
Hydrocarbon Cycling in the Tokamachi Mud Volcano (Japan): Insights from Isotopologue and Metataxonomic Analyses

 , , , and
, , , and
Abstract
:1. Introduction
2. Materials and Methods
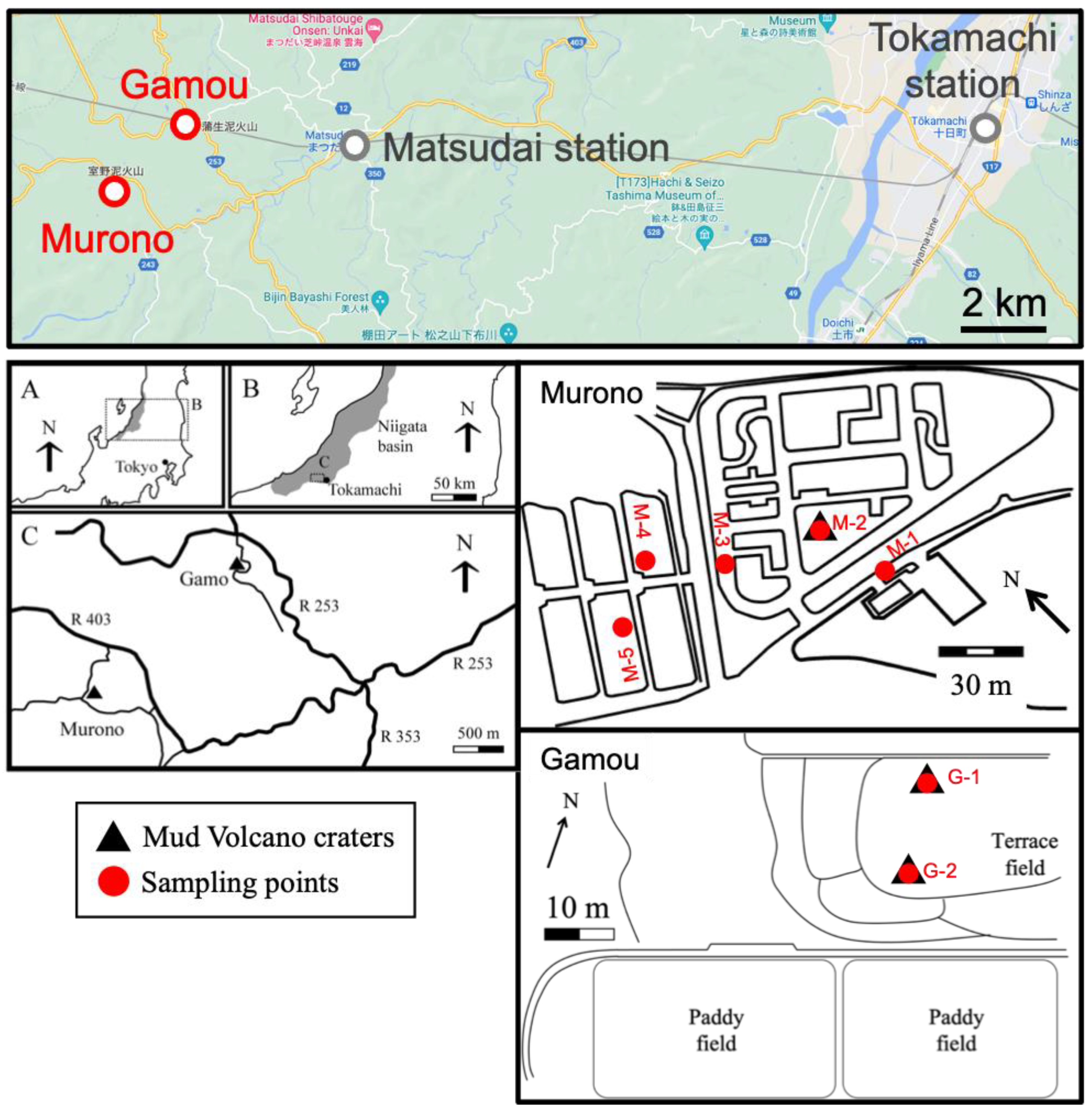
2.1. Study Site and Sampling Points
2.2. DNA Extraction and Bioinformatic Analysis
2.3. Bulk Isotope Analysis of Hydrocarbons and CO2
2.4. Position-Specific 13C Isotope Analysis of Propane
3. Results and Discussion
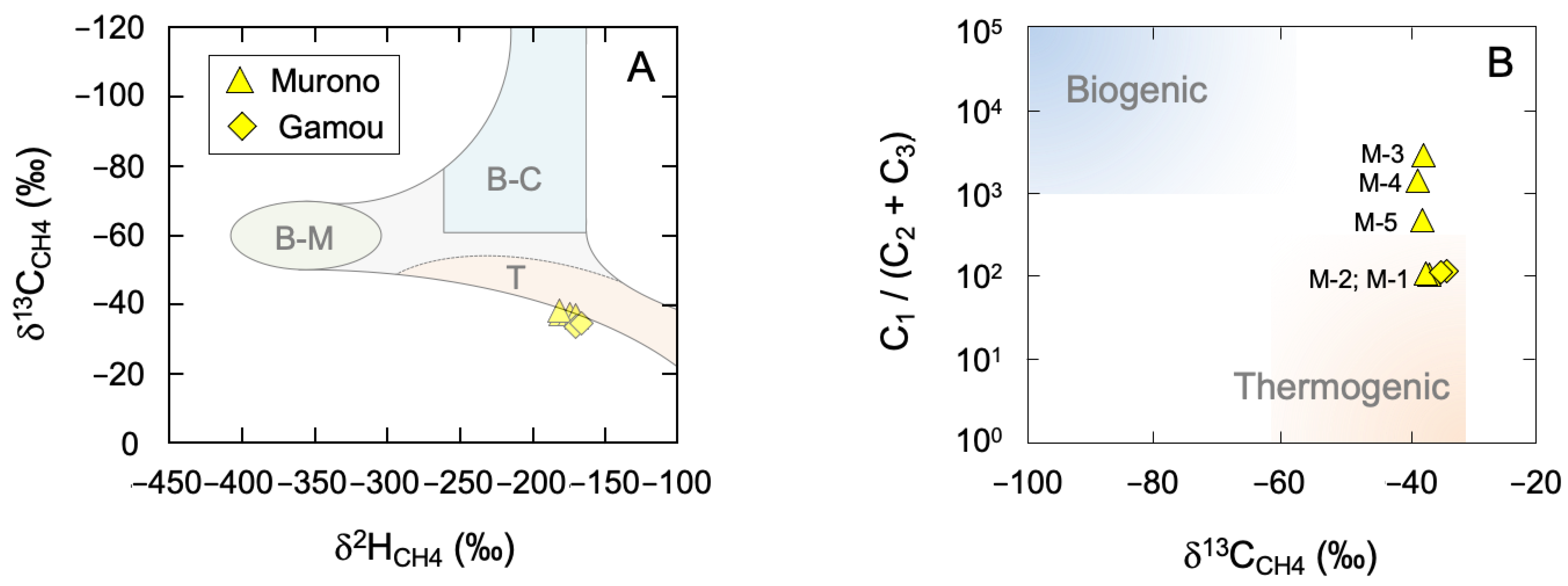
3.1. Bulk and Intramolecular Isotope and Chemical Composition
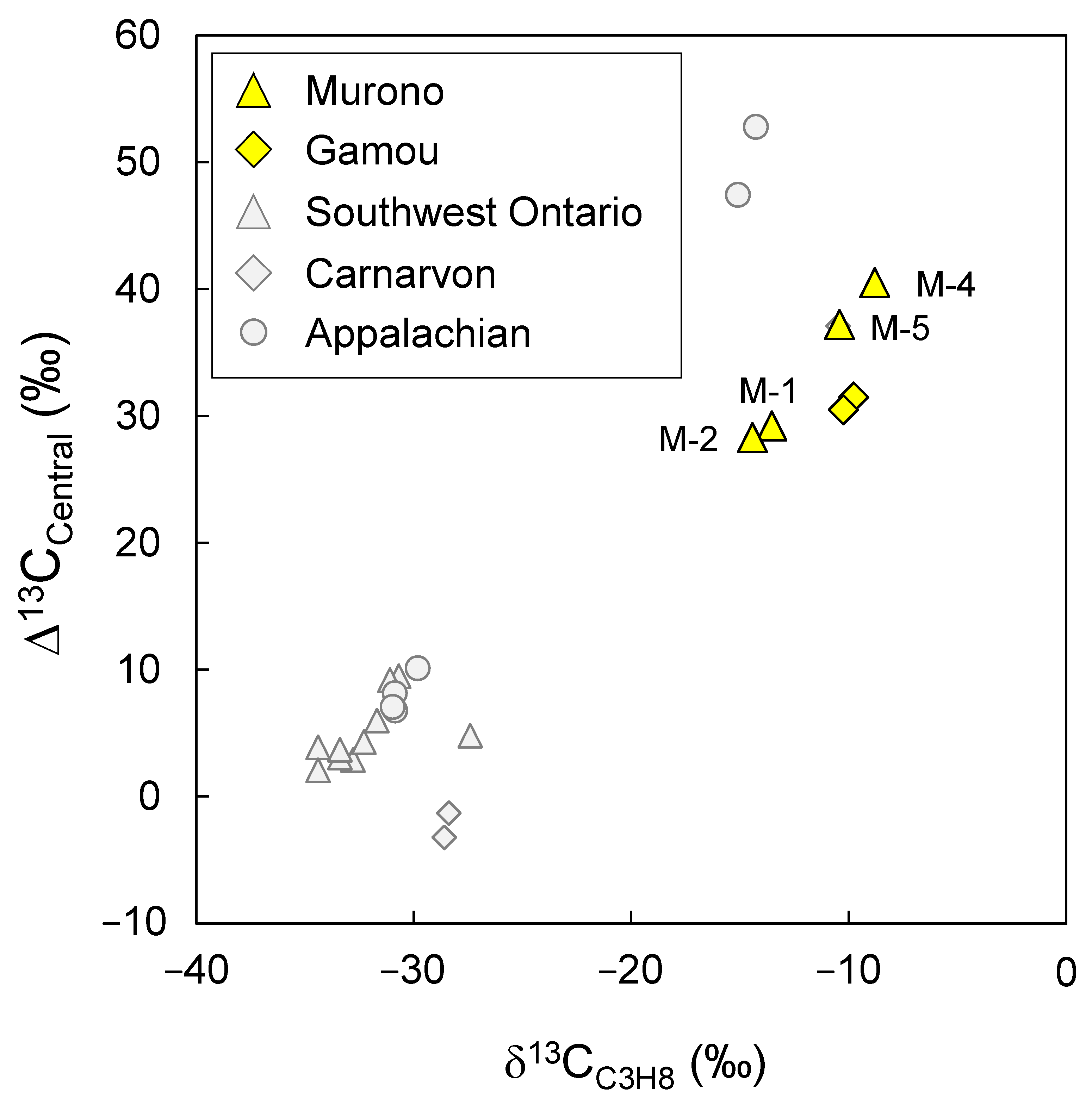
3.2. Position-Specific 13C Composition of Propane
3.3. Is Ethane Also Biodegraded?
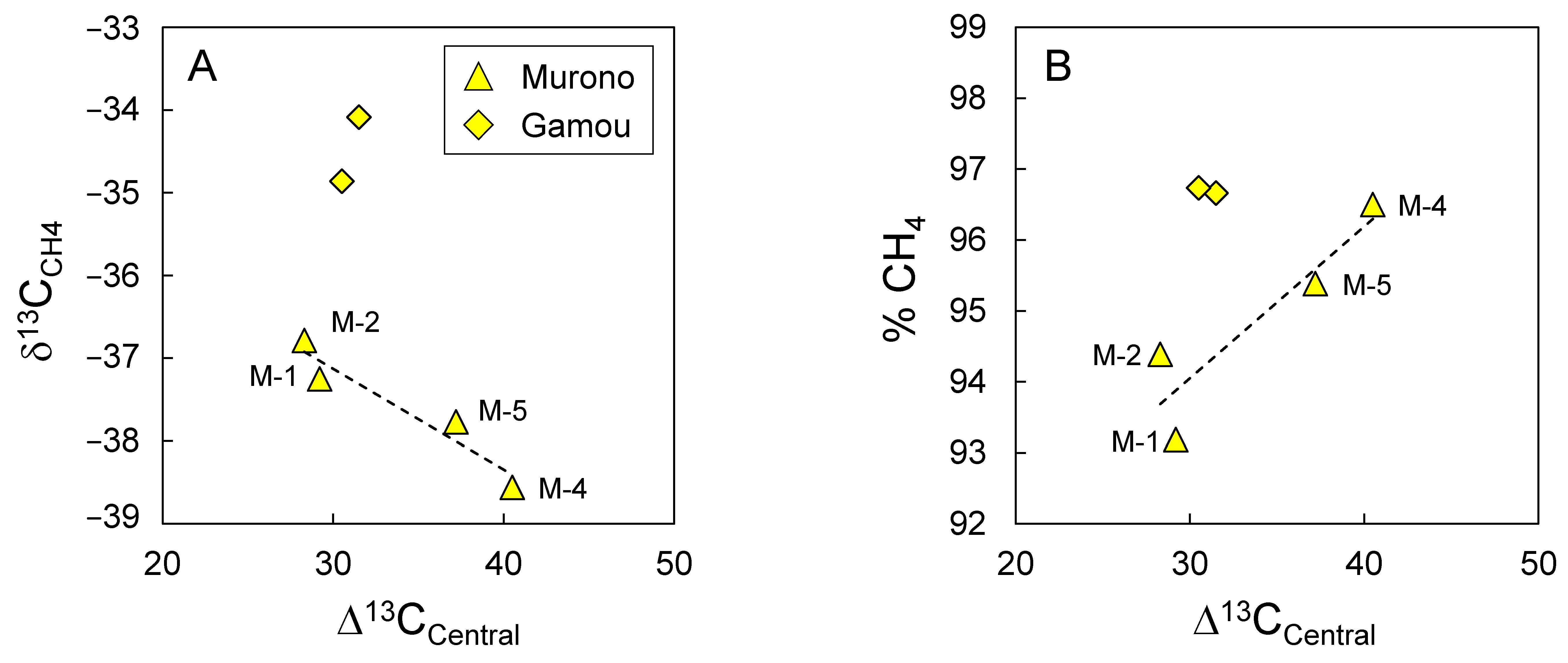
3.4. Evidence for Secondary Microbial Methane
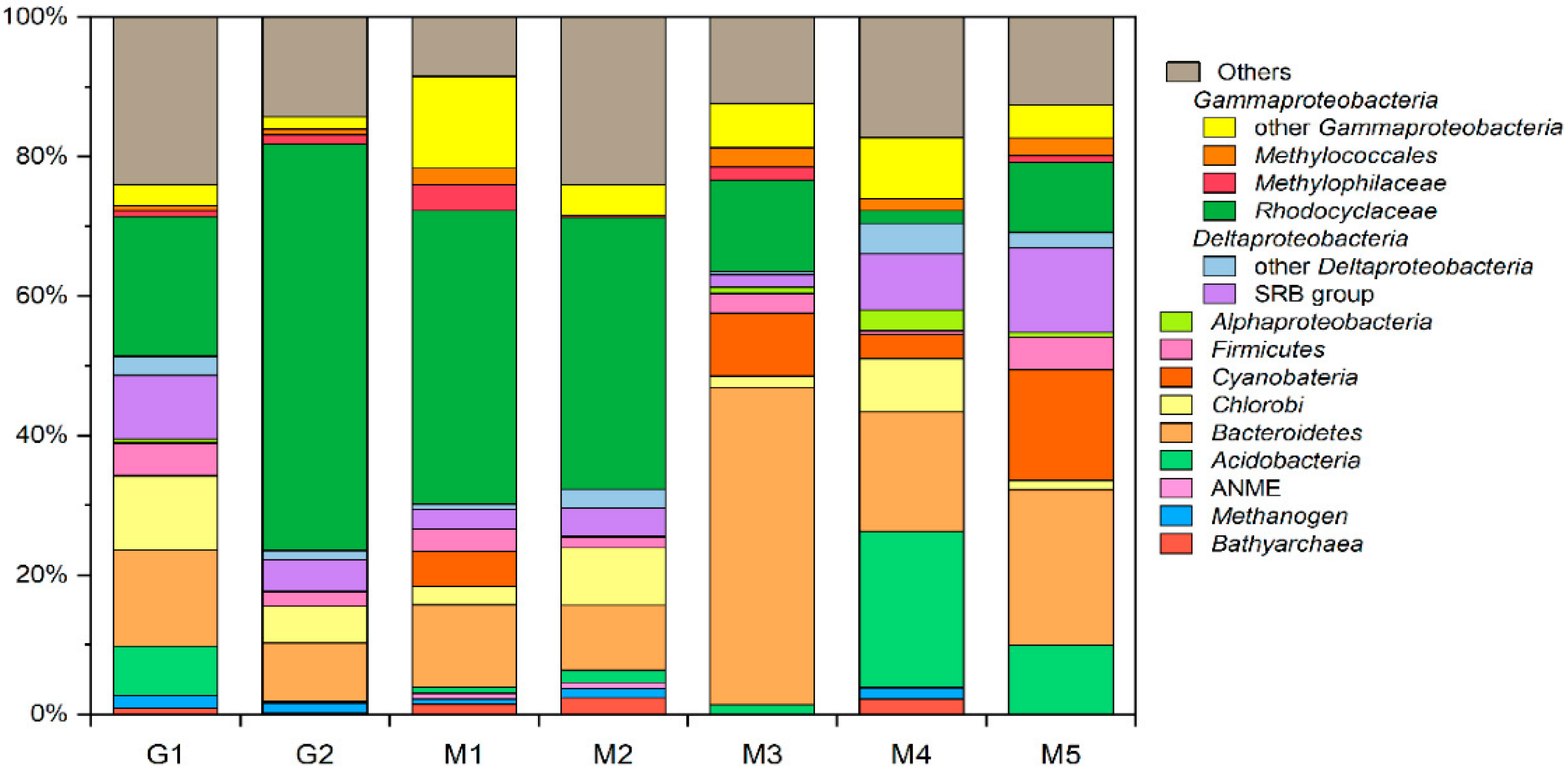
3.5. Microbial Community Composition and Members Involved in Hydrocarbon Degradation
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Mazzini, A.; Etiope, G. Mud Volcanism: An Updated Review. Earth Sci. Rev. 2017, 168, 81–112. [Google Scholar] [CrossRef] [Green Version]
- Hoshino, T.; Toki, T.; Ijiri, A.; Morono, Y.; Machiyama, H.; Ashi, J.; Okamura, K.; Inagaki, F. Atribacteria from the Subseafloor Sedimentary Biosphere Disperse to the Hydrosphere through Submarine Mud Volcanoes. Front. Microbiol. 2017, 8, 1135. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Manna, S.K.; Das, B.K.; Mohanty, B.P.; Bandopadhyay, C.; Das, N.; Baitha, R.; Das, A.K. Exploration of Heterotrophic Bacterial Diversity in Sediments of the Mud Volcano in the Andaman and Nicobar Islands, India. Environ. Nanotechnol. Monit. Manag. 2021, 16, 100465. [Google Scholar] [CrossRef]
- Mardanov, A.V.; Kadnikov, V.V.; Beletsky, A.V.; Ravin, N.V. Sulfur and Methane-Oxidizing Microbial Community in a Terrestrial Mud Volcano Revealed by Metagenomics. Microorganisms 2020, 8, 1333. [Google Scholar] [CrossRef]
- Brož, P.; Krýza, O.; Wilson, L.; Conway, S.J.; Hauber, E.; Mazzini, A.; Raack, J.; Balme, M.R.; Sylvest, M.E.; Patel, M.R. Experimental Evidence for Lava-like Mud Flows under Martian Surface Conditions. Nat. Geosci. 2020, 13, 403–407. [Google Scholar] [CrossRef]
- Komatsu, G.; Okubo, C.H.; Wray, J.J.; Ojha, L.; Cardinale, M.; Murana, A.; Orosei, R.; Chan, M.A.; Ormö, J.; Gallagher, R. Small Edifice Features in Chryse Planitia, Mars: Assessment of a Mud Volcano Hypothesis. Icarus 2016, 268, 56–75. [Google Scholar] [CrossRef]
- Pondrelli, M.; Rossi, A.P.; Ori, G.G.; van Gasselt, S.; Praeg, D.; Ceramicola, S. Mud Volcanoes in the Geologic Record of Mars: The Case of Firsoff Crater. Earth Planet. Sci. Lett. 2011, 304, 511–519. [Google Scholar] [CrossRef]
- Skinner, J.A.; Mazzini, A. Martian Mud Volcanism: Terrestrial Analogs and Implications for Formational Scenarios. Mar. Pet. Geol. 2009, 26, 1866–1878. [Google Scholar] [CrossRef]
- Milkov, A.V.; Sassen, R.; Apanasovich, T.V.; Dadashev, F.G. Global Gas Flux from Mud Volcanoes: A Significant Source of Fossil Methane in the Atmosphere and the Ocean. Geophys. Res. Lett. 2003, 30, 1037. [Google Scholar] [CrossRef] [Green Version]
- Liu, Q.; Wu, X.; Wang, X.; Jin, Z.; Zhu, D.; Meng, Q.; Huang, S.; Liu, J.; Fu, Q. Carbon and Hydrogen Isotopes of Methane, Ethane, and Propane: A Review of Genetic Identification of Natural Gas. Earth Sci. Rev. 2019, 190, 247–272. [Google Scholar] [CrossRef]
- Eiler, J.M. The Isotopic Anatomies of Molecules and Minerals. Annu. Rev. Earth Planet. Sci. 2013, 41, 411–441. [Google Scholar] [CrossRef]
- Gilbert, A. The Organic Isotopologue Frontier. Annu. Rev. Earth Planet. Sci. 2021, 49, 435–464. [Google Scholar] [CrossRef]
- Douglas, P.M.J.; Stolper, D.A.; Eiler, J.M.; Sessions, A.L.; Lawson, M.; Shuai, Y.; Bishop, A.; Podlaha, O.G.; Ferreira, A.A.; Neto, E.V.S.; et al. Methane Clumped Isotopes: Progress and Potential for a New Isotopic Tracer. Org. Geochem. 2017, 113, 262–282. [Google Scholar] [CrossRef] [Green Version]
- Stolper, D.A.; Lawson, M.; Davis, C.L.; Ferreira, A.A.; Neto, E.V.S.; Ellis, G.S.; Lewan, M.D.; Martini, A.M.; Tang, Y.; Schoell, M.; et al. Formation Temperatures of Thermogenic and Biogenic Methane. Science 2014, 344, 1500. [Google Scholar] [CrossRef]
- Wang, D.T.; Gruen, D.S.; Lollar, B.S.; Hinrichs, K.-U.; Stewart, L.C.; Holden, J.F.; Hristov, A.N.; Pohlman, J.W.; Morrill, P.L.; Könneke, M.; et al. Nonequilibrium Clumped Isotope Signals in Microbial Methane. Science 2015, 348, 428. [Google Scholar] [CrossRef] [Green Version]
- Young, E.D. A Two-Dimensional Perspective on CH4 Isotope Clumping: Distinguishing Process from Source. In Deep Carbon: Past to Present; Orcutt, B.N., Daniel, I., Dasgupta, R., Eds.; Cambridge University Press: Cambridge, UK, 2019; pp. 388–414. ISBN 978-1-108-47749-9. [Google Scholar]
- Clog, M.; Lawson, M.; Peterson, B.; Ferreira, A.A.; Neto, E.V.S.; Eiler, J.M. A Reconnaissance Study of 13C–13C Clumping in Ethane from Natural Gas. Geochim. Cosmochim. Acta 2018, 223, 229–244. [Google Scholar] [CrossRef] [Green Version]
- Taguchi, K.; Yamamoto, T.; Nakagawa, M.; Gilbert, A.; Ueno, Y. A Fluorination Method for Measuring the 13C-13C Isotopologue of C2 Molecules. Rapid Commun. Mass Spectrom. 2020, 34, e8761. [Google Scholar] [CrossRef]
- Gilbert, A.; Yamada, K.; Suda, K.; Ueno, Y.; Yoshida, N. Measurement of Position-Specific 13C Isotopic Composition of Propane at the Nanomole Level. Geochim. Cosmochim. Acta 2016, 177, 205–216. [Google Scholar] [CrossRef]
- Li, Y.; Zhang, L.; Xiong, Y.; Gao, S.; Yu, Z.; Peng, P. Determination of Position-Specific Carbon Isotope Ratios of Propane from Natural Gas. Org. Geochem. 2018, 119, 11–21. [Google Scholar] [CrossRef]
- Liu, C.; McGovern, G.P.; Liu, P.; Zhao, H.; Horita, J. Position-Specific Carbon and Hydrogen Isotopic Compositions of Propane from Natural Gases with Quantitative NMR. Chem. Geol. 2018, 491, 14–26. [Google Scholar] [CrossRef]
- Piasecki, A.; Sessions, A.; Lawson, M.; Ferreira, A.A.; Neto, E.V.S.; Eiler, J.M. Analysis of the Site-Specific Carbon Isotope Composition of Propane by Gas Source Isotope Ratio Mass Spectrometer. Geochim. Cosmochim. Acta 2016, 188, 58–72. [Google Scholar] [CrossRef] [Green Version]
- Gilbert, A.; Lollar, B.S.; Musat, F.; Giunta, T.; Chen, S.; Kajimoto, Y.; Yamada, K.; Boreham, C.J.; Yoshida, N.; Ueno, Y. Intramolecular Isotopic Evidence for Bacterial Oxidation of Propane in Subsurface Natural Gas Reservoirs. Proc. Natl. Acad. Sci. USA 2019, 116, 6653–6658. [Google Scholar] [CrossRef] [Green Version]
- Kniemeyer, O.; Musat, F.; Sievert, S.M.; Knittel, K.; Wilkes, H.; Blumenberg, M.; Michaelis, W.; Classen, A.; Bolm, C.; Joye, S.B.; et al. Anaerobic Oxidation of Short-Chain Hydrocarbons by Marine Sulphate-Reducing Bacteria. Nature 2007, 449, 898–901. [Google Scholar] [CrossRef] [PubMed]
- Etiope, G.; Nakada, R.; Tanaka, K.; Yoshida, N. Gas Seepage from Tokamachi Mud Volcanoes, Onshore Niigata Basin (Japan): Origin, Post-Genetic Alterations and CH4–CO2 Fluxes. Appl. Geochem. 2011, 26, 348–359. [Google Scholar] [CrossRef]
- Shinya, T.; Tanaka, K. Mud Volcanoes in Matsudai, Tokamachi City, Niigata Prefecture. J. Jpn. Soc. Nat. Disaster Sci. 2009, 118, 340–349, (In Japanese with English Abstract). [Google Scholar] [CrossRef] [Green Version]
- Ishihara, T.; Tanaka, K. Geological Structure and Groundwater Geochemistry around Mud Volcanoes in the Kamou Area, Tokamachi City, Niigata Prefecture. J. Geogr. 2009, 118, 350–372. [Google Scholar] [CrossRef] [Green Version]
- Kato, S.; Waseda, A.; Nishita, H.; Iwano, H. Geochemistry of Crude Oils and Gases from Mud Volcanoes and Their Vicinities in the Higashi-Kubiki Area, Niigata Prefecture. J. Geogr. Chigaku Zasshi 2009, 118, 455–471, (In Japanese with English Abstract). [Google Scholar] [CrossRef] [Green Version]
- Suda, K.; Gilbert, A.; Yamada, K.; Yoshida, N.; Ueno, Y. Compound– and Position–Specific Carbon Isotopic Signatures of Abiogenic Hydrocarbons from on–Land Serpentinite–Hosted Hakuba Happo Hot Spring in Japan. Geochim. Cosmochim. Acta 2017, 206, 201–215. [Google Scholar] [CrossRef]
- Parada, A.E.; Needham, D.M.; Fuhrman, J.A. Every Base Matters: Assessing Small Subunit RRNA Primers for Marine Microbiomes with Mock Communities, Time Series and Global Field Samples. Environ. Microbiol. 2016, 18, 1403–1414. [Google Scholar] [CrossRef] [PubMed]
- Bolyen, E.; Rideout, J.R.; Dillon, M.R.; Bokulich, N.A.; Abnet, C.C.; Al-Ghalith, G.A.; Alexander, H.; Alm, E.J.; Arumugam, M.; Asnicar, F.; et al. Reproducible, Interactive, Scalable and Extensible Microbiome Data Science Using QIIME 2. Nat. Biotechnol. 2019, 37, 852–857. [Google Scholar] [CrossRef]
- Quast, C.; Pruesse, E.; Yilmaz, P.; Gerken, J.; Schweer, T.; Yarza, P.; Peplies, J.; Glöckner, F.O. The SILVA Ribosomal RNA Gene Database Project: Improved Data Processing and Web-Based Tools. Nucleic Acids Res. 2013, 41, D590–D596. [Google Scholar] [CrossRef] [PubMed]
- Hut, G. Consultants’ Group Meeting on Stable Isotope Reference Samples for Geochemical and Hydrological Investigations; International Atomic Energy Agency (IAEA): Vienna, Austria, 1987. [Google Scholar]
- Zhang, N.; Snyder, G.T.; Lin, M.; Nakagawa, M.; Gilbert, A.; Yoshida, N.; Matsumoto, R.; Sekine, Y. Doubly Substituted Isotopologues of Methane Hydrate (13CH3D and 12CH2D2): Implications for Methane Clumped Isotope Effects, Source Apportionments and Global Hydrate Reservoirs. Geochim. Cosmochim. Acta 2021, 315, 127–151. [Google Scholar] [CrossRef]
- Piasecki, A.; Sessions, A.; Lawson, M.; Ferreira, A.A.; Neto, E.V.S.; Ellis, G.S.; Lewan, M.D.; Eiler, J.M. Position-Specific 13C Distributions within Propane from Experiments and Natural Gas Samples. Geochim. Cosmochim. Acta 2018, 220, 110–124. [Google Scholar] [CrossRef] [Green Version]
- Peterson, B.K.; Formolo, M.J.; Lawson, M. Molecular and Detailed Isotopic Structures of Petroleum: Kinetic Monte Carlo Analysis of Alkane Cracking. Geochim. Cosmochim. Acta 2018, 243, 169–185. [Google Scholar] [CrossRef]
- Laso-Pérez, R.; Wegener, G.; Knittel, K.; Widdel, F.; Harding, K.J.; Krukenberg, V.; Meier, D.V.; Richter, M.; Tegetmeyer, H.E.; Riedel, D.; et al. Thermophilic Archaea Activate Butane via Alkyl-Coenzyme M Formation. Nature 2016, 539, 396–401. [Google Scholar] [CrossRef]
- Adams, M.; Hoarfrost, A.; Bose, A.; Joye, S.; Girguis, P. Anaerobic Oxidation of Short-Chain Alkanes in Hydrothermal Sediments: Potential Influences on Sulfur Cycling and Microbial Diversity. Front. Microbiol. 2013, 4, 110. [Google Scholar] [CrossRef] [Green Version]
- Chen, S.-C.; Musat, N.; Lechtenfeld, O.J.; Paschke, H.; Schmidt, M.; Said, N.; Popp, D.; Calabrese, F.; Stryhanyuk, H.; Jaekel, U.; et al. Anaerobic Oxidation of Ethane by Archaea from a Marine Hydrocarbon Seep. Nature 2019, 568, 108–111. [Google Scholar] [CrossRef]
- Milkov, A.V. Worldwide Distribution and Significance of Secondary Microbial Methane Formed during Petroleum Biodegradation in Conventional Reservoirs. Org. Geochem. 2011, 42, 184–207. [Google Scholar] [CrossRef]
- Chen, S.-C.; Ji, J.; Popp, D.; Jaekel, U.; Richnow, H.-H.; Sievert, S.M.; Musat, F. Genome and Proteome Analyses Show the Gaseous Alkane Degrader Desulfosarcina Sp. Strain BuS5 as an Extreme Metabolic Specialist. Environ. Microbiol. 2022, 24, 1964–1976. [Google Scholar] [CrossRef]
- Ehrenreich, P.; Behrends, A.; Harder, J.; Widdel, F. Anaerobic Oxidation of Alkanes by Newly Isolated Denitrifying Bacteria. Arch. Microbiol. 2000, 173, 58–64. [Google Scholar] [CrossRef]
- Lovley, D.R. Dissimilatory Fe(III) and Mn(IV) Reduction. Microbiol. Rev. 1991, 55, 259–287. [Google Scholar] [CrossRef] [PubMed]
- Musat, F.; Wilkes, H.; Behrends, A.; Woebken, D.; Widdel, F. Microbial Nitrate-Dependent Cyclohexane Degradation Coupled with Anaerobic Ammonium Oxidation. ISME J. 2010, 4, 1290–1301. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Anderson, R.T.; Lovley, D.R. Hexadecane Decay by Methanogenesis. Nature 2000, 404, 722–723. [Google Scholar] [CrossRef]
- Embree, M.; Nagarajan, H.; Movahedi, N.; Chitsaz, H.; Zengler, K. Single-Cell Genome and Metatranscriptome Sequencing Reveal Metabolic Interactions of an Alkane-Degrading Methanogenic Community. ISME J. 2014, 8, 757–767. [Google Scholar] [CrossRef] [PubMed]
- Taguchi, K.; Gilbert, A.; Ueno, Y. Standardization for 13C-13C Clumped Isotope Analysis by the Fluorination Method. Rapid Commun. Mass Spectrom. 2021, 35, e9109. [Google Scholar] [CrossRef]
- Giunta, T.; Young, E.D.; Warr, O.; Kohl, I.; Ash, J.L.; Martini, A.; Mundle, S.O.C.; Rumble, D.; Pérez-Rodríguez, I.; Wasley, M.; et al. Methane Sources and Sinks in Continental Sedimentary Systems: New Insights from Paired Clumped Isotopologues 13CH3D and 12CH2D2. Geochim. Cosmochim. Acta 2019, 245, 327–351. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample | CH4 | CO2 | C2H6 | C3H8 | i-C4H10 | n-C4H10 |
|---|---|---|---|---|---|---|
| G-1 | 96.66 | 2.18 | 0.74 | 0.10 | 0.26 | 0.05 |
| G-2 | 96.74 | 2.07 | 0.76 | 0.11 | 0.28 | 0.05 |
| M-1 | 93.19 | 5.95 | 0.67 | 0.20 | bdl | bdl |
| M-2 | 94.39 | 4.70 | 0.59 | 0.32 | bdl | bdl |
| M-3 | 93.94 | 6.02 | 0.03 | 0.002 | bdl | bdl |
| M-4 | 96.50 | 3.43 | 0.05 | 0.02 | bdl | bdl |
| M-5 | 95.38 | 4.41 | 0.13 | 0.07 | bdl | bdl |
| Sample | δ13CCH4 | δDCH4 | δ13CCO2 | δ13CC2H6 | δ13CC3H8 | δ13Ci-C4H10 | δ13Cn-C4H10 | C3H8 | ||
|---|---|---|---|---|---|---|---|---|---|---|
| ∆13CCentral | δ13CCentral | δ13CTerminal | ||||||||
| G-1 | −34.1 | −170 | 28.5 | −21.2 | −9.8 | −24.3 | −14.6 | 31.5 | 11.2 | −20.3 |
| G-2 | −34.9 | −167 | 24.4 | −21.6 | −10.2 | −25.2 | −15.3 | 30.5 | 10.1 | −20.4 |
| M-1 | −37.2 | −170 | 34.5 | −26.0 | −13.5 | bdl | bdl | 29.2 | 5.9 | −23.3 |
| M-2 | −36.8 | −170 | 34.1 | −26.3 | −14.4 | bdl | bdl | 28.3 | 4.4 | −23.9 |
| M-3 | −37.0 | −181 | 32.7 | −23.3 | −1.7 | bdl | bdl | bdl | - | - |
| M-4 | −38.6 | −182 | 19.1 | −24.9 | −8.8 | bdl | bdl | 40.5 | 18.2 | −22.3 |
| M-5 | −37.8 | −175 | 33.8 | −25.1 | −10.4 | bdl | bdl | 37.2 | 14.4 | −22.8 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gilbert, A.; Nakagawa, M.; Taguchi, K.; Zhang, N.; Nishida, A.; Yoshida, N. Hydrocarbon Cycling in the Tokamachi Mud Volcano (Japan): Insights from Isotopologue and Metataxonomic Analyses. Microorganisms 2022, 10, 1417. https://doi.org/10.3390/microorganisms10071417
Gilbert A, Nakagawa M, Taguchi K, Zhang N, Nishida A, Yoshida N. Hydrocarbon Cycling in the Tokamachi Mud Volcano (Japan): Insights from Isotopologue and Metataxonomic Analyses. Microorganisms. 2022; 10(7):1417. https://doi.org/10.3390/microorganisms10071417
Chicago/Turabian StyleGilbert, Alexis, Mayuko Nakagawa, Koudai Taguchi, Naizhong Zhang, Akifumi Nishida, and Naohiro Yoshida. 2022. "Hydrocarbon Cycling in the Tokamachi Mud Volcano (Japan): Insights from Isotopologue and Metataxonomic Analyses" Microorganisms 10, no. 7: 1417. https://doi.org/10.3390/microorganisms10071417