri dish were calculated. Additionally, egg length (as proxy of larvae size) was measured from 30 randomly selected eggs that originated from different parental females using a dissecting microscope.
2.3. Preimaginal Development of Lysathia flavipes
Twenty neonate larvae (≤24 h) from the egg development study were individually transferred using a fine brush onto the young leaves of an L. hexapetala stem (10 cm) inserted into a floral water tube situated within an enclosed 237 mL plastic container. Fresh stems were provided weekly and water in the floral tube was replenished three times per week. Larvae were monitored daily and developmental stage (visualized by the presence of exuviae) was recorded until adult metamorphosis. On the day each molt occurred, head capsule size was measured on the greatest width of the head (genae) using a dissecting microscope equipped with an ocular micrometer. Subsequently, the number of instars, head capsule size of each instar (mm), and development time (days) of each stage were calculated. The total development time from egg to adult was calculated by adding the mean egg, larval, and pupal development times. Finally, the length of 40 randomly selected adults (20 females and 20 males) were measured from the most forward part of the head (at the frons between the eyes) to the last abdominal segment.
2.4. Fecundity of Lysathia flavipes
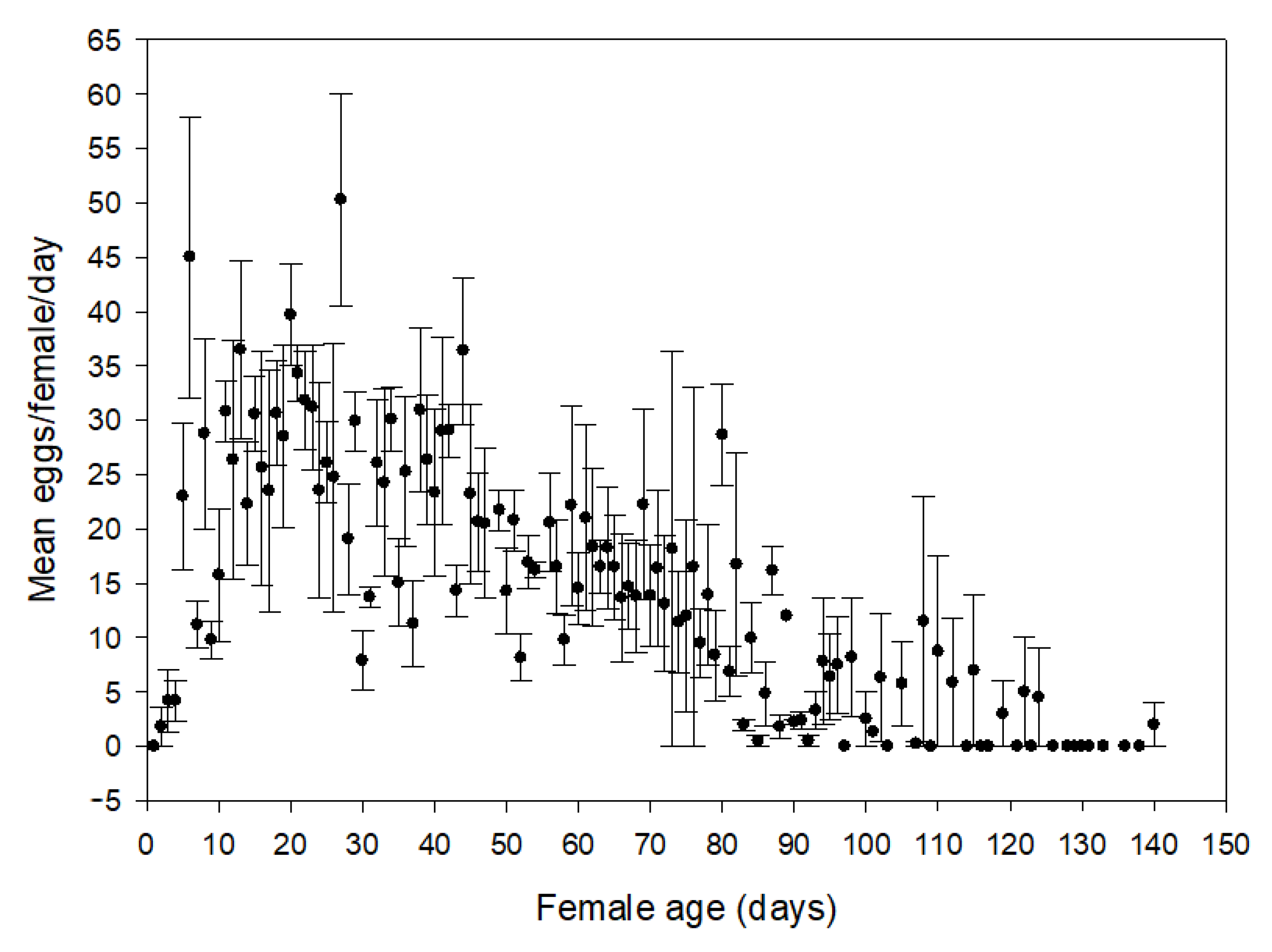
Newly molted adults from the larval development study were collected and combined in 15 replicate groups of one female and two males. Each group was enclosed in a 473 mL plastic container with a 10 cm-long L. hexapetala stem inserted into a floral water tube. The L. hexapetala stem was replaced daily and examined for eggs under a dissecting microscope. Once oviposition was confirmed, the L. hexapetala stem was replaced three times per week and eggs laid on the old stem were counted. This process was continued until the death of the female. If a male died, it was replaced with another male from the colony. Measures of pre-oviposition and oviposition periods, as well as longevity were recorded. Mean daily fecundity during the oviposition period and total fecundity were calculated for each female.
2.5. Host Range Experiments: Test Plants
A subset of the
Ludwigia taxa and close relatives were used to provide insights into the suitability of
L. flavipes as a biocontrol agent in the USA. The test plant list was comprised of eleven taxa from the Onagraceae: three exotic
Ludwigia targets (
L. hexapetala,
L. peploides subsp.
peploides, and
L. peploides subsp.
montevidensis) and seven native taxa (
Ludwigia polycarpa Short & Peter,
Ludwigia repens J. R. Forst.,
Ludwigia palustris (L.) Elliott,
Epilobium ciliatum Raf. subsp.
ciliatum,
Epilobium canum (Greene) P. H. Raven,
Clarkia amoena (Lehm.) A. Nelson & J. F. Macbr., and
Oenothera elata Kunth subsp.
hookeri (Torr. & A. Gray) W. Dietr. & W. L. Wagner). We also tested
Ludwigia decurrens Walter, an additional congener with biogeographical similarity to the target plants that is presumed native to eastern-central USA.
Ludwigia decurrens is non-native to California where it established approximately ten years ago as an invasive noxious weed in rice fields [
22]. The native test plants (non-targets) were selected based on their range in phylogenetic relationship to the three target plants [
18]. All test plants were used in both no-choice and multiple-choice host range experiments. Plants were propagated over time in a greenhouse under controlled temperature (20–32 °C), with a 14:10 h (L:D) photoperiod and ambient humidity conditions and incorporated into the host range tests as available, with
L. hexapetala replicated in each test as the control.
2.6. No-Choice Development and Oviposition Tests
Four neonate larvae (≤24 h) were randomly assigned a host plant species and transferred with a fine brush onto the young leaves of a 10 cm-long stem (experimental unit) inserted into a floral water tube. Five replicate stems were individually placed in a 473 mL plastic container (4 neonates × 5 replicate stems = 20 larvae per test plant species). Larvae were transferred to fresh stems of their assigned test plant species twice per week. Water in the floral tubes was replenished three times per week during which time the larvae were observed under a dissecting microscope to record survival and developmental stage. Larval survival rate (proportion) and mean development time (n = 4 larvae per replicate) from 1st instar to adult was calculated for each replicate stem.
The resulting adults from the no-choice development tests were collected and grouped by emergence date. Following the colony-rearing methods described above, adults were fed and kept in a rearing container for one week to allow sexual maturation and mating. Gravid females were identified by conducting 48 h oviposition tests, then each was paired with two males and placed in a 473 mL plastic container together with a bouquet (2–3 10 mL-long stems) from the test plant the female was reared on. This process was repeated until five replicate females per test plant were evaluated. If there were not enough males from the experiments described above, adult males from the colony were used. Eggs were collected from each bouquet after seven days and counted under a dissecting microscope. Eggs from each female were then placed on moistened filter paper in individual Petri dishes sealed with Parafilm®. Egg viability (hatching) was monitored three times a week until all eggs hatched or became deflated (indicating mortality). Water was added to the filter paper as needed during monitoring. Subsequently, the number of eggs oviposited and egg viability (hatching rate: eggs hatched/eggs oviposited) was calculated for each replicate female.
2.7. Multiple-Choice Oviposition Tests
Eight adult pairs were collected from the colony and placed in a plastic container (36 L × 28 W × 24 H cm) together with 3–5 bouquets (one bouquet per test plant). Because the 11 test plant species were not available simultaneously, this study was conducted in three separate trials where a different set of plant species, including L. hexapetala as the control, was tested in each trial (i.e., 5, 5, and 3 plant species per trial). Five replicate bouquets per plant species were assessed in each trial (5 replicates × 11 plant species = 55 bouquets total). Each bouquet was composed of three 15 mL-long stems from a single test plant species inserted into a floral water tube as a potential source for feeding and oviposition. The side walls of the container were modified with a piece of fine mesh cloth to allow air circulation and prevent condensation within the container. Adults were collected and returned to the colony after a five-day oviposition period and eggs oviposited on each bouquet were quantified. The presence of feeding damage was noted, but not quantified.
2.8. Data Analyses
One-way ANOVAs were used to compare body length and longevity between female and male adults, and differences in larval survival, mean larval development time, eggs per female, and egg viability (hatching rate per female) between test plant species in no-choice tests. A Linear Mixed Model was used to test the effect of plant species on oviposition (eggs per plant) with trial as a blocking factor in the multiple-choice tests. Plant species was a fixed effect factor in all models. Post-hoc pairwise comparisons between test plants were made with Tukey’s HSD (α = 0.05). Plants species on which larvae failed to survive were omitted from the analyses. Data were tested for normality using Shapiro–Wilk tests. Egg hatch rate and larval survival data were arcsine square-root transformed, and eggs per female, eggs per plant, and adult body length data were square-root transformed to normalize results prior to analyses. All analyses were conducted using SAS Software (JMP®, version 13).
4. Discussion
The life-history characteristics and host range of
L. flavipes were studied to determine if the flea beetle is a suitable biological control agent of invasive
Ludwigia spp. in the USA. Particular interest was placed on testing
L. flavipes to complete the work started by Cordo and DeLoach [
20] and because of the success of other flea beetles in weed biological control programs, including
Euphorbia esula L. (leafy spurge),
Alternanthera philoxeroides (Mart.) Griseb. (alligatorweed),
Myriophyllum aquaticum (Vell.) Verdc. (parrot feather), and
Jacobaea vulgaris Gaertn. (tansy ragwort) [
23,
24,
25,
26]. Beyond biological control, however, these data also have relevance to general biological parameters of the South American herbivore. Development times of immature stages on
L. hexapetala were similar those on
M. aquaticum, with generation time from egg to adult approximately 20 d on both host plants [
20]. The size of the
L. flavipes females (ca. 8 mm) and eggs (ca. 0.7 mm) are longer than the conspecific
L. ludoviciana, which are 4–5 mm and ca. 0.5 mm, respectively [
27]. The generation time of
L. flavipes also appears at least seven days faster than
L. ludoviciana, although direct comparisons are complicated by different temperature conditions [
27]. Possibly the greatest disparity between these two closely related species is their respective reproductive performance and longevity. The number of eggs oviposited by
L. flavipes during a female’s lifetime averaged over 1500 (longevity: 90 d), as compared to that of
L. ludoviciana which ranged from 5 to 142 (longevity: 57 d). This fast generation time and high fecundity greatly facilitated rearing and experimentation with
L. flavipes during host-specificity testing.
We found no evidence to support the hypothesis that
L. flavipes is host specific to species within the
Ludwigia section
Jussiaea, including the target weeds
L. hexapetala,
L. peploides subsp.
peploides, and
L. peploides subsp.
montevidensis. Under no-choice conditions,
L. flavipes larvae fed and completed development on the three target weed species but also successfully developed on five of the seven native plant species tested. Additionally, there was no difference in
L. flavipes survival or development time when comparing these eight host species (
Table 1). Interestingly, higher levels of variability in survivorship was observed between
Ludwigia species as compared to other confamilials. It is noteworthy that
L. flavipes was unable to develop on
L. decurrens as compared to more distantly related hosts.
While larval survival and development provides important insights into host specificity, comparing adult fitness between individuals reared on different species can reveal sublethal effects of suboptimal hosts. However, no-choice oviposition tests demonstrate that
L. flavipes females readily oviposited viable eggs on the same plant species that supported their development. In terms of number of eggs oviposited,
L. flavipes females did not distinguish between the three exotic weeds and two native plant species (
L. palustris and
E. ciliatum subsp.
ciliatum). Surprisingly, the highest number of eggs were observed on the native
L. polycarpa. Egg hatching rates did not differ between the test plant species, suggesting no apparent decrease in fitness over a generation of feeding exclusively on the test plant species. It is surmised from these data that several native plants included in this study are likely to support sustained
L. flavipes populations for more than one generation. Collectively, these data also indicate that the physiological host range of
L. flavipes does not mirror the phylogenetic relationship of the
Ludwigia species and their more distant relatives [
18].
While larvae may lack host specificity, as shown for
L. flavipes, females can restrict host use through selective oviposition. Therefore, multiple-choice tests were conducted to provide insights into the herbivore’s ovipositional host plant selection preferences. Herein, however,
L. flavipes females did not demonstrate a strong ovipositional preference for species in the
Jussiaea over more distantly related species (
Table 2). As with larval development and survival, female ovipositional patterns did not correlate with host phylogenetic relatedness, as evidenced by the highly variable results for species within the
Jussiaea as well as between confamilials. When provided a choice,
L. flavipes placed markedly more eggs on
C. amoena as compared to
L. hexapetala, the host from which the insects were originally collected in South America. Additionally,
L. flavipes also oviposited on species that do not support development. The flea beetle oviposited on
L. decurrens,
E. canum, and
O. elata subsp.
hookeri, yet larval development tests showed that the larvae cannot complete development on these species. These data provide strong evidence that
L. flavipes females oviposit broadly among hosts that range from optimal to unacceptable suitability for larval survival.
Collectively, these data indicate that
L. flavipes is not a specialist of the
Jussiaea or
Ludwigia but is a more generalist feeding herbivore. The findings are consistent with field observations of
L. flavipes by Vogt and Cordo [
28], who recorded adults feeding on
Ludwigia peploides (Kunth) P. H. Raven and
Myriophyllum brasiliense Cambess (=
M. aquaticum). Cordo and DeLoach [
20] also documented heavy adult feeding by
L. flavipes on leaf discs of
L. peploides and
M. aquaticum, as well as slight feeding on species from four other genera:
Salvinia auriculata Aubl. (salvinia),
Spirodela intermedia W.D.J. Koch (giant duckweed),
Beta vulgaris L. (leaf beet), and
Brassica oleracea L. (cabbage).
Lysathia flavipes adults have also been recorded in association with other species in the Onagraceae in Argentina:
Oenothera indecora Cambess,
Oenothera rosea L’Hér. ex Aiton, and
Oenothera glazioviana Micheli (=
O. erythrosepala Borbás) [
29]. Hernández and Cabrera Walsh [
21] observed feeding by a
Lysathia spp., which is likely to be
L. flavipes, on
L. grandiflora,
Ludwigia elegans (Cambess.) H. Hara,
Ludwigia leptocarpa (Nutt.) H. Hara, and
Ludwigia bonariensis (Micheli) H. Hara in Argentina. The data reported herein, however, are the first to quantify larval developmental parameters and oviposition behaviors of the flea beetle, which are critical for estimating the herbivore’s host range.
Although our results demonstrate that
L. flavipes is not a suitable biological control agent for invasive
Ludwigia spp. in the USA, these should not be extended to presume
L. flavipes is equally unsuitable for biological control in other parts of the world where exotic
Ludwigia spp. are also problematic (e.g., European countries). Flea beetles have been among the most effective weed biological control agents worldwide, including another
Lysathia spp. introduced into South Africa from Brazil in 1994 to control
M. aquaticum [
30].
Lysathia flavipes may still be considered for introduction elsewhere and these data can guide future host range testing as well as facilitate the rearing and handling of
Lysathia spp. in general.


 ,
, 


{kind=link}