

Habitat Suitability of Ixodes ricinus Ticks Carrying Pathogens in North-East Italy

 , ,
, ,  , , ,
, , ,  , , ,
, , ,  , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
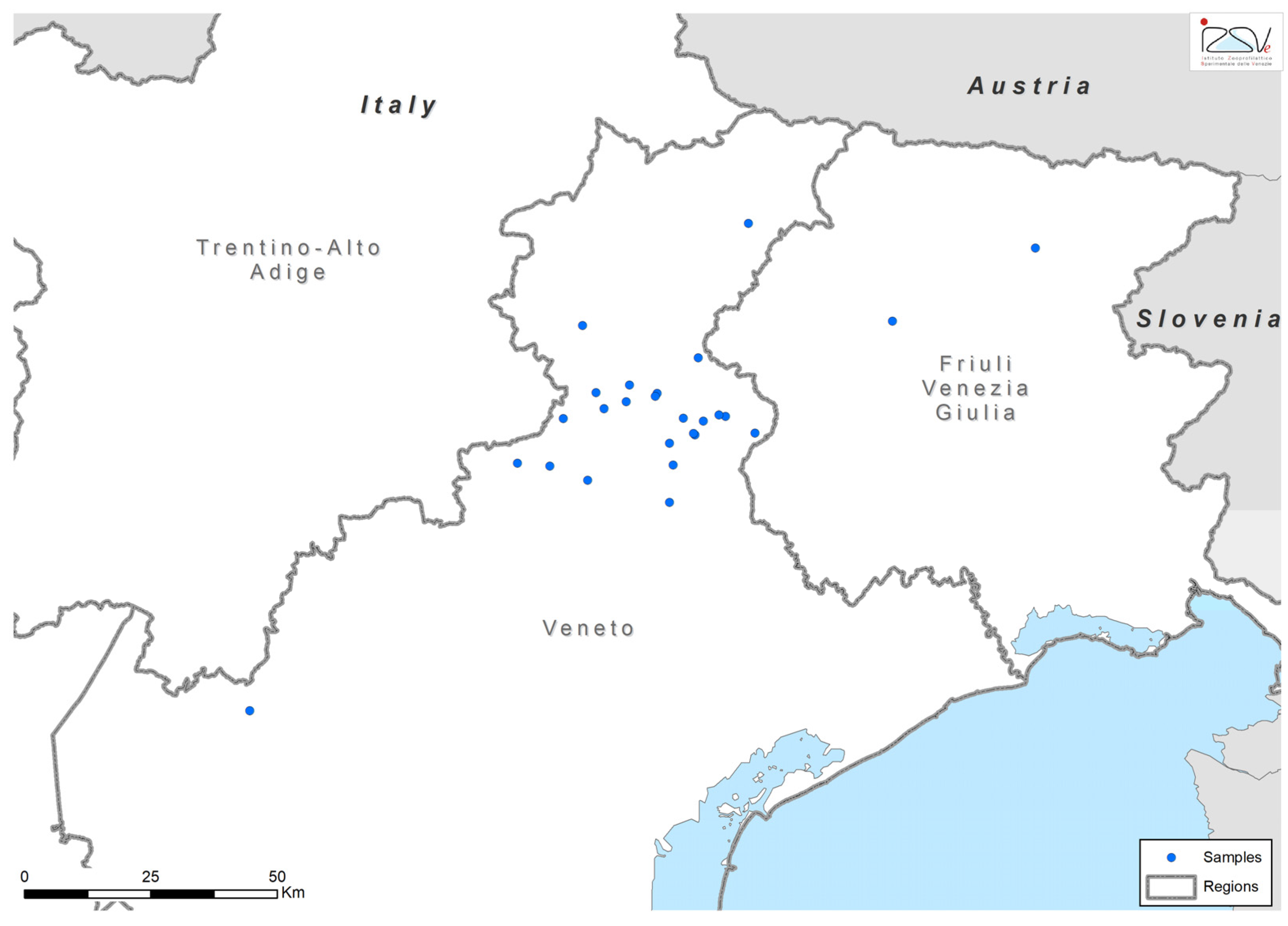
2.1. Study Design and Data Collection
2.2. Molecular Detection of Tick-Borne Pathogens
2.3. Environmental Data
2.4. Habitat Suitability Modelling
2.4.1. Probability of Pathogen Presence in Ticks
2.4.2. Model Assessment
3. Results
3.1. Tick and Tick-Borne Pathogen Detection
3.2. Model Results for Probability of Pathogen Presence
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Beltrame, A.; Laroche, M.; Degani, M.; Perandin, F.; Bisoffi, Z.; Raoult, D.; Parola, P. Tick-borne pathogens in removed ticks Veneto, northeastern Italy: A cross-sectional investigation. Travel Med. Infect. Dis. 2018, 26, 58–61. [Google Scholar] [CrossRef] [PubMed]
- Otranto, D.; Dantas-Torres, F.; Giannelli, A.; Latrofa, M.S.; Cascio, A.; Cazzin, S.; Ravagnan, S.; Montarsi, F.; Zanzani, S.A.; Manfredi, M.T.; et al. Ticks infesting humans in Italy and associated pathogens. Parasites Vectors 2014, 7, 328. [Google Scholar] [CrossRef] [PubMed]
- Bertola, M.; Montarsi, F.; Obber, F.; Da Rold, G.; Carlin, S.; Toniolo, F.; Porcellato, E.; Falcaro, C.; Mondardini, V.; Ormelli, S.; et al. Occurrence and Identification of Ixodes ricinus Borne Pathogens in Northeastern Italy. Pathogens 2021, 10, 1181. [Google Scholar] [CrossRef] [PubMed]
- Rezza, G.; Farchi, F.; Pezzotti, P.; Ruscio, M.; Presti, A.L.; Ciccozzi, M.; Mondardini, V.; Paternoster, C.; Bassetti, M.; Merelli, M.; et al. Tick-borne encephalitis in north-east Italy: A 14-year retrospective study, January 2000 to December 2013. Eurosurveillance 2015, 20, 30034. [Google Scholar] [CrossRef] [PubMed]
- Ruiz-Fons, F.; Fernández-de-Mera, I.G.; Acevedo, P.; Gortázar, C.; de la Fuente, J. Factors driving the abundance of Ixodes ricinus ticks and the prevalence of zoonotic I. ricinus-borne pathogens in natural foci. Appl. Environ. Microbiol. 2012, 78, 2669–2676. [Google Scholar] [CrossRef] [PubMed]
- Tagliapietra, V.; Rosà, R.; Arnoldi, D.; Cagnacci, F.; Capelli, G.; Montarsi, F.; Hauffe, H.C.; Rizzoli, A. Saturation deficit and deer density affect questing activity and local abundance of Ixodes ricinus (Acari, Ixodidae) in Italy. Vet. Parasitol. 2011, 183, 114–124. [Google Scholar] [CrossRef]
- Cayol, C.; Koskela, E.; Mappes, T.; Siukkola, A.; Kallio, E.R. Temporal dynamics of the tick Ixodes ricinus in northern Europe: Epidemiological implications. Parasites Vectors 2017, 10, 166. [Google Scholar] [CrossRef]
- Medlock, J.M.; Hansford, K.M.; Bormane, A.; Derdakova, M.; Estrada-Peña, A.; George, J.-C.; Golovljova, I.; Jaenson, T.G.T.; Jensen, J.-K.; Jensen, P.M.; et al. Driving forces for changes in geographical distribution of Ixodes ricinus ticks in Europe. Parasites Vectors 2013, 6, 1. [Google Scholar] [CrossRef]
- John, H.K.S.; Masuoka, P.; Jiang, J.; Takhampunya, R.; Klein, T.A.; Kim, H.C.; Chong, S.T.; Song, J.W.; Kim, Y.J.; Farris, C.M.; et al. Geographic distribution and modeling of ticks in the Republic of Korea and the application of tick models towards understanding the distribution of associated pathogenic agents. Ticks Tick Borne Dis. 2021, 12, 101686. [Google Scholar] [CrossRef]
- Namgyal, J.; Lysyk, T.J.; Couloigner, I.; Checkley, S.; Gurung, R.B.; Tenzin, T.; Dorjee, S.; Cork, S.C. Identification, Distribution, and Habitat Suitability Models of Ixodid Tick Species in Cattle in Eastern Bhutan. Trop. Med. Infect. Dis. 2021, 6, 27. [Google Scholar] [CrossRef]
- Pangrácová, L.; Derdáková, M.; Pekárik, L.; Hviščová, I.; Víchová, B.; Stanko, M.; Hlavatá, H.; Peťko, B. Ixodes ricinus abundance and its infection with the tick-borne pathogens in urban and suburban areas of Eastern Slovakia. Parasites Vectors 2013, 6, 238. [Google Scholar] [CrossRef] [PubMed]
- Gethmann, J.; Hoffmann, B.; Kasbohm, E.; Süss, J.; Habedank, B.; Conraths, F.J.; Beer, M.; Klaus, C. Research paper on abiotic factors and their influence on Ixodes ricinus activity—Observations over a two-year period at several tick collection sites in Germany. Parasitol. Res. 2020, 119, 1455–1466. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Vozmediano, A.; Krawczyk, A.I.; Sprong, H.; Rossi, L.; Ramassa, E.; Tomassone, L. Ticks climb the mountains: Ixodid tick infestation and infection by tick-borne pathogens in the Western Alps. Ticks Tick Borne Dis. 2020, 11, 101489. [Google Scholar] [CrossRef]
- Estrada-Peña, A. Distribution, abundance, and habitat preferences of Ixodes ricinus (Acari: Ixodidae) in northern Spain. J. Med. Entomol. 2001, 38, 361–370. [Google Scholar] [CrossRef] [PubMed]
- Capelli, G.; Ravagnan, S.; Montarsi, F.; Ciocchetta, S.; Cazzin, S.; Porcellato, E.; Babiker, A.M.; Cassini, R.; Salviato, A.; Cattoli, G.; et al. Occurrence and identification of risk areas of Ixodes ricinus-borne pathogens: A cost-effectiveness analysis in north-eastern Italy. Parasites Vectors 2012, 5, 61. [Google Scholar] [CrossRef]
- Da Rold, G.; Ravagnan, S.; Soppelsa, F.; Porcellato, E.; Soppelsa, M.; Obber, F.; Citterio, C.V.; Carlin, S.; Danesi, P.; Montarsi, F.; et al. Ticks are more suitable than red foxes for monitoring zoonotic tick-borne pathogens in northeastern Italy. Parasites Vectors 2018, 11, 137. [Google Scholar] [CrossRef]
- Mertens, M.; Schmidt, K.; Ozkul, A.; Groschup, M.H. The impact of Crimean-Congo hemorrhagic fever virus on public health. Antivir. Res. 2013, 98, 248–260. [Google Scholar] [CrossRef]
- Manilla, G. Acari, Ixodida (Fauna d’Italia 36); Calderini: Bologna, Italy, 1998. [Google Scholar]
- Cringoli, G.; Iori, A.; Rinaldi, L.; Veneziano, V.; Genchi, C. Mappe Parassitologiche: Zecche; Rolando Editore: Napoli, Italy, 2005. [Google Scholar]
- Schwaiger, M.; Cassinotti, P. Development of a quantitative real-time RT-PCR assay with internal control for the laboratory detection of tick borne encephalitis virus (TBEV) RNA. J. Clin. Virol. Off. Publ. Pan Am. Soc. Clin. Virol. 2003, 27, 136–145. [Google Scholar] [CrossRef]
- Bioclimatic Variables—WorldClim 1 Documentation. 2020. Available online: https://www.worldclim.org/data/bioclim.html (accessed on 13 February 2022).
- Corine Land Cover, (n.d.). Available online: https://land.copernicus.eu/en/products/corine-land-cover (accessed on 1 October 2021).
- Mazzucato, M.; Marchetti, G.; Barbujani, M.; Mulatti, P.; Fornasiero, D.; Casarotto, C.; Scholamacchia, F.; Manca, G.; Ferre, N. An integrated system for the management of environmental data to support veterinary epidemiology. Front. Vet. Sci. 2023, 21, 1069979. [Google Scholar] [CrossRef]
- Phillips, S.J.; Dudík, M.; Schapire, R.E. Maxent Software for Modelling Species Niches and Distributions (Version 3.4.1). Available online: http://biodiversityinformatics.amnh.org/open_source/maxent/ (accessed on 13 February 2022).
- Phillips, S.J. A Brief Tutorial on Maxent. 2017. Available online: http://biodiversityinformatics.amnh.org/open_source/maxent/ (accessed on 13 February 2022).
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2020; Available online: https://www.R-project.org/ (accessed on 13 February 2022).
- RStudio Team. RStudio: Integrated Development for R; RStudio Inc.: Boston, MA, USA, 2019; Available online: http://www.rstudio.com/ (accessed on 13 February 2022).
- Nasirian, H. Ticks infected with Crimean-Congo hemorrhagic fever virus (CCHFV): A decision approach systematic review and meta-analysis regarding their role as vectors. Travel Med. Infect. Dis. 2022, 47, 102309. [Google Scholar] [CrossRef]
- Vanroy, T.; Martel, A.; Baeten, L.; Fonville, M.; Lens, L.; Pasmans, F.; Sprong, H.; Strubbe, D.; Van Gestel, M.; Verheyen, K. The effect of forest structural complexity on tick-borne pathogens in questing ticks and small mammals. For. Ecol. Manag. 2024, 562, 121944. [Google Scholar] [CrossRef]
- Jacobs, D.; Fox, M.; Gibbons, L.; Hermosilla, C. Principles of Veterinary Parasitology; John Wiley & Sons, Ltd.: Chichester, UK, 2015. [Google Scholar]
- Liu, W.; Han, Y.; Li, J.; Tian, X.; Liu, Y. Factors affecting relative humidity and its relationship with the long-term variation of fog-haze events in the Yangtze River Delta. Atmos. Environ. 2018, 193, 242–250. [Google Scholar] [CrossRef]
- Kraetzig, N.M. 5 Things to Know about NDVI (Normalized Difference Vegetation Index) UP42. 2020. Available online: https://up42.com/blog/tech/5-things-to-know-about-ndvi (accessed on 23 March 2022).
- Ogden, N.; Lindsay, R. Effects of Climate and Climate Change on Vectors and Vector-Borne Diseases: Ticks Are Different. Trends Parasitol. 2016, 32, 646–656. [Google Scholar] [CrossRef] [PubMed]
- Signorini, M.; Stensgaard, A.-S.; Drigo, M.; Simonato, G.; Marcer, F.; Montarsi, F.; Martini, M.; Cassini, R. Towards improved, cost-effective surveillance of ixodes ricinus ticks and associated pathogens using species distribution modelling. Geospat. Health 2019, 14, 46–52. [Google Scholar] [CrossRef] [PubMed]
- Rizzoli, A.; Merler, S.; Furlanello, C.; Genchi, C. Geographical Information Systems and Bootstrap Aggregation (Bagging) of Tree-Based Classifiers for Lyme Disease Risk Prediction in Trentino, Italian Alps. J. Med. Entomol. 2002, 39, 485–492. [Google Scholar] [CrossRef]
- Fitzpatrick, M.; Gotelli, N.; Ellison, A. MaxEnt versus MaxLike: Empirical comparisons with ant species distributions distributions. Ecosphere 2013, 4, 1–15. [Google Scholar] [CrossRef]
- Walker, D.H. Rickettsiae. Int. Encycl. Public Health 2017, 370–377. [Google Scholar] [CrossRef]
- Petzke, M.; Schwartz, I. Borrelia burgdorferi Pathogenesis and the Immune Response. Clin. Lab. Med. 2015, 35, 745–764. [Google Scholar] [CrossRef]
- Snowden, J.; Bartman, M.; Kong, E.L.; Simonsen, K.A. Ehrlichiosis. Encycl. Neurol. Sci. 2014, 1070–1071. [Google Scholar] [CrossRef]
- Wu, J.; Zhang, X. Ecology, Epidemiology and Global Public Health Burden of Tick-Borne Diseases. In Transmission Dynamics Tick-Borne Diseases with Co-Feeding, Developmental and Behavioural Diapause; Springer International Publishing: Cham, Switzerland, 2020; pp. 1–15. [Google Scholar] [CrossRef]
- Directive 2000/54/EC of the European Parliament and of the Council of 18 September 2000 on the Protection of Workers from Risks Related to Exposure to Biological Agents at Work (Seventh Individual Directive within the Meaning of Article 16(1) of Directive, (n.d.). Available online: https://eur-lex.europa.eu/legal-content/EN/ALL/?uri=CELEX%3A32000L0054 (accessed on 31 January 2022).
- Seasonal Changes and Vector Borne Diseases-POFT IE. 2021. Available online: https://www.protectourfuturetoo.com/en-ie/seasonal-changes-and-vector-borne-diseases/ (accessed on 31 January 2022).
- Wood, S.N. Generalized Additive Models: An Introduction with R, 2nd ed.; Chapman and Hall/CRC: Boca Raton, FL, USA, 2017. [Google Scholar]
- Venables, W.N.; Ripley, B.D. Modern Applied Statistics with S, 4th ed.; Springer: New York, NY, USA, 2002. [Google Scholar]
- Wei, T.; Simko, V. R Package ‘Corrplot’: Visualization of a Correlation Matrix (Version 0.92). 2021. Available online: https://github.com/taiyun/corrplot (accessed on 13 February 2022).
- Hijmans, R.J. Raster: Geographic Data Analysis and Modeling. R Package Version 3.4-10. 2021. Available online: https://CRAN.R-project.org/package=raster (accessed on 13 February 2022).
- Wickham, H. Ggplot2: Elegant Graphics for Data Analysis, 2nd ed.; Springer: New York, NY, USA, 2016. [Google Scholar]
- Gondard, M.; Delannoy, S.; Pinarello, V.; Aprelon, R.; Devillers, E.; Galon, C.; Pradel, J.; Vayssier-Taussat, M.; Albina, E.; Moutailler, S. Upscaling the Surveillance of Tick-Borne Pathogens in the French Caribbean Islands. Pathogens 2020, 9, 176. [Google Scholar] [CrossRef]
- Loh, S.-M.; Gofton, A.W.; Lo, N.; Gillett, A.; Ryan, U.M.; Irwin, P.J.; Oskam, C.L. Novel Borrelia species detected in echidna ticks, Bothriocroton concolor, in Australia. Parasit. Vectors 2016, 9, 339. [Google Scholar] [CrossRef] [PubMed]
- Michelet, L.; Delannoy, S.; Devillers, E.; Umhang, G.; Aspan, A.; Juremalm, M.; Chirico, J.; van der Wal, F.J.; Sprong, H.; Boye Pihl, T.P.; et al. High-throughput screening of tick-borne pathogens in Europe. Front. Cell. Infect. Microbiol. 2014, 4, 103. [Google Scholar] [CrossRef] [PubMed]
- Rar, V.A.; Fomenko, N.V.; Dobrotvorsky, A.K.; Livanova, N.N.; Rudakova, S.A.; Fedorov, E.G.; Astanin, V.B.; Morozova, O.V. Tickborne pathogen detection, Western Siberia, Russia. Emerg. Infect. Dis. 2005, 11, 1708–1715. [Google Scholar] [CrossRef] [PubMed]
- Regnery, R.L.; Spruill, C.L.; Plikaytis, B.D. Genotypic identification of rickettsiae and estimation of intraspecies sequence divergence for portions of two rickettsial genes. J. Bacteriol. 1991, 173, 1576–1589. [Google Scholar] [CrossRef] [PubMed]
- Choi, Y.-J.; Jang, W.-J.; Kim, J.-H.; Ryu, J.-S.; Lee, S.-H.; Park, K.-H.; Paik, H.-S.; Koh, Y.-S.; Choi, M.-S.; Kim, I.-S. Spotted fever group and typhus group rickettsioses in humans, South Korea. Emerg. Infect. Dis. 2005, 11, 237–244. [Google Scholar] [CrossRef] [PubMed]
- Centeno-Lima, S.; Do Rosário, V.; Parreira, R.; Maia, A.J.; Freudenthal, A.M.; Nijhof, A.M.; Jongejan, F. A fatal case of human babesiosis in Portugal: Molecular and phylogenetic analysis. Trop. Med. Int. Health 2008, 8, 760–764. [Google Scholar] [CrossRef]
- Monteil, V.; Salata, C.; Appelberg, S.; Mirazimi, A. Hazara virus and Crimean-Congo Hemorrhagic Fever Virus show a different pattern of entry in fully-polarized Caco-2 cell line. PLoS Negl. Trop. Dis. 2020, 14, e0008863. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
| Spatial Predictors | Code | Source |
|---|---|---|
| Annual Mean Temperature | Bio 1 | WorldClim *2 |
| Mean Diurnal Range | Bio 2 | WorldClim |
| Isothermality | Bio 3 | WorldClim |
| Temperature Seasonality | Bio 4 | WorldClim |
| Maximum Temperature of Warmest Month | Bio 5 | WorldClim |
| Minimum Temperature of Coldest Month | Bio 6 | WorldClim |
| Temperature Annual Range | Bio 7 | WorldClim |
| Mean Temperature of Wettest Quarter | Bio 8 | WorldClim |
| Mean Temperature of Driest Quarter | Bio 9 | WorldClim |
| Mean Temperature of Warmest Quarter | Bio 10 | WorldClim |
| Mean Temperature of Coldest Quarter | Bio 11 | WorldClim |
| Annual Precipitation | Bio 12 | WorldClim |
| Precipitation of Wettest Month | Bio 13 | WorldClim |
| Precipitation of Driest Month | Bio 14 | WorldClim |
| Precipitation Seasonality | Bio 15 | WorldClim |
| Precipitation of Wettest Quarter | Bio 16 | WorldClim |
| Precipitation of Driest Quarter | Bio 17 | WorldClim |
| Precipitation of Warmest Quarter | Bio 18 | WorldClim |
| Precipitation of Coldest Quarter | Bio 19 | WorldClim |
| Digital Terrain Model | DTM | SPOT *3 |
| Normalized Difference Vegetation Index | NDVI | COPERNICUS *4 |
| Corine Land Cover, level 2 *1 | CLC | COPERNICUS |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Huitink, M.; de Rooij, M.; Montarsi, F.; Salvati, M.V.; Obber, F.; Da Rold, G.; Sgubin, S.; Mazzotta, E.; di Martino, G.; Mazzucato, M.; et al. Habitat Suitability of Ixodes ricinus Ticks Carrying Pathogens in North-East Italy. Pathogens 2024, 13, 836. https://doi.org/10.3390/pathogens13100836
Huitink M, de Rooij M, Montarsi F, Salvati MV, Obber F, Da Rold G, Sgubin S, Mazzotta E, di Martino G, Mazzucato M, et al. Habitat Suitability of Ixodes ricinus Ticks Carrying Pathogens in North-East Italy. Pathogens. 2024; 13(10):836. https://doi.org/10.3390/pathogens13100836
Chicago/Turabian StyleHuitink, Maartje, Myrna de Rooij, Fabrizio Montarsi, Maria Vittoria Salvati, Federica Obber, Graziana Da Rold, Sofia Sgubin, Elisa Mazzotta, Guido di Martino, Matteo Mazzucato, and et al. 2024. "Habitat Suitability of Ixodes ricinus Ticks Carrying Pathogens in North-East Italy" Pathogens 13, no. 10: 836. https://doi.org/10.3390/pathogens13100836