
Acinetobacter baumannii from Samples of Commercially Reared Turkeys: Genomic Relationships, Antimicrobial and Biocide Susceptibility


 , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Sample Collection and Isolation
2.2. Antimicrobial Susceptibility Testing
2.3. Biocide Susceptibility Testing
2.4. Macrorestricton Analysis with Subsequent Pulsed-Field Gel Electrophoresis
2.5. Whole-Genome Sequencing
3. Results
3.1. Isolation
3.2. Antimicrobial Susceptibility Testing
3.3. Biocide Susceptibility Testing
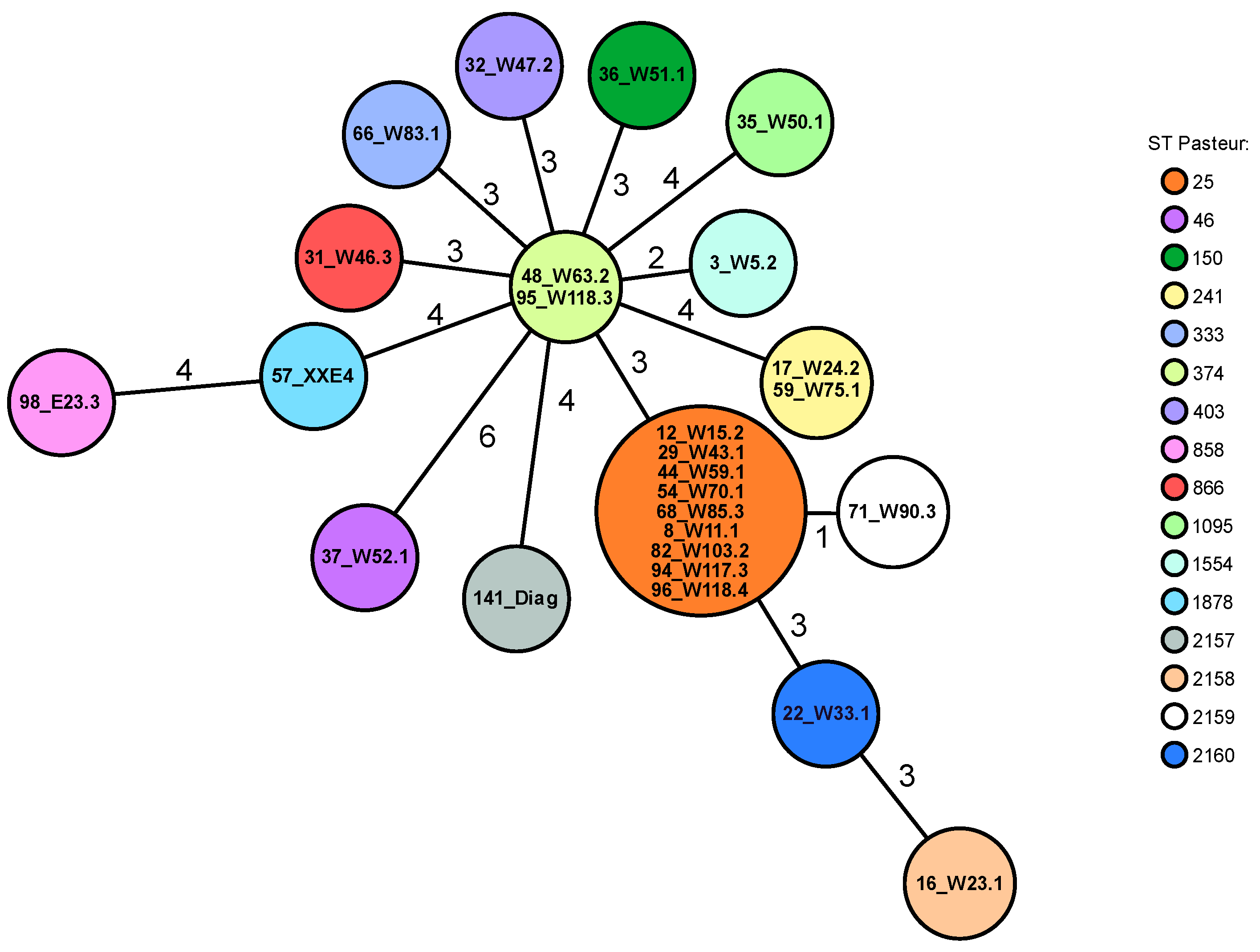
3.4. Macrorestricton Analysis with Subsequent Pulsed-Field Gel Electrophoresis
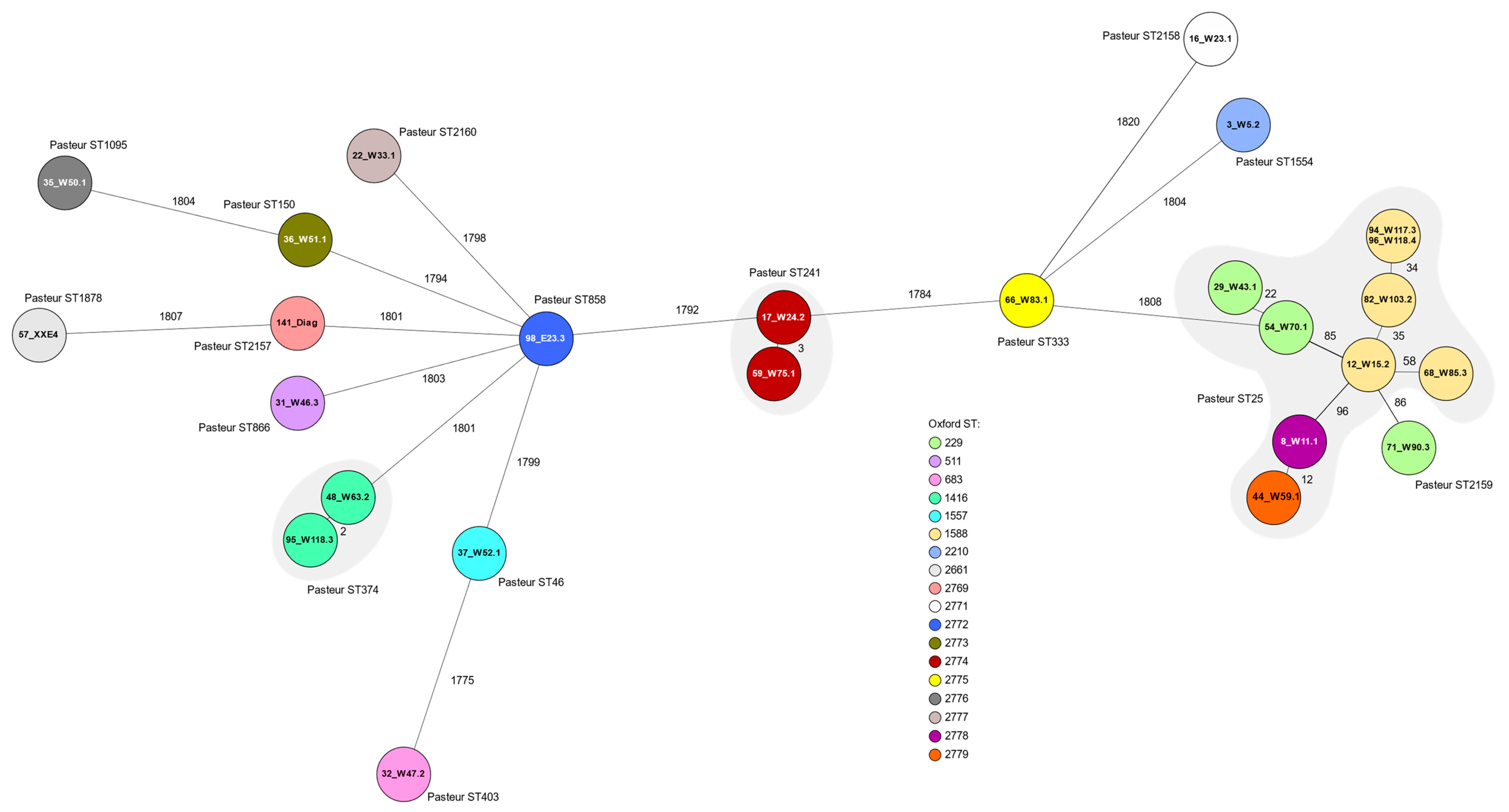
3.5. Whole-Genome Sequencing
4. Discussion
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Juni, E. Interspecies transformation of Acinetobacter: Genetic evidence for a ubiquitous genus. J. Bacteriol. 1972, 112, 917–931. [Google Scholar] [CrossRef] [Green Version]
- Higgins, P.G.; Dammhayn, C.; Hackel, M.; Seifert, H. Global spread of carbapenem-resistant Acinetobacter baumannii. J. Antimicrob. Chemother. 2010, 65, 233–238. [Google Scholar] [CrossRef] [Green Version]
- Peleg, A.Y.; Seifert, H.; Paterson, D.L. Acinetobacter baumannii: Emergence of a successful pathogen. Clin. Microbiol. Rev. 2008, 21, 538–582. [Google Scholar] [CrossRef] [Green Version]
- Turton, J.F.; Kaufmann, M.E.; Glover, J.; Coelho, J.M.; Warner, M.; Pike, R.; Pitt, T.L. Detection and typing of integrons in epidemic strains of Acinetobacter baumannii found in the United Kingdom. J. Clin. Microbiol. 2005, 43, 3074–3082. [Google Scholar] [CrossRef] [Green Version]
- Sharifipour, E.; Shams, S.; Esmkhani, M.; Khodadadi, J.; Fotouhi-Ardakani, R.; Koohpaei, A.; Doosti, Z.; Golzari, S.E. Evaluation of bacterial co-infections of the respiratory tract in COVID-19 patients admitted to ICU. BMC Infect. Dis. 2020, 20, 646. [Google Scholar] [CrossRef]
- Shinohara, D.R.; dos Santos Saalfeld, S.M.; Martinez, H.V.; Altafini, D.D.; Costa, B.B.; Fedrigo, N.H.; Tognim, M.C.B. Outbreak of endemic carbapenem-resistant Acinetobacter baumannii in a coronavirus disease 2019 (COVID-19)—Specific intensive care unit. Infect. Control Hosp. Epidemiol. 2021, 43, 815–817. [Google Scholar] [CrossRef]
- Karageorgopoulos, D.E.; Falagas, M.E. Current control and treatment of multidrug-resistant Acinetobacter baumannii infections. Lancet Infect. Dis. 2008, 8, 751–762. [Google Scholar] [CrossRef] [PubMed]
- Howard, A.; O’Donoghue, M.; Feeney, A.; Sleator, R.D. Acinetobacter baumannii: An emerging opportunistic pathogen. Virulence 2012, 3, 243–250. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vrancianu, C.O.; Gheorghe, I.; Czobor, I.B.; Chifiriuc, M.C. Antibiotic resistance profiles, molecular mechanisms and innovative treatment strategies of Acinetobacter baumannii. Microorganisms 2020, 8, 935. [Google Scholar] [CrossRef] [PubMed]
- Clinical and Laboratory Standards Institute. Performance Standards for Antimicrobial Susceptibility Testing, 32nd ed.; CLSI supplement M100: Wayne, PA, USA, 2022. [Google Scholar]
- D’Onofrio, V.; Conzemius, R.; Varda-Brkić, D.; Bogdan, M.; Grisold, A.; Gyssens, I.C.; Bedenić, B.; Barišić, I. Epidemiology of colistin-resistant, carbapenemase-producing Enterobacteriaceae and Acinetobacter baumannii in Croatia. Infect. Genet. Evol. 2020, 81, 104263. [Google Scholar] [CrossRef] [PubMed]
- Dahdouh, E.; Gómez-Gil, R.; Pacho, S.; Mingorance, J.; Daoud, Z.; Suárez, M. Clonality, virulence determinants, and profiles of resistance of clinical Acinetobacter baumannii isolates obtained from a Spanish hospital. PLoS ONE 2017, 12, e0176824. [Google Scholar] [CrossRef] [PubMed]
- Pendleton, J.N.; Gorman, S.P.; Gilmore, B.F. Clinical relevance of the ESKAPE pathogens. Expert Rev. Anti-Infect. Ther. 2013, 11, 297–308. [Google Scholar] [CrossRef]
- Rice, L.B. Federal funding for the study of antimicrobial resistance in nosocomial pathogens: No ESKAPE. J. Infect. Dis. 2008, 197, 1079–1081. [Google Scholar] [CrossRef] [PubMed]
- Wareth, G.; Neubauer, H.; Sprague, L.D. Acinetobacter baumannii—A neglected pathogen in veterinary and environmental health in Germany. Vet. Res. Commun. 2019, 43, 1–6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Klotz, P.; Higgins, P.G.; Schaubmar, A.R.; Failing, K.; Leidner, U.; Seifert, H.; Scheufen, S.; Semmler, T.; Ewers, C. Seasonal occurrence and carbapenem susceptibility of bovine Acinetobacter baumannii in Germany. Front. Microbiol. 2019, 10, 272. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, D.; Liu, Z.S.; Hu, P.; Hui, Q.; Fu, B.Q.; Lu, S.Y.; Li, Y.S.; Zou, D.Y.; Li, Z.H.; Yan, D.M.; et al. Characterization of a highly virulent and antimicrobial-resistant Acinetobacter baumannii strain isolated from diseased chicks in China. Microbiol. Immunol. 2016, 60, 533–539. [Google Scholar] [CrossRef]
- Wilharm, G.; Skiebe, E.; Higgins, P.G.; Poppel, M.T.; Blaschke, U.; Leser, S.; Heider, C.; Heindorf, M.; Brauner, P.; Jäckel, U.; et al. Relatedness of wildlife and livestock avian isolates of the nosocomial pathogen Acinetobacter baumannii to lineages spread in hospitals worldwide. Environ. Microbiol. 2017, 19, 4349–4364. [Google Scholar] [CrossRef] [Green Version]
- Rafei, R.; Hamze, M.; Pailhoriès, H.; Eveillard, M.; Marsollier, L.; Joly-Guillou, M.L.; Dabboussi, F.; Kempf, M. Extrahuman epidemiology of Acinetobacter baumannii in Lebanon. Appl. Environ. Microbiol. 2015, 81, 2359–2367. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Muller, M.G.; George, A.R.; Walochnik, J. Acinetobacter baumannii in localised cutaneous mycobacteriosis in falcons. Vet. Med. Int. 2010, 2010, 321797. [Google Scholar] [CrossRef] [Green Version]
- Zordan, S.; Prenger-Berninghoff, E.; Weiss, R.; van der Reijden, T.; van den Broek, P.; Baljer, G.; Dijkshoorn, L. Multidrug-resistant Acinetobacter baumannii in veterinary clinics, Germany. Emerg. Infect. Dis. 2011, 17, 1751–1754. [Google Scholar] [CrossRef]
- Ahmed, A.M.; Motoi, Y.; Sato, M.; Maruyama, A.; Watanabe, H.; Fukumoto, Y.; Shimamoto, T. Zoo animals as reservoirs of gram-negative bacteria harboring integrons and antimicrobial resistance genes. Appl. Environ. Microbiol. 2007, 73, 6686–6690. [Google Scholar] [CrossRef] [Green Version]
- Al Bayssari, C.; Dabboussi, F.; Hamze, M.; Rolain, J.M. Emergence of carbapenemase-producing Pseudomonas aeruginosa and Acinetobacter baumannii in livestock animals in Lebanon. J. Antimicrob. Chemother. 2015, 70, 950–951. [Google Scholar] [CrossRef] [Green Version]
- Łopińska, A.; Indykiewicz, P.; Skiebe, E.; Pfeifer, Y.; Trček, J.; Jerzak, L.; Minias, P.; Nowakowski, J.; Ledwoń, M.; Betleja, J.; et al. Low occurrence of Acinetobacter baumannii in gulls and songbirds. Pol. J. Microbiol. 2020, 69, 85–90. [Google Scholar] [CrossRef] [Green Version]
- Savin, M.; Parcina, M.; Schmoger, S.; Kreyenschmidt, J.; Käsbohrer, A.; Hammerl, J.A. Draft Genome sequences of Acinetobacter baumannii isolates recovered from sewage water from a poultry slaughterhouse in Germany. Microbiol. Resour. Announc. 2019, 8, e00553-19. [Google Scholar] [CrossRef] [Green Version]
- Martin, E.; Ernst, S.; Lotz, G.; Linsel, G.; Jäckel, U. Microbial exposure and respiratory dysfunction in poultry hatchery workers. Environ. Sci. Process. Impacts 2013, 15, 478–484. [Google Scholar] [CrossRef] [PubMed]
- Martin, E.; Jäckel, U. Characterization of bacterial contaminants in the air of a duck hatchery by cultivation based and molecular methods. J. Environ. Monit. 2011, 13, 464–470. [Google Scholar] [CrossRef] [PubMed]
- Carvalheira, A.; Casquete, R.; Silva, J.; Teixeira, P. Prevalence and antimicrobial susceptibility of Acinetobacter spp. isolated from meat. Int. J. Food Microbiol. 2017, 243, 58–63. [Google Scholar] [CrossRef] [PubMed]
- Lupo, A.; Vogt, D.; Seiffert, S.N.; Endimiani, A.; Perreten, V. Antibiotic resistance and phylogenetic characterization of Acinetobacter baumannii strains isolated from commercial raw meat in Switzerland. J. Food Prot. 2014, 77, 1976–1981. [Google Scholar] [CrossRef] [Green Version]
- Bitar, I.; Medvecky, M.; Gelbicova, T.; Jakubu, V.; Hrabak, J.; Zemlickova, H.; Karpiskova, R.; Dolejska, M. Complete nucleotide sequences of mcr-4.3-carrying plasmids in Acinetobacter baumannii sequence type 345 of human and food origin from the Czech Republic, the first case in Europe. Antimicrob. Agents Chemother. 2019, 63, e01166-19. [Google Scholar] [CrossRef]
- Tavakol, M.; Momtaz, H.; Mohajeri, P.; Shokoohizadeh, L.; Tajbakhsh, E. Genotyping and distribution of putative virulence factors and antibiotic resistance genes of Acinetobacter baumannii strains isolated from raw meat. Antimicrob. Resist. Infect. Control 2018, 7, 120. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Askari, N.; Momtaz, H.; Tajbakhsh, E. Prevalence and phenotypic pattern of antibiotic resistance of Acinetobacter baumannii isolated from different types of raw meat samples in Isfahan, Iran. Vet. Med. Sci. 2020, 6, 147–153. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schug, A.R.; Bartel, A.; Scholtzek, A.D.; Meurer, M.; Brombach, J.; Hensel, V.; Fanning, S.; Schwarz, S.; Feßler, A.T. Biocide susceptibility testing of bacteria: Development of a broth microdilution method. Vet. Microbiol. 2020, 248, 108791. [Google Scholar] [CrossRef] [PubMed]
- Schug, A.R.; Scholtzek, A.D.; Turnidge, J.; Meurer, M.; Schwarz, S.; Feßler, A.T.; The Biocide Susceptibility Study Group. Development of quality control ranges for biocide susceptibility testing. Pathogens 2022, 11, 223. [Google Scholar] [CrossRef] [PubMed]
- Seifert, H.; Dolzani, L.; Bressan, R.; van der Reijden, T.; van Strijen, B.; Stefanik, D.; Heersma, H.; Dijkshoorn, L. Standardization and interlaboratory reproducibility assessment of pulsed-field gel electrophoresis-generated fingerprints of Acinetobacter baumannii. J. Clin. Microbiol. 2005, 43, 4328–4335. [Google Scholar] [CrossRef] [Green Version]
- Hamouda, A.; Evans, B.A.; Towner, K.J.; Amyes, S.G. Characterization of epidemiologically unrelated Acinetobacter baumannii isolates from four continents by use of multilocus sequence typing, pulsed-field gel electrophoresis, and sequence-based typing of blaOXA-51-like genes. J. Clin. Microbiol. 2010, 48, 2476–2483. [Google Scholar] [CrossRef] [Green Version]
- Andrews, S. FastQC: A Quality Control Analysis Tool for High Throughput Sequencing Data. 2019. Available online: https://www.bioinformatics.babraham.ac.uk/projects/fastqc/ (accessed on 21 February 2023).
- Wick, R.R.; Judd, L.M.; Gorrie, C.L.; Holt, K.E. Unicycler: Resolving bacterial genome assemblies from short and long sequencing reads. PLoS Comput. Biol. 2017, 13, e1005595. [Google Scholar] [CrossRef] [Green Version]
- Seemann, T. Abricate, Github. Available online: https://github.com/tseeman/abricate/ (accessed on 26 August 2022).
- Feldgarden, M.; Brover, V.; Haft, D.H.; Prasad, A.B.; Slotta, D.J.; Tolstoy, I.; Tyson, G.H.; Zhao, S.; Hsu, C.-H.; McDermott, P.F.; et al. Validating the AMRFinder tool and resistance gene database by using antimicrobial resistance genotype-phenotype correlations in a collection of isolates. Antimicrob. Agents Chemother. 2019, 63, e00483-19. [Google Scholar] [CrossRef] [Green Version]
- Bortolaia, V.; Kaas, R.S.; Ruppe, E.; Roberts, M.C.; Schwarz, S.; Cattoir, V.; Philippon, A.; Allesoe, R.L.; Rebelo, A.R.; Florensa, A.F.; et al. ResFinder 4.0 for predictions of phenotypes from genotypes. J. Antimicrob. Chemother. 2020, 75, 3491–3500. [Google Scholar] [CrossRef]
- Alcock, B.P.; Raphenya, A.R.; Lau, T.T.Y.; Tsang, K.K.; Bouchard, M.; Edalatmand, A.; Huynh, W.; Nguyen, A.V.; Cheng, A.A.; Liu, S.; et al. CARD 2020: Antibiotic resistome surveillance with the comprehensive antibiotic resistance database. Nucleic Acids Res. 2020, 48, D517–D525. [Google Scholar] [CrossRef]
- Jolley, K.A.; Bray, J.E.; Maiden, M.C.J. Open-access bacterial population genomics: BIGSdb software, the PubMLST.org website and their applications. Wellcome Open Res. 2018, 3, 124. [Google Scholar] [CrossRef]
- Jolley, K.A.; Bliss, C.M.; Bennett, J.S.; Bratcher, H.B.; Brehony, C.; Colles, F.M.; Wimalarathna, H.; Harrison, O.B.; Sheppard, S.K.; Cody, A.J.; et al. Ribosomal multilocus sequence typing: Universal characterization of bacteria from domain to strain. Microbiology 2012, 158, 1005–1015. [Google Scholar] [CrossRef]
- Diancourt, L.; Passet, V.; Nemec, A.; Dijkshoorn, L.; Brisse, S. The population structure of Acinetobacter baumannii: Expanding multiresistant clones from an ancestral susceptible genetic pool. PLoS ONE 2010, 5, e10034. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bartual, S.G.; Seifert, H.; Hippler, C.; Luzon, M.A.; Wisplinghoff, H.; Rodríguez-Valera, F. Development of a multilocus sequence typing scheme for characterization of clinical isolates of Acinetobacter baumannii. J. Clin. Microbiol. 2005, 43, 4382–4390. [Google Scholar] [CrossRef] [Green Version]
- Wisplinghoff, H.; Hippler, C.; Bartual, S.G.; Haefs, C.; Stefanik, D.; Higgins, P.G.; Seifert, H. Molecular epidemiology of clinical Acinetobacter baumannii and Acinetobacter genomic species 13TU isolates using a multilocus sequencing typing scheme. Clin. Microbiol. Infect. 2008, 14, 708–715. [Google Scholar] [CrossRef] [Green Version]
- Jünemann, S.; Sedlazeck, F.J.; Prior, K.; Albersmeier, A.; John, U.; Kalinowski, J.; Mellmann, A.; Goesmann, A.; von Haeseler, A.; Stoye, J.; et al. Updating benchtop sequencing performance comparison. Nat. Biotechnol. 2013, 31, 294–296. [Google Scholar] [CrossRef] [Green Version]
- Naas, T.; Oueslati, S.; Bonnin, R.A.; Dabos, M.L.; Zavala, A.; Dortet, L.; Retailleau, P.; Iorga, B.I. Beta-lactamase database (BLDB) —Structure and function. J. Enzym. Inhib. Med. Chem. 2017, 32, 917–919. [Google Scholar] [CrossRef] [Green Version]
- Corvec, S.; Caroff, N.; Espaze, E.; Giraudeau, C.; Drugeon, H.; Reynaud, A. AmpC cephalosporinase hyperproduction in Acinetobacter baumannii clinical strains. J. Antimicrob. Chemother. 2003, 52, 629–635. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Segal, H.; Nelson, E.C.; Elisha, B.G. Genetic environment and transcription of ampC in an Acinetobacter baumannii clinical isolate. Antimicrob. Agents Chemother. 2004, 48, 612–614. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Turton, J.F.; Ward, M.E.; Woodford, N.; Kaufmann, M.E.; Pike, R.; Livermore, D.M.; Pitt, T.L. The role of ISAba1 in expression of OXA carbapenemase genes in Acinetobacter baumannii. FEMS Microbiol. Lett. 2006, 258, 72–77. [Google Scholar] [CrossRef] [Green Version]
- Figueiredo, S.; Poirel, L.; Papa, A.; Koulourida, V.; Nordmann, P. Overexpression of the naturally occurring blaOXA-51 gene in Acinetobacter baumannii mediated by novel insertion sequence ISAba9. Antimicrob. Agents Chemother. 2009, 53, 4045–4047. [Google Scholar] [CrossRef] [Green Version]
- Fallschissel, K.; Klug, K.; Kämpfer, P.; Jäckel, U. Detection of airborne bacteria in a German turkey house by cultivation-based and molecular methods. Ann. Occup. Hyg. 2010, 54, 934–943. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weernink, A.; Severin, W.P.; Tjernberg, I.; Dijkshoorn, L. Pillows, an unexpected source of Acinetobacter. J. Hosp. Infect. 1995, 29, 189–199. [Google Scholar] [CrossRef]
- Doughari, H.J.; Ndakidemi, P.A.; Human, I.S.; Benade, S. The ecology, biology and pathogenesis of Acinetobacter spp.: An overview. Microbes Environ. 2011, 26, 101–112. [Google Scholar] [CrossRef] [Green Version]
- Yusuf, I.; Skiebe, E.; Wilharm, G. Evaluation of CHROMagar Acinetobacter and MacConkey media for the recovery of Acinetobacter baumannii from soil samples. Lett. Appl. Microbiol. 2022, 76, ovac051. [Google Scholar] [CrossRef]
- Dekic, S.; Hrenovic, J.; Durn, G.; Venter, C. Survival of extensively- and pandrug-resistant isolates of Acinetobacter baumannii in soils. Appl. Soil Ecol. 2020, 147, 103396. [Google Scholar] [CrossRef]
- Mohammed, M.A.; Salim, M.T.A.; Anwer, B.E.; Aboshanab, K.M.; Aboulwafa, M.M. Impact of target site mutations and plasmid associated resistance genes acquisition on resistance of Acinetobacter baumannii to fluoroquinolones. Sci. Rep. 2021, 11, 20136. [Google Scholar] [CrossRef]
- Agersø, Y.; Guardabassi, L. Identification of Tet 39, a novel class of tetracycline resistance determinant in Acinetobacter spp. of environmental and clinical origin. J. Antimicrob. Chemother. 2005, 55, 566–569. [Google Scholar] [CrossRef]
- Agersø, Y.; Petersen, A. The tetracycline resistance determinant Tet 39 and the sulphonamide resistance gene sulII are common among resistant Acinetobacter spp. isolated from integrated fish farms in Thailand. J. Antimicrob. Chemother. 2007, 59, 23–27. [Google Scholar] [CrossRef]
- Rådström, P.; Swedberg, G.; Sköld, O. Genetic analyses of sulfonamide resistance and its dissemination in gram-negative bacteria illustrate new aspects of R plasmid evolution. Antimicrob. Agents Chemother. 1991, 35, 1840–1848. [Google Scholar] [CrossRef] [Green Version]
- Wareth, G.; Brandt, C.; Sprague, L.D.; Neubauer, H.; Pletz, M.W. WGS based analysis of acquired antimicrobial resistance in human and non-human Acinetobacter baumannii isolates from a German perspective. BMC Microbiol. 2021, 21, 210. [Google Scholar] [CrossRef]
- Wareth, G.; Linde, J.; Nguyen, N.H.; Nguyen, T.N.M.; Sprague, L.D.; Pletz, M.W.; Neubauer, H. WGS-Based Analysis of carbapenem-resistant Acinetobacter baumannii in Vietnam and molecular characterization of antimicrobial determinants and MLST in Southeast Asia. Antibiotics 2021, 10, 563. [Google Scholar] [CrossRef] [PubMed]
- Nagano, N.; Nagano, Y.; Cordevant, C.; Shibata, N.; Arakawa, Y. Nosocomial transmission of CTX-M-2 beta-lactamase-producing Acinetobacter baumannii in a neurosurgery ward. J. Clin. Microbiol. 2004, 42, 3978–3984. [Google Scholar] [CrossRef] [Green Version]
- Celenza, G.; Pellegrini, C.; Caccamo, M.; Segatore, B.; Amicosante, G.; Perilli, M. Spread of blaCTX-M-type and blaPER-2 beta-lactamase genes in clinical isolates from Bolivian hospitals. J. Antimicrob. Chemother. 2006, 57, 975–978. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pandey, D.; Singhal, N.; Kumar, M. Investigating the OXA Variants of ESKAPE Pathogens. Antibiotics 2021, 10, 1539. [Google Scholar] [CrossRef]
- Sahl, J.W.; Del Franco, M.; Pournaras, S.; Colman, R.E.; Karah, N.; Dijkshoorn, L.; Zarrilli, R. Phylogenetic and genomic diversity in isolates from the globally distributed Acinetobacter baumannii ST25 lineage. Sci. Rep. 2015, 5, 15188. [Google Scholar] [CrossRef] [Green Version]
- Tomaschek, F.; Higgins, P.G.; Stefanik, D.; Wisplinghoff, H.; Seifert, H. Head-to-head comparison of two multi-locus sequence typing (MLST) schemes for characterization of Acinetobacter baumannii outbreak and sporadic isolates. PLoS ONE 2016, 11, e0153014. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Feßler, A.T.; Scholtzek, A.D.; Schug, A.R.; Kohn, B.; Weingart, C.; Hanke, D.; Schink, A.K.; Bethe, A.; Lübke-Becker, A.; Schwarz, S. Antimicrobial and biocide resistance among canine and feline Enterococcus faecalis, Enterococcus faecium, Escherichia coli, Pseudomonas aeruginosa, and Acinetobacter baumannii isolates from diagnostic submissions. Antibiotics 2022, 11, 152. [Google Scholar] [CrossRef]
- Adams, M.D.; Pasteran, F.; Traglia, G.M.; Martinez, J.; Huang, F.; Liu, C.; Fernandez, J.S.; Lopez, C.; Gonzalez, L.J.; Albornoz, E.J.A.A.; et al. Distinct mechanisms of dissemination of NDM-1 metallo-β-lactamase in Acinetobacter species in Argentina. Antimicrob. Agents Chemother. 2020, 64, e00324-20. [Google Scholar] [CrossRef]
- Montaña, S.; Vilacoba, E.; Fernandez, J.S.; Traglia, G.M.; Sucari, A.; Pennini, M.; Iriarte, A.; Centron, D.; Melano, R.G.; Ramírez, M.S. Genomic analysis of two Acinetobacter baumannii strains belonging to two different sequence types (ST172 and ST25). J. Glob. Antimicrob. Resist. 2020, 23, 154–161. [Google Scholar] [CrossRef]
- Sennati, S.; Villagran, A.L.; Bartoloni, A.; Rossolini, G.M.; Pallecchi, L. OXA-23-producing ST25 Acinetobacter baumannii: First report in Bolivia. J. Glob. Antimicrob. Resist. 2016, 4, 70–71. [Google Scholar] [CrossRef] [Green Version]
- Higgins, P.G.; Prior, K.; Harmsen, D.; Seifert, H. Development and evaluation of a core genome multilocus typing scheme for whole-genome sequence-based typing of Acinetobacter baumannii. PLoS ONE 2017, 12, e0179228. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cerezales, M.; Xanthopoulou, K.; Wille, J.; Bustamante, Z.; Seifert, H.; Gallego, L.; Higgins, P.G. Acinetobacter baumannii analysis by core genome multi-locus sequence typing in two hospitals in Bolivia: Endemicity of international clone 7 isolates (CC25). Int. J. Antimicrob. Agents 2019, 53, 844–849. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Qiao, F.; Yu, R.; Gao, Y.; Zong, Z. Clonal diversity of Acinetobacter baumannii clinical isolates revealed by a snapshot study. BMC Microbiol. 2013, 13, 234. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fedrigo, N.H.; Xavier, D.E.; Cerdeira, L.; Fuga, B.; Marini, P.V.B.; Shinohara, D.R.; Carrara-Marroni, F.E.; Lincopan, N.; Tognim, M.C.B. Genomic insights of Acinetobacter baumannii ST374 reveal wide and increasing resistome and virulome. Infect. Genet. Evol. 2022, 97, 105148. [Google Scholar] [CrossRef]
- Das, S.R.; Haque, M.A.; Akbor, M.A.; Abdullah-Al-Mamun, M.; Debnath, G.C.; Hossain, M.S.; Hasan, Z.; Rahman, A.; Islam, M.A.; Hossain, M.A.; et al. Organophosphorus insecticides mineralizing endophytic and rhizo-spheric soil bacterial consortium influence eggplant growth-promotion. Arch. Microbiol. 2022, 204, 199. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
| Samples | No. | No. Positive | Detection Rate (%) | No. of Isolates | |
|---|---|---|---|---|---|
| chick-box-papers (meconium samples) | 118 | 94 | 79.7 | 96 * | |
| boot swab samples during rearing | 50 | 0 | 0 | 0 | |
| boot swab samples before slaughter | 82 | 2 | 2.4 | 2 | |
| lung-heart swabs (diagnostics) | 217 | 1 | 0.5 | 1 | |
| liver (diagnostics) | 88 # | 0 | 0 | 0 | |
| yolk sac (diagnostics) | 88 # | 0 | 0 | 0 | |
| total | 643 | 97 | 15.1 | 99 |
| Antimicrobial Agent | No. of Isolates for Which the MIC (mg/L) Is a: | MIC50 (mg/L) | MIC90 (mg/L) | |||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 0.008 | 0.015 | 0.03 | 0.06 | 0.12 | 0.25 | 0.5 | 1 | 2 | 4 | 8 | 16 | 32 | 64 | 128 | 256 | 512 | 1024 | |||
| Colistin | 17 | 76 | 6 | 1 | 1 | |||||||||||||||
| Streptomycin | 12 | 22 | 20 | 23 | 18 | 4 | 16 | 64 | ||||||||||||
| Neomycin | 1 | 10 | 53 | 33 | 2 | 1 | 2 | |||||||||||||
| Trimethoprim/ Sulfamethoxazole (1:19) b | 1 | 37 | 44 | 15 | 1 | 1 | 0.25 | 0.5 | ||||||||||||
| Gentamicin | 1 | 6 | 49 | 40 | 3 | 0.5 | 1 | |||||||||||||
| Nalidixic Acid | 1 | 19 | 38 | 22 | 1 | 1 | 17 | 4 | ≥256 | |||||||||||
| Ciprofloxacin | 3 | 19 | 33 | 24 | 3 | 1 | 4 | 12 | 0.25 | ≥32 | ||||||||||
| Enrofloxacin | 4 | 13 | 50 | 12 | 3 | 3 | 6 | 7 | 1 | 0.06 | 4 | |||||||||
| Marbofloxacin | 2 | 41 | 35 | 3 | 1 | 2 | 10 | 4 | 1 | 0.12 | 4 | |||||||||
| Tetracycline | 3 | 27 | 51 | 14 | 2 | 1 | 1 | 2 | 4 | |||||||||||
| Doxycycline | 12 | 45 | 32 | 8 | 1 | 1 | 0.12 | 0.5 | ||||||||||||
| Florfenicol | 1 | 20 | 61 | 17 | 128 | 256 | ||||||||||||||
| Imipenem | 37 | 61 | 1 | 0.25 | 0.25 | |||||||||||||||
| Ceftiofur | 1 | 3 | 66 | 28 | 1 | 16 | 32 | |||||||||||||
| Cefquinome | 1 | 11 | 22 | 44 | 14 | 1 | 4 | 2 | 4 | 8 | ||||||||||
| Cefotaxime | 1 | 4 | 26 | 45 | 20 | 3 | 16 | 32 | ||||||||||||
| Cefoperazone | 1 | 31 | 67 | ≥64 | ≥64 | |||||||||||||||
| Tiamulin | 99 | ≥128 | ≥128 | |||||||||||||||||
| Biocide Agent | No. of Isolates for Which the MIC (%) Is: | ||||||
|---|---|---|---|---|---|---|---|
| 0.000125 | 0.00025 | 0.0005 | 0.001 | 0.002 | 0.004 | 0.008 | |
| Benzalkonium chloride | - | - | 29 | 51 | 19 | - | - |
| Octenidine | 10 | 50 | 34 | 4 | 1 | - | - |
| Chlorhexidine | 10 | 8 | 9 | 24 | 34 | 13 | 1 |
| Polyhexanide | 3 | 12 | 32 | 25 | 19 | 5 | 3 |
| ID | PFGE | Pasteur ST 1 | Oxford ST 1 | Resistance Phenotype 2 | blaOXA3 | blaADC3 | ant(3″)-IIa | aph(3″)-Ib aph(6)-Id | sul2 | tet(39) | GyrA | ParC | Accession Number |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 68_W85.3 | A1 | 25 | 1588 | NAL, CIP | 64 | 26 | x 4 | Ser81Leu | Ser84Leu | JAPQZB010000000 | |||
| 54_W70.1 | A2 | 25 | 229 | NAL, CIP | 64 | 26 (99.7%) | x | Ser81Leu | Ser84Leu | JAPQYW010000000 | |||
| 29_W43.1 | A3 | 25 | 229 | NAL, CIP | 64 | 26 (99.7%) | x | Ser81Leu | Ser84Leu | JAPQYV010000000 | |||
| 17_W24.2 | B1 | 241 | 2774 | FOT | 91 | 52 | x | JAPQYR010000000 | |||||
| 82_W103.2 | C | 25 | 1588 | NAL, CIP, (TET) | 64 | 26 | x | Ser81Leu | Ser84Leu | JAPQZA010000000 | |||
| 94_W117.3 | D2 | 25 | 1588 | NAL, CIP | 64 | 26 | x | Ser81Leu | Ser84Leu | JAPQYZ010000000 | |||
| 3_W5.2 | E | 1554 | 2210 | 424 | 158/274 (99.2%) | x | JAPQYH010000000 | ||||||
| 48_W63.2 | F | 374 | 1416 | 259 | 26 | x | JAPQYL010000000 | ||||||
| 71_W90.3 | G | 2159 | 229 | NAL | 64 | 26 | x | Ser81Leu | JAPQZC010000000 | ||||
| 44_W59.1 | H | 25 | 2779 | NAL, CIP, TET, DOX | 64 | 26 | x | x | Ser81Leu | Ser84Leu | JAPQYU010000000 | ||
| 8_W11.1 | H | 25 | 2778 | NAL, CIP, TET, (DOX) | 64 | 26 | x | x | Ser81Leu | Ser84Leu | JAPQYT010000000 | ||
| 59_W75.1 | I | 241 | 2774 | 91 | 52 | x | JAPQYQ010000000 | ||||||
| 12_W15.2 | J | 25 | 1588 | NAL, CIP | 64 | 26 | x | Ser81Leu | Ser84Phe | JAPQYY010000000 | |||
| 96_W118.4 | K1 | 25 | 1588 | NAL, CIP | 64 | 26 | x | Ser81Leu | Ser84Leu | JAPQYX010000000 | |||
| 35_W50.1 | L | 1095 | 2776 | SXT | 208 | 249 | x | x | x | JAPQYM010000000 | |||
| 66_W83.1 | M1 | 333 | 2775 | NAL, CIP | 111 | 179 | x | Ser81Leu | Ser84Leu | JAPQYO010000000 | |||
| 22_W33.1 | M2 | 2160 | 2777 | 69 (99.8%) | 159 (99.5%) | x | JAPQYG010000000 | ||||||
| 32_W47.2 | N | 403 | 683 | 263 | 163 (99.2%) | x | JAPQYJ010000000 | ||||||
| 141_Diag * | O | 2157 | 2769 | 51 | 165 | x | JAPQZE010000000 | ||||||
| 57_XXE4 | O | 1878 | 2661 | 863 | 192 (99.7%) | x | JAPQYF010000000 | ||||||
| 16_W23.1 | P1 | 2158 | 2771 | 68 | 76 | x | JAPQYS010000000 | ||||||
| 36_W51.1 | Q2 | 150 | 2773 | SXT | 121 | 163 | x | JAPQYN010000000 | |||||
| 98_E23.3 | R | 858 | 2772 | 51 | 192 (99.5%) | x | JAPQZD010000000 | ||||||
| 37_W52.1 | S1 | 46 | 1557 | (TET) | 104 | 26 | x | JAPQYP010000000 | |||||
| 31_W46.3 | T | 866 | 511 | 385 | 158 (99.7%) | x | JAPQYI010000000 | ||||||
| 95_W118.3 | U1 | 374 | 1416 | 259 | 26 | x | JAPQYK010000000 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Schmitz, A.; Hanke, D.; Lüschow, D.; Schwarz, S.; Higgins, P.G.; Feßler, A.T. Acinetobacter baumannii from Samples of Commercially Reared Turkeys: Genomic Relationships, Antimicrobial and Biocide Susceptibility. Microorganisms 2023, 11, 759. https://doi.org/10.3390/microorganisms11030759
Schmitz A, Hanke D, Lüschow D, Schwarz S, Higgins PG, Feßler AT. Acinetobacter baumannii from Samples of Commercially Reared Turkeys: Genomic Relationships, Antimicrobial and Biocide Susceptibility. Microorganisms. 2023; 11(3):759. https://doi.org/10.3390/microorganisms11030759
Chicago/Turabian StyleSchmitz, Anna, Dennis Hanke, Dörte Lüschow, Stefan Schwarz, Paul G. Higgins, and Andrea T. Feßler. 2023. "Acinetobacter baumannii from Samples of Commercially Reared Turkeys: Genomic Relationships, Antimicrobial and Biocide Susceptibility" Microorganisms 11, no. 3: 759. https://doi.org/10.3390/microorganisms11030759