
Novel Insights on Extracellular Electron Transfer Networks in the Desulfovibrionaceae Family: Unveiling the Potential Significance of Horizontal Gene Transfer

 , and
, and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Genome Selection and Phylogenomic Analysis
2.2. Creation of Custom Databases: Genes Related to Extracellular Electron Transfer (EET)
2.3. Ecophysiological Analysis of SRB and FeRB
2.4. Comparative Genomics
2.5. Similarity Network
2.6. Phylogenetic Analysis and Comparison of Genetic Contexts
2.7. Statistical Analysis
3. Results and Discussion
3.1. Phylogenomic Analysis of Sulfate-Reducing and Iron-Reducing Metabolism
3.1.1. Abundance of Multi-Heme Cytochromes
3.1.2. Similarity Network Analysis of Extracellular Multi-Heme Cytochromes
3.1.3. Comparative Genomic Analysis of Genes Related to Extracellular Electron Transfer Mechanisms
Similarities with the EET Mechanism of G. sulfurreducens
Similarities with the EET Mechanism of Shewanella oneidensis
3.2. Mobilome Analyses across Members of Desulfovibrionaceae, Geobacteraceae, and Shewanellaceae Families
3.3. Evolutionary Relationship of the Most Prevalent EET-Related Genes in SRPs
3.3.1. The Periplasmic Cytochrome PpcA, an Intermediary in Extracellular Electron Transfer
3.3.2. OmpJ, an Integral and Widespread Outer Membrane Protein in the Desulfovibrionaceae Family
3.3.3. CymA, a Common Branch Point in the Electron Transport Chain
3.3.4. Inner-Membrane Quinone Oxidoreductase Protein Complexes: CbcC and CbcT Subunits Provide Plasticity and Modularity to Different Complexes Involved in EET
4. Implications
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Bao, P.; Li, G.X.; Sun, G.X.; Xu, Y.Y.; Meharg, A.A.; Zhu, Y.G. The role of sulfate-reducing prokaryotes in the coupling of element biogeochemical cycling. Sci. Total Environ. 2018, 613–614, 398–408. [Google Scholar] [CrossRef] [PubMed]
- Yu, S.-S.; Chen, J.-J.; Cheng, R.-F.; Min, Y.; Yu, H.-Q. Iron Cycle Tuned by Outer-Membrane Cytochromes of Dissimilatory Metal-Reducing Bacteria: Interfacial Dynamics and Mechanisms In Vitro. Environ. Sci. Technol. 2021, 55, 11424–11433. [Google Scholar] [CrossRef]
- Wunder, L.C.; Aromokeye, D.A.; Yin, X.; Richter-Heitmann, T.; Willis-Poratti, G.; Schnakenberg, A.; Otersen, C.; Dohrmann, I.; Römer, M.; Bohrmann, G.; et al. Iron and sulfate reduction structure microbial communities in (sub-)Antarctic sediments. ISME J. 2021, 15, 3587–3604. [Google Scholar] [CrossRef]
- Berg, J.S.; Jézéquel, D.; Duverger, A.; Lamy, D.; Laberty-Robert, C.; Miot, J. Microbial diversity involved in iron and cryptic sulfur cycling in the ferruginous, low-sulfate waters of Lake Pavin. PLoS ONE 2019, 14, e0212787. [Google Scholar] [CrossRef] [PubMed]
- Hu, K.; Xu, D.; Chen, Y. An assessment of sulfate reducing bacteria on treating sulfate-rich metal-laden wastewater from electroplating plant. J. Hazard. Mater. 2020, 393, 122376. [Google Scholar] [CrossRef] [PubMed]
- Kovacik, W.P., Jr.; Takai, K.; Mormile, M.R.; McKinley, J.P.; Brockman, F.J.; Fredrickson, J.K.; Holben, W.E. Molecular analysis of deep subsurface Cretaceous rock indicates abundant Fe(III)- and S°-reducing bacteria in a sulfate-rich environment. Environ. Microbiol. 2006, 8, 141–155. [Google Scholar] [CrossRef] [PubMed]
- Muyzer, G.; Stams, A.J.M. The ecology and biotechnology of sulphate-reducing bacteria. Nat. Rev. Microbiol. 2008, 6, 441–454. [Google Scholar] [CrossRef]
- Rey, F.E.; Gonzalez, M.D.; Cheng, J.; Wu, M.; Ahern, P.P.; Gordon, J.I. Metabolic niche of a prominent sulfate-reducing human gut bacterium. Proc. Natl. Acad. Sci. USA 2013, 110, 13582–13587. [Google Scholar] [CrossRef]
- Reyes, C.; Dellwig, O.; Dähnke, K.; Gehre, M.; Noriega-Ortega, B.E.; Böttcher, M.E.; Meister, P.; Friedrich, M.W. Bacterial communities potentially involved in iron-cycling in Baltic Sea and North Sea sediments revealed by pyrosequencing. FEMS Microbiol. Ecol. 2016, 92, fiw054. [Google Scholar] [CrossRef]
- Buongiorno, J.; Herbert, L.C.; Wehrmann, L.M.; Michaud, A.B.; Laufer, K.; Røy, H.; Jørgensen, B.B.; Szynkiewicz, A.; Faiia, A.; Yeager, K.M.; et al. Complex Microbial Communities Drive Iron and Sulfur Cycling in Arctic Fjord Sediments. Appl. Environ. Microbiol. 2019, 85, e00949-19. [Google Scholar] [CrossRef]
- Chapelle, F.H.; Lovley, D.R. Competitive Exclusion of Sulfate Reduction by Fe(lll)-Reducing Bacteria: A Mechanism for Producing Discrete Zones of High-Iron Ground Water. Groundwater 1992, 30, 29–36. [Google Scholar] [CrossRef]
- Ayangbenro, A.S.; Olanrewaju, O.S.; Babalola, O.O. Sulfate-Reducing Bacteria as an Effective Tool for Sustainable Acid Mine Bioremediation. Front. Microbiol. 2018, 9, 1986. [Google Scholar] [CrossRef] [PubMed]
- Canfield, D.E.; Jørgensen, B.B.; Fossing, H.; Glud, R.; Gundersen, J.; Ramsing, N.B.; Thamdrup, B.; Hansen, J.W.; Nielsen, L.P.; Hall, P.O.J. Pathways of organic carbon oxidation in three continental margin sediments. Mar. Geol. 1993, 113, 27–40. [Google Scholar] [CrossRef]
- Canfield, D.E.; Thamdrup, B.; Hansen, J.W. The anaerobic degradation of organic matter in Danish coastal sediments: Iron reduction, manganese reduction, and sulfate reduction. Geochim. Cosmochim. Acta 1993, 57, 3867–3883. [Google Scholar] [CrossRef]
- Xing, C.; Lang, X.; Ma, H.; Peng, Y.; Peng, Y.; Liu, Y.; Wang, R.; Ning, M.; Cui, Y.; Yu, X.; et al. Predominant microbial iron reduction in sediment in early Cambrian sulfidic oceans. Glob. Planet. Chang. 2021, 206, 103637. [Google Scholar] [CrossRef]
- Bowles, M.W.; Mogollón, J.M.; Kasten, S.; Zabel, M.; Hinrichs, K.-U. Global rates of marine sulfate reduction and implications for sub–sea-floor metabolic activities. Science 2014, 344, 889–891. [Google Scholar] [CrossRef]
- Dong, H.; Zeng, Q.; Sheng, Y.; Chen, C.; Yu, G.; Kappler, A. Coupled iron cycling and organic matter transformation across redox interfaces. Nat. Rev. Earth Environ. 2023, 4, 659–673. [Google Scholar] [CrossRef]
- Plugge, C.; Zhang, W.; Scholten, J.; Stams, A. Metabolic Flexibility of Sulfate-Reducing Bacteria. Front. Microbiol. 2011, 2, 81. [Google Scholar] [CrossRef]
- Jørgensen, B.B. Mineralization of organic matter in the sea bed—The role of sulphate reduction. Nature 1982, 296, 643–645. [Google Scholar] [CrossRef]
- Jørgensen, B.B.; Kasten, S. Sulfur Cycling and Methane Oxidation. In Marine Geochemistry; Schulz, H.D., Zabel, M., Eds.; Springer: Berlin/Heidelberg, Germany, 2006; pp. 271–309. [Google Scholar]
- Fike, D.A.; Bradley, A.S.; Rose, C.V. Rethinking the Ancient Sulfur Cycle. Annu. Rev. Earth Planet. Sci. 2015, 43, 593–622. [Google Scholar] [CrossRef]
- Raven, M.R.; Keil, R.G.; Webb, S.M. Microbial sulfate reduction and organic sulfur formation in sinking marine particles. Science 2021, 371, 178–181. [Google Scholar] [CrossRef]
- Lovley, D.R.; Holmes, D.E.; Nevin, K.P. Dissimilatory Fe(III) and Mn(IV) reduction. Adv. Microb. Physiol. 2004, 49, 219–286. [Google Scholar] [CrossRef] [PubMed]
- Jickells, T.D.; An, Z.S.; Andersen, K.K.; Baker, A.R.; Bergametti, G.; Brooks, N.; Cao, J.J.; Boyd, P.W.; Duce, R.A.; Hunter, K.A.; et al. Global iron connections between desert dust, ocean biogeochemistry, and climate. Science 2005, 308, 67–71. [Google Scholar] [CrossRef]
- Champ, D.; Gulens, J.; Jackson, R. Oxidation-Reduction Sequences in Ground Water Flow Systems. Can. J. Earth Sci. 2011, 16, 12–23. [Google Scholar] [CrossRef]
- Lovley, D.R.; Chapelle, F.H.; Woodward, J.C. Use of Dissolved H2 Concentrations To Determine Distribution of Microbially Catalyzed Redox Reactions in Anoxic Groundwater. Environ. Sci. Technol. 1994, 28, 1205–1210. [Google Scholar] [CrossRef] [PubMed]
- Bethke, C.; Sanford, R.; Kirk, M.; Jin, Q.; Flynn, T. The Thermodynamic ladder in Geomicrobiology. Am. J. Sci. 2011, 311, 183–210. [Google Scholar] [CrossRef]
- Lovley, D.R.; Goodwin, S. Hydrogen concentrations as an indicator of the predominant terminal electron-accepting reactions in aquatic sediments. Geochim. Cosmochim. Acta 1988, 52, 2993–3003. [Google Scholar] [CrossRef]
- Lovley, D.R.; Phillips, E.J.P. Competitive Mechanisms for Inhibition of Sulfate Reduction and Methane Production in the Zone of Ferric Iron Reduction in Sediments. Appl. Environ. Microbiol. 1987, 53, 2636–2641. [Google Scholar] [CrossRef]
- Kirk, M.F.; Jin, Q.; Haller, B.R. Broad-Scale Evidence That pH Influences the Balance Between Microbial Iron and Sulfate Reduction. Groundwater 2016, 54, 406–413. [Google Scholar] [CrossRef]
- Canfield, D.E.; Erik, K.; Bo, T. Thermodynamics and Microbial Metabolism. In Advances in Marine Biology; Canfield, D.E., Kristensen, E., Thamdrup, B., Eds.; Academic Press: Cambridge, MA, USA, 2005; Volume 48, pp. 65–94. [Google Scholar]
- Kappler, A.; Bryce, C.; Mansor, M.; Lueder, U.; Byrne, J.M.; Swanner, E.D. An evolving view on biogeochemical cycling of iron. Nat. Rev. Microbiol. 2021, 19, 360–374. [Google Scholar] [CrossRef]
- Jørgensen, B.B.; Findlay, A.J.; Pellerin, A. The Biogeochemical Sulfur Cycle of Marine Sediments. Front. Microbiol. 2019, 10, 849. [Google Scholar] [CrossRef]
- Sinninghe Damst’e, J.S.; Rijpstra, W.I.C.; Kock-van Dalen, A.C.; De Leeuw, J.W.; Schenck, P.A. Quenching of labile functionalised lipids by inorganic sulphur species: Evidence for the formation of sedimentary organic sulphur compounds at the early stages of diagenesis. Geochim. Cosmochim. Acta 1989, 53, 1343–1355. [Google Scholar] [CrossRef]
- Yu, Z.-G.; Peiffer, S.; Göttlicher, J.; Knorr, K.-H. Electron Transfer Budgets and Kinetics of Abiotic Oxidation and Incorporation of Aqueous Sulfide by Dissolved Organic Matter. Environ. Sci. Technol. 2015, 49, 5441–5449. [Google Scholar] [CrossRef] [PubMed]
- Heitmann, T.; Blodau, C. Oxidation and incorporation of hydrogen sulfide by dissolved organic matter. Chem. Geol. 2006, 235, 12–20. [Google Scholar] [CrossRef]
- Froelich, P.N.; Klinkhammer, G.P.; Bender, M.L.; Luedtke, N.A.; Heath, G.R.; Cullen, D.; Dauphin, P.; Hammond, D.; Hartman, B.; Maynard, V. Early oxidation of organic matter in pelagic sediments of the eastern equatorial Atlantic: Suboxic diagenesis. Geochim. Cosmochim. Acta 1979, 43, 1075–1090. [Google Scholar] [CrossRef]
- Hoehler, T.M.; Alperin, M.J.; Albert, D.B.; Martens, C.S. Thermodynamic control on hydrogen concentrations in anoxic sediments. Geochim. Cosmochim. Acta 1998, 62, 1745–1756. [Google Scholar] [CrossRef]
- Coleman, M.L.; Hedrick, D.B.; Lovley, D.R.; White, D.C.; Pye, K. Reduction of Fe(III) in sediments by sulphate-reducing bacteria. Nature 1993, 361, 436–438. [Google Scholar] [CrossRef]
- Ke, C.; Guo, C.; Zhang, S.; Deng, Y.; Li, X.; Li, Y.; Lu, G.; Ling, F.; Dang, Z. Microbial reduction of schwertmannite by co-cultured iron- and sulfate-reducing bacteria. Sci. Total Environ. 2023, 861, 160551. [Google Scholar] [CrossRef] [PubMed]
- Lovley, D.R.; Roden, E.E.; Phillips, E.J.P.; Woodward, J.C. Enzymatic iron and uranium reduction by sulfate-reducing bacteria. Mar. Geol. 1993, 113, 41–53. [Google Scholar] [CrossRef]
- Holmes, D.E.; Bond, D.R.; Lovley, D.R. Electron transfer by Desulfobulbus propionicus to Fe(III) and graphite electrodes. Appl. Environ. Microbiol. 2004, 70, 1234–1237. [Google Scholar] [CrossRef] [PubMed]
- Park, H.S.; Lin, S.; Voordouw, G. Ferric iron reduction by Desulfovibrio vulgaris Hildenborough wild type and energy metabolism mutants. Antonie Van Leeuwenhoek 2008, 93, 79–85. [Google Scholar] [CrossRef] [PubMed]
- Dalla Vecchia, E.; Shao, P.P.; Suvorova, E.; Chiappe, D.; Hamelin, R.; Bernier-Latmani, R. Characterization of the surfaceome of the metal-reducing bacterium Desulfotomaculum reducens. Front. Microbiol. 2014, 5, 432. [Google Scholar] [CrossRef] [PubMed]
- Brown, S.D.; Utturkar, S.M.; Arkin, A.P.; Deutschbauer, A.M.; Elias, D.A.; Hazen, T.C.; Chakraborty, R. Draft Genome Sequence for Desulfovibrio africanus Strain PCS. Genome Announc. 2013, 1, e00144-13. [Google Scholar] [CrossRef]
- Vandieken, V.; Knoblauch, C.; Jørgensen, B.B. Desulfovibrio frigidus sp. nov. and Desulfovibrio ferrireducens sp. nov., psychrotolerant bacteria isolated from Arctic fjord sediments (Svalbard) with the ability to reduce Fe(III). Int. J. Syst. Evol. Microbiol. 2006, 56, 681–685. [Google Scholar] [CrossRef]
- Badalamenti, J.P.; Summers, Z.M.; Chan, C.H.; Gralnick, J.A.; Bond, D.R. Isolation and Genomic Characterization of ‘Desulfuromonas soudanensis WTL’, a Metal- and Electrode-Respiring Bacterium from Anoxic Deep Subsurface Brine. Front. Microbiol. 2016, 7, 913. [Google Scholar] [CrossRef]
- Liang, D.; Liu, X.; Woodard, T.L.; Holmes, D.E.; Smith, J.A.; Nevin, K.P.; Feng, Y.; Lovley, D.R. Extracellular Electron Exchange Capabilities of Desulfovibrio ferrophilus and Desulfopila corrodens. Environ. Sci. Technol. 2021, 55, 16195–16203. [Google Scholar] [CrossRef]
- Enning, D.; Garrelfs, J. Corrosion of iron by sulfate-reducing bacteria: New views of an old problem. Appl. Environ. Microbiol. 2014, 80, 1226–1236. [Google Scholar] [CrossRef]
- Ueki, T.; Lovley, D.R. Desulfovibrio vulgaris as a model microbe for the study of corrosion under sulfate-reducing conditions. mLife 2022, 1, 13–20. [Google Scholar] [CrossRef]
- Ayala Nuñez, T.; Cerbino, G.N.; Rapisardi, M.F.; Quiroga, C.; Centrón, D. Novel Mobile Integrons and Strain-Specific Integrase Genes within Shewanella spp. Unveil Multiple Lateral Genetic Transfer Events within the Genus. Microorganisms 2022, 10, 1102. [Google Scholar] [CrossRef] [PubMed]
- Cerbino, G.N.; Traglia, G.M.; Ayala Nuñez, T.; Parmeciano Di Noto, G.; Ramírez, M.S.; Centrón, D.; Iriarte, A.; Quiroga, C. Comparative genome analysis of the genus Shewanella unravels the association of key genetic traits with known and potential pathogenic lineages. Front. Microbiol. 2023, 14, 1124225. [Google Scholar] [CrossRef]
- Orellana, R.; Arancibia, A.; Badilla, L.; Acosta, J.; Arancibia, G.; Escar, R.; Ferrada, G.; Seeger, M. Ecophysiological Features Shape the Distribution of Prophages and CRISPR in Sulfate Reducing Prokaryotes. Microorganisms 2021, 9, 931. [Google Scholar] [CrossRef] [PubMed]
- Holmes, D.E.; Giloteaux, L.; Chaurasia, A.K.; Williams, K.H.; Luef, B.; Wilkins, M.J.; Wrighton, K.C.; Thompson, C.A.; Comolli, L.R.; Lovley, D.R. Evidence of Geobacter-associated phage in a uranium-contaminated aquifer. ISME J. 2015, 9, 333–346. [Google Scholar] [CrossRef] [PubMed]
- Crispim, J.S.; Dias, R.S.; Vidigal, P.M.P.; de Sousa, M.P.; da Silva, C.C.; Santana, M.F.; de Paula, S.O. Screening and characterization of prophages in Desulfovibrio genomes. Sci. Rep. 2018, 8, 9273. [Google Scholar] [CrossRef] [PubMed]
- Hernández, S.; Vives, M.J. Phages in Anaerobic Systems. Viruses 2020, 12, 1091. [Google Scholar] [CrossRef]
- Caro-Quintero, A.; Deng, J.; Auchtung, J.; Brettar, I.; Höfle, M.G.; Klappenbach, J.; Konstantinidis, K.T. Unprecedented levels of horizontal gene transfer among spatially co-occurring Shewanella bacteria from the Baltic Sea. ISME J. 2011, 5, 131–140. [Google Scholar] [CrossRef]
- Zhong, C.; Han, M.; Yu, S.; Yang, P.; Li, H.; Ning, K. Pan-genome analyses of 24 Shewanella strains re-emphasize the diversification of their functions yet evolutionary dynamics of metal-reducing pathway. Biotechnol. Biofuels 2018, 11, 193. [Google Scholar] [CrossRef]
- Baker, I.R.; Conley, B.E.; Gralnick, J.A.; Girguis, P.R. Evidence for Horizontal and Vertical Transmission of Mtr-Mediated Extracellular Electron Transfer among the Bacteria. mBio 2021, 13, e0290421. [Google Scholar] [CrossRef]
- Kumar, A.; Hsu, L.H.-H.; Kavanagh, P.; Barrière, F.; Lens, P.N.L.; Lapinsonnière, L.; Lienhard, V.J.H.; Schröder, U.; Jiang, X.; Leech, D. The ins and outs of microorganism–electrode electron transfer reactions. Nat. Rev. Chem. 2017, 1, 0024. [Google Scholar] [CrossRef]
- Shi, L.; Dong, H.; Reguera, G.; Beyenal, H.; Lu, A.; Liu, J.; Yu, H.-Q.; Fredrickson, J.K. Extracellular electron transfer mechanisms between microorganisms and minerals. Nat. Rev. Microbiol. 2016, 14, 651–662. [Google Scholar] [CrossRef]
- Cai, X.; Huang, L.; Yang, G.; Yu, Z.; Wen, J.; Zhou, S. Transcriptomic, Proteomic, and Bioelectrochemical Characterization of an Exoelectrogen Geobacter soli Grown with Different Electron Acceptors. Front. Microbiol. 2018, 9, 1075. [Google Scholar] [CrossRef]
- Antunes, J.M.A.; Silva, M.A.; Salgueiro, C.A.; Morgado, L. Electron Flow From the Inner Membrane Towards the Cell Exterior in Geobacter sulfurreducens: Biochemical Characterization of Cytochrome CbcL. Front. Microbiol. 2022, 13, 898015. [Google Scholar] [CrossRef] [PubMed]
- Ueki, T. Cytochromes in Extracellular Electron Transfer in Geobacter. Appl. Environ. Microbiol. 2021, 87, e03109-20. [Google Scholar] [CrossRef]
- Hau, H.H.; Gralnick, J.A. Ecology and Biotechnology of the Genus Shewanella. Annu. Rev. Microbiol. 2007, 61, 237–258. [Google Scholar] [CrossRef] [PubMed]
- Parks, D.H.; Imelfort, M.; Skennerton, C.T.; Hugenholtz, P.; Tyson, G.W. CheckM: Assessing the quality of microbial genomes recovered from isolates, single cells, and metagenomes. Genome Res. 2015, 25, 1043–1055. [Google Scholar] [CrossRef] [PubMed]
- Emms, D.M.; Kelly, S. OrthoFinder: Phylogenetic orthology inference for comparative genomics. Genome Biol. 2019, 20, 238. [Google Scholar] [CrossRef]
- Katoh, K.; Standley, D.M. MAFFT multiple sequence alignment software version 7: Improvements in performance and usability. Mol. Biol. Evol. 2013, 30, 772–780. [Google Scholar] [CrossRef]
- Price, M.N.; Dehal, P.S.; Arkin, A.P. FastTree 2—Approximately Maximum-Likelihood Trees for Large Alignments. PLoS ONE 2010, 5, e9490. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular Evolutionary Genetics Analysis across Computing Platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef]
- Letunic, I.; Bork, P. Interactive Tree Of Life (iTOL) v5: An online tool for phylogenetic tree display and annotation. Nucleic Acids Res. 2021, 49, W293–W296. [Google Scholar] [CrossRef]
- Kuever, J.; Rainey, F.A.; Widdel, F. Desulfovibrio. In Bergey’s Manual of Systematics of Archaea and Bacteria; Wiley: Hoboken, NJ, USA, 2015; pp. 1–17. [Google Scholar]
- Campbell, L.L.; Kasprzycki, M.A.; Postgate, J.R. Desulfovibrio africanus sp. n., a New Dissimilatory Sulfate-reducing Bacterium. J. Bacteriol. 1966, 92, 1122–1127. [Google Scholar] [CrossRef]
- Motamedi, M.; Pedersen, K. Desulfovibrio aespoeensis sp. nov., a mesophilic sulfate-reducing bacterium from deep groundwater at Äspö hard rock laboratory, Sweden. Int. J. Syst. Evol. Microbiol. 1998, 48, 311–315. [Google Scholar] [CrossRef]
- Sakaguchi, T.; Arakaki, A.; Matsunaga, T. Desulfovibrio magneticus sp. nov., a novel sulfate-reducing bacterium that produces intracellular single-domain-sized magnetite particles. Int. J. Syst. Evol. Microbiol. 2002, 52, 215–221. [Google Scholar] [CrossRef] [PubMed]
- Legall, J. A New species of Desulfovibrio. J. Bacteriol. 1963, 86, 1120. [Google Scholar] [CrossRef] [PubMed]
- Loubinoux, J.; Valente, F.M.A.; Pereira, I.A.C.; Costa, A.; Grimont, P.A.D.; Le Faou, A.E. Reclassification of the only species of the genus Desulfomonas, Desulfomonas pigra, as Desulfovibrio piger comb. nov. Int. J. Syst. Evol. Microbiol. 2002, 52, 1305–1308. [Google Scholar] [CrossRef] [PubMed]
- Krekeler, D.; Sigalevich, P.; Teske, A.; Cypionka, H.; Cohen, Y. A sulfate-reducing bacterium from the oxic layer of a microbial mat from Solar Lake (Sinai), Desulfovibrio oxyclinae sp. nov. Arch. Microbiol. 1997, 167, 369–375. [Google Scholar] [CrossRef]
- Sass, H.; Berchtold, M.; Branke, J.; König, H.; Cypionka, H.; Babenzien, H.-D. Psychrotolerant Sulfate-reducing Bacteria from an Oxic Freshwater Sediment Description of Desulfovibrio cuneatus sp. nov. and Desulfovibrio litoralis sp. nov. Syst. Appl. Microbiol. 1998, 21, 212–219. [Google Scholar] [CrossRef]
- Klouche, N.; Basso, O.; Lascourrèges, J.-F.; Cayol, J.-L.; Thomas, P.; Fauque, G.; Fardeau, M.-L.; Magot, M. Desulfocurvus vexinensis gen. nov., sp. nov., a sulfate-reducing bacterium isolated from a deep subsurface aquifer. Int. J. Syst. Evol. Microbiol. 2009, 59, 3100–3104. [Google Scholar] [CrossRef]
- Abildgaard, L.; Nielsen, M.B.; Kjeldsen, K.U.; Ingvorsen, K. Desulfovibrio alkalitolerans sp. nov., a novel alkalitolerant, sulphate-reducing bacterium isolated from district heating water. Int. J. Syst. Evol. Microbiol. 2006, 56, 1019–1024. [Google Scholar] [CrossRef]
- Ollivier, B.; Cord-Ruwisch, R.; Hatchikian, E.C.; Garcia, J.L. Characterization of Desulfovibrio fructosovorans sp. nov. Arch. Microbiol. 1988, 149, 447–450. [Google Scholar] [CrossRef]
- Alazard, D.; Dukan, S.; Urios, A.; Verhé, F.; Bouabida, N.; Morel, F.; Thomas, P.; Garcia, J.-L.; Ollivier, B. Desulfovibrio hydrothermalis sp. nov., a novel sulfate-reducing bacterium isolated from hydrothermal vents. Int. J. Syst. Evol. Microbiol. 2003, 53, 173–178. [Google Scholar] [CrossRef]
- Brown, S.D.; Hurt, R.A., Jr.; Gilmour, C.C.; Elias, D.A. Draft genome sequences for three mercury-methylating, sulfate-reducing bacteria. Genome Announc. 2013, 1, e00618-13. [Google Scholar] [CrossRef] [PubMed]
- Karnachuk, O.V.; Sasaki, K.; Gerasimchuk, A.L.; Sukhanova, O.; Ivasenko, D.A.; Kaksonen, A.H.; Puhakka, J.A.; Tuovinen, O.H. Precipitation of Cu-Sulfides by Copper-Tolerant Desulfovibrio Isolates. Geomicrobiol. J. 2008, 25, 219–227. [Google Scholar] [CrossRef]
- Mancini, S.; Abicht, H.K.; Karnachuk, O.V.; Solioz, M. Genome sequence of Desulfovibrio sp. A2, a highly copper resistant, sulfate-reducing bacterium isolated from effluents of a zinc smelter at the Urals. J. Bacteriol. 2011, 193, 6793–6794. [Google Scholar] [CrossRef] [PubMed]
- Baena, S.; Fardeau, M.L.; Labat, M.; Ollivier, B.; Garcia, J.L.; Patel, B.K. Desulfovibrio aminophilus sp. nov., a novel amino acid degrading and sulfate reducing bacterium from an anaerobic dairy wastewater lagoon. Syst. Appl. Microbiol. 1998, 21, 498–504. [Google Scholar] [CrossRef] [PubMed]
- Khelaifia, S.; Fardeau, M.-L.; Pradel, N.; Aussignargues, C.; Garel, M.; Tamburini, C.; Cayol, J.-L.; Gaudron, S.; Gaill, F.; Ollivier, B. Desulfovibrio piezophilus sp. nov., a piezophilic, sulfate-reducing bacterium isolated from wood falls in the Mediterranean Sea. Int. J. Syst. Evol. Microbiol. 2011, 61, 2706–2711. [Google Scholar] [CrossRef]
- Feio, M.J.; Zinkevich, V.; Beech, I.B.; Llobet-Brossa, E.; Eaton, P.; Schmitt, J.; Guezennec, J. Desulfovibrio alaskensis sp. nov., a sulphate-reducing bacterium from a soured oil reservoir. Int. J. Syst. Evol. Microbiol. 2004, 54, 1747–1752. [Google Scholar] [CrossRef]
- Ramsay, B.D.; Hwang, C.; Woo, H.L.; Carroll, S.L.; Lucas, S.; Han, J.; Lapidus, A.L.; Cheng, J.F.; Goodwin, L.A.; Pitluck, S.; et al. High-Quality Draft Genome Sequence of Desulfovibrio carbinoliphilus FW-101-2B, an Organic Acid-Oxidizing Sulfate-Reducing Bacterium Isolated from Uranium(VI)-Contaminated Groundwater. Genome Announc. 2015, 3, e00092-15. [Google Scholar] [CrossRef]
- Zhou, J.; He, Q.; Hemme, C.L.; Mukhopadhyay, A.; Hillesland, K.; Zhou, A.; He, Z.; Van Nostrand, J.D.; Hazen, T.C.; Stahl, D.A.; et al. How sulphate-reducing microorganisms cope with stress: Lessons from systems biology. Nat. Rev. Microbiol. 2011, 9, 452–466. [Google Scholar] [CrossRef]
- Magot, M.; Basso, O.; Tardy-Jacquenod, C.; Caumette, P. Desulfovibrio bastinii sp. nov. and Desulfovibrio gracilis sp. nov., moderately halophilic, sulfate-reducing bacteria isolated from deep subsurface oilfield water. Int. J. Syst. Evol. Microbiol. 2004, 54, 1693–1697. [Google Scholar] [CrossRef]
- Qatibi, A.-I.; Nivière, V.; Garcia, J. Desulfovibrio alcoholovorans sp. nov., a sulfate-reducing bacterium able to grow on glycerol, 1,2- and 1,3-propanediol. Arch. Microbiol. 1991, 155, 143–148. [Google Scholar] [CrossRef]
- Nielsen, J.T.; Liesack, W.; Finster, K. Desulfovibrio zosterae sp. nov., a new sulfate reducer isolated from surface-sterilized roots of the seagrass Zostera marina. Int. J. Syst. Evol. Microbiol. 1999, 49, 859–865. [Google Scholar] [CrossRef]
- Basso, O.; Caumette, P.; Magot, M. Desulfovibrio putealis sp. nov., a novel sulfate-reducing bacterium isolated from a deep subsurface aquifer. Int. J. Syst. Evol. Microbiol. 2005, 55, 101–104. [Google Scholar] [CrossRef] [PubMed]
- Reichenbecher, W.; Schink, B. Desulfovibrio inopinatus, sp. nov., a new sulfate-reducing bacterium that degrades hydroxyhydroquinone (1,2,4-trihydroxybenzene). Arch. Microbiol. 1997, 168, 338–344. [Google Scholar] [CrossRef] [PubMed]
- Magot, M.; Caumette, P.; Desperrier, J.M.; Matheron, R.; Dauga, C.; Grimont, F.; Carreau, L. Desulfovibrio longus sp. nov., a Sulfate-Reducing Bacterium Isolated from an Oil-Producing Well. Int. J. Syst. Evol. Microbiol. 1992, 42, 398–402. [Google Scholar] [CrossRef] [PubMed]
- Gilmour, C.C.; Soren, A.B.; Gionfriddo, C.M.; Podar, M.; Wall, J.D.; Brown, S.D.; Michener, J.K.; Urriza, M.S.G.; Elias, D.A. Pseudodesulfovibrio mercurii sp. nov., a mercury-methylating bacterium isolated from sediment. Int. J. Syst. Evol. Microbiol. 2021, 71, 004697. [Google Scholar] [CrossRef]
- Wolicka, D.; Jarzynowska, L. Microbiological Reduction of Sulphates in Salty Environments and Mineralogical Characterization of the Transformation Products. Geomicrobiol. J. 2012, 29, 528–536. [Google Scholar] [CrossRef]
- Karnachuk, O.V.; Rusanov, I.I.; Panova, I.A.; Grigoriev, M.A.; Zyusman, V.S.; Latygolets, E.A.; Kadyrbaev, M.K.; Gruzdev, E.V.; Beletsky, A.V.; Mardanov, A.V.; et al. Microbial sulfate reduction by Desulfovibrio is an important source of hydrogen sulfide from a large swine finishing facility. Sci. Rep. 2021, 11, 10720. [Google Scholar] [CrossRef]
- Boeckman, J.; Korn, A.; Yao, G.; Ravindran, A.; Gonzalez, C.; Gill, J. Sheep in wolves’ clothing: Temperate T7-like bacteriophages and the origins of the Autographiviridae. Virology 2022, 568, 86–100. [Google Scholar] [CrossRef]
- Hatchikian, E.C.; Forget, N.; Fernandez, V.M.; Williams, R.; Cammack, R. Further characterization of the [Fe]-hydrogenase from Desulfovibrio desulfuricans ATCC 7757. Eur. J. Biochem. 1992, 209, 357–365. [Google Scholar] [CrossRef]
- Day, L.A.; León, K.B.D.; Kempher, M.L.; Zhou, J.; Wall, J.D. Complete Genome Sequence of Desulfovibrio desulfuricans IC1, a Sulfonate-Respiring Anaerobe. Microbiol. Resour. Announc. 2019, 8, 10–1128. [Google Scholar] [CrossRef]
- Lie, T.J.; Pitta, T.; Leadbetter, E.R.; Godchaux Iii, W.; Leadbetter, J.R. Sulfonates: Novel electron acceptors in anaerobic respiration. Arch. Microbiol. 1996, 166, 204–210. [Google Scholar] [CrossRef]
- Sheik, C.S.; Sieber, J.R.; Badalamenti, J.P.; Carden, K.; Olson, A. Complete Genome Sequence of Desulfovibrio desulfuricans Strain G11, a Model Sulfate-Reducing, Hydrogenotrophic, and Syntrophic Partner Organism. Genome Announc. 2017, 5, 10–1128. [Google Scholar] [CrossRef] [PubMed]
- McInerney, M.J.; Bryant, M.P.; Pfennig, N. Anaerobic bacterium that degrades fatty acids in syntrophic association with methanogens. Arch. Microbiol. 1979, 122, 129–135. [Google Scholar] [CrossRef]
- Karnachuk, O.V.; Ikkert, O.P.; Avakyan, M.R.; Knyazev, Y.V.; Volochaev, N.M.; Zyusman, V.S.; Panov, V.L.; Kadnikov, V.V.; Mardanov, A.V.; Ravin, N.V. Desulfovibrio desulfuricans AY5 Isolated from a Patient with Autism Spectrum Disorder Binds Iron in Low-Soluble Greigite and Pyrite. Microorganisms 2021, 9, 2558. [Google Scholar] [CrossRef]
- Sun, D.; Wang, A.; Cheng, S.; Yates, M.; Logan, B.E. Geobacter anodireducens sp. nov., an exoelectrogenic microbe in bioelectrochemical systems. Int. J. Syst. Evol. Microbiol. 2014, 64, 3485–3491. [Google Scholar] [CrossRef]
- Shelobolina, E.S.; Nevin, K.P.; Blakeney-Hayward, J.D.; Johnsen, C.V.; Plaia, T.W.; Krader, P.; Woodard, T.; Holmes, D.E.; VanPraagh, C.G.; Lovley, D.R. Geobacter pickeringii sp. nov., Geobacter argillaceus sp. nov. and Pelosinus fermentans gen. nov., sp. nov., isolated from subsurface kaolin lenses. Int. J. Syst. Evol. Microbiol. 2007, 57, 126–135. [Google Scholar] [CrossRef] [PubMed]
- Coates, J.D.; Bhupathiraju, V.K.; Achenbach, L.A.; Mclnerney, M.J.; Lovley, D.R. Geobacter hydrogenophilus, Geobacter chapellei and Geobacter grbiciae, three new, strictly anaerobic, dissimilatory Fe(III)-reducers. Int. J. Syst. Evol. Microbiol. 2001, 51, 581–588. [Google Scholar] [CrossRef]
- Prakash, O.; Gihring, T.M.; Dalton, D.D.; Chin, K.-J.; Green, S.J.; Akob, D.M.; Wanger, G.; Kostka, J.E. Geobacter daltonii sp. nov., an Fe(III)- and uranium(VI)-reducing bacterium isolated from a shallow subsurface exposed to mixed heavy metal and hydrocarbon contamination. Int. J. Syst. Evol. Microbiol. 2010, 60, 546–553. [Google Scholar] [CrossRef] [PubMed]
- Viulu, S.; Nakamura, K.; Okada, Y.; Saitou, S.; Takamizawa, K. Geobacter luticola sp. nov., an Fe(III)-reducing bacterium isolated from lotus field mud. Int. J. Syst. Evol. Microbiol. 2013, 63, 442–448. [Google Scholar] [CrossRef]
- Childers, S.E.; Ciufo, S.; Lovley, D.R. Geobacter metallireducens accesses insoluble Fe(III) oxide by chemotaxis. Nature 2002, 416, 767–769. [Google Scholar] [CrossRef]
- Aklujkar, M.; Krushkal, J.; DiBartolo, G.; Lapidus, A.; Land, M.L.; Lovley, D.R. The genome sequence of Geobacter metallireducens: Features of metabolism, physiology and regulation common and dissimilar to Geobacter sulfurreducens. BMC Microbiol. 2009, 9, 109. [Google Scholar] [CrossRef] [PubMed]
- Lovley, D.R.; Phillips, E.J. Novel mode of microbial energy metabolism: Organic carbon oxidation coupled to dissimilatory reduction of iron or manganese. Appl. Environ. Microbiol. 1988, 54, 1472–1480. [Google Scholar] [CrossRef] [PubMed]
- Badalamenti, J.P.; Bond, D.R. Complete Genome of Geobacter pickeringii G13 T, a Metal-Reducing Isolate from Sedimentary Kaolin Deposits. Genome Announc. 2015, 3, e00038-15. [Google Scholar] [CrossRef]
- Yang, G.; Chen, S.; Zhou, S.; Liu, Y. Genome sequence of a dissimilatory Fe(III)-reducing bacterium Geobacter soli type strain GSS01(T). Stand Genom. Sci. 2015, 10, 118. [Google Scholar] [CrossRef]
- Guo, Y.; Aoyagi, T.; Hori, T. Draft Genome Sequences of the Ferric Iron-Reducing Geobacter sp. Strains AOG1 and AOG2, Isolated from Enrichment Cultures on Crystalline Iron(III) Oxides. Microbiol. Resour. Announc. 2021, 10, e0091321. [Google Scholar] [CrossRef]
- Hori, T.; Aoyagi, T.; Itoh, H.; Narihiro, T.; Oikawa, A.; Suzuki, K.; Ogata, A.; Friedrich, M.W.; Conrad, R.; Kamagata, Y. Isolation of microorganisms involved in reduction of crystalline iron(III) oxides in natural environments. Front. Microbiol. 2015, 6, 386. [Google Scholar] [CrossRef]
- Yadav, P.; Antony-Babu, S.; Hayes, E.; Healy, O.M.; Pan, D.; Yang, W.H.; Silver, W.L.; Anderson, C.L.; Voshall, A.; Fernando, S.C.; et al. Complete Genome Sequence of Geobacter sp. Strain FeAm09, a Moderately Acidophilic Soil Bacterium. Microbiol. Resour. Announc. 2021, 10, 10–1128. [Google Scholar] [CrossRef] [PubMed]
- Yang, G.; Li, Y.; Lin, A.; Zhuang, L. Geobacter benzoatilyticus sp. nov., a novel benzoate-oxidizing, iron-reducing bacterium isolated from petroleum contaminated soil. Int. J. Syst. Evol. Microbiol. 2022, 72, 005281. [Google Scholar] [CrossRef]
- Ehara, A.; Suzuki, H.; Amachi, S. Draft Genome Sequence of Geobacter sp. Strain OR-1, an Arsenate-Respiring Bacterium Isolated from Japanese Paddy Soil. Genome Announc. 2015, 3, e01478-14. [Google Scholar] [CrossRef]
- Ohtsuka, T.; Yamaguchi, N.; Makino, T.; Sakurai, K.; Kimura, K.; Kudo, K.; Homma, E.; Dong, D.T.; Amachi, S. Arsenic Dissolution from Japanese Paddy Soil by a Dissimilatory Arsenate-Reducing Bacterium Geobacter sp. OR-1. Environ. Sci. Technol. 2013, 47, 6263–6271. [Google Scholar] [CrossRef]
- Warashina, T.; Yamamura, S.; Suzuki, H.; Amachi, S.; Arakawa, K. Complete Genome Sequence of Geobacter sp. Strain SVR, an Antimonate-Reducing Bacterium Isolated from Antimony-Rich Mine Soil. Microbiol. Resour. Announc. 2021, 10, 10-1128. [Google Scholar] [CrossRef]
- Warashina, T.; Harada, M.; Nakajima, N.; Yamamura, S.; Tomita, M.; Suzuki, H.; Amachi, S. Draft Genome Sequence of Geobacter sp. Strain SVR, Isolated from Antimony Mine Soil. Microbiol. Resour. Announc. 2020, 9, 10–1128. [Google Scholar] [CrossRef] [PubMed]
- Yamamura, S.; Iida, C.; Kobayashi, Y.; Watanabe, M.; Amachi, S. Production of two morphologically different antimony trioxides by a novel antimonate-reducing bacterium, Geobacter sp. SVR. J. Hazard. Mater. 2021, 411, 125100. [Google Scholar] [CrossRef] [PubMed]
- Nagarajan, H.; Butler, J.E.; Klimes, A.; Qiu, Y.; Zengler, K.; Ward, J.; Young, N.D.; Methé, B.A.; Palsson, B.; Lovley, D.R.; et al. De Novo assembly of the complete genome of an enhanced electricity-producing variant of Geobacter sulfurreducens using only short reads. PLoS ONE 2010, 5, e10922. [Google Scholar] [CrossRef]
- Yi, H.; Nevin, K.P.; Kim, B.C.; Franks, A.E.; Klimes, A.; Tender, L.M.; Lovley, D.R. Selection of a variant of Geobacter sulfurreducens with enhanced capacity for current production in microbial fuel cells. Biosens. Bioelectron. 2009, 24, 3498–3503. [Google Scholar] [CrossRef]
- Caccavo, F., Jr.; Lonergan, D.J.; Lovley, D.R.; Davis, M.; Stolz, J.F.; McInerney, M.J. Geobacter sulfurreducens sp. nov., a hydrogen- and acetate-oxidizing dissimilatory metal-reducing microorganism. Appl. Environ. Microbiol. 1994, 60, 3752–3759. [Google Scholar] [CrossRef]
- Methé, B.A.; Nelson, K.E.; Eisen, J.A.; Paulsen, I.T.; Nelson, W.; Heidelberg, J.F.; Wu, D.; Wu, M.; Ward, N.; Beanan, M.J.; et al. Genome of Geobacter sulfurreducens: Metal reduction in subsurface environments. Science 2003, 302, 1967–1969. [Google Scholar] [CrossRef] [PubMed]
- Inoue, K.; Ogura, Y.; Kawano, Y.; Hayashi, T. Complete Genome Sequence of Geobacter sulfurreducens Strain YM18, Isolated from River Sediment in Japan. Genome Announc. 2018, 6, e00352-18. [Google Scholar] [CrossRef]
- Fujikawa, T.; Ogura, Y.; Ishigami, K.; Kawano, Y.; Nagamine, M.; Hayashi, T.; Inoue, K. Unexpected genomic features of high current density-producing Geobacter sulfurreducens strain YM18. FEMS Microbiol. Lett. 2021, 368, fnab119. [Google Scholar] [CrossRef]
- Fujikawa, T.; Ogura, Y.; Hayashi, T.; Inoue, K. Complete Genome Sequence of High Current-Producing Geobacter sulfurreducens Strain YM35, Isolated from River Sediment in Japan. Microbiol. Resour. Announc. 2021, 10, e0053921. [Google Scholar] [CrossRef]
- Shelobolina, E.S.; Vrionis, H.A.; Findlay, R.H.; Lovley, D.R. Geobacter uraniireducens sp. nov., isolated from subsurface sediment undergoing uranium bioremediation. Int. J. Syst. Evol. Microbiol. 2008, 58, 1075–1078. [Google Scholar] [CrossRef] [PubMed]
- Semple, K.M.; Westlake, D.W.S. Characterization of iron-reducing Alteromonas putrefaciens strains from oil field fluids. Can. J. Microbiol. 1987, 33, 366–371. [Google Scholar] [CrossRef]
- Kim, J.-Y.; Yoo, H.-S.; Lee, D.-H.; Park, S.-H.; Kim, Y.-J.; Oh, D.-C. Shewanella algicola sp. nov., a marine bacterium isolated from brown algae. Int. J. Syst. Evol. Microbiol. 2016, 66, 2218–2224. [Google Scholar] [CrossRef] [PubMed]
- Ohama, Y.; Aoki, K.; Harada, S.; Nagasawa, T.; Sawabe, T.; Nonaka, L.; Moriya, K.; Ishii, Y.; Tateda, K. Genetic Environment Surrounding blaOXA-55-like in Clinical Isolates of Shewanella algae Clade and Enhanced Expression of blaOXA-55-like in a Carbapenem-Resistant Isolate. mSphere 2021, 6, e00593-21. [Google Scholar] [CrossRef]
- Wang, Y.; Chen, H.; Liu, Z.; Ming, H.; Zhou, C.; Zhu, X.; Zhang, P.; Jing, C.; Feng, H. Shewanella gelidii sp. nov., isolated from the red algae Gelidium amansii, and emended description of Shewanella waksmanii. Int. J. Syst. Evol. Microbiol. 2016, 66, 2899–2905. [Google Scholar] [CrossRef] [PubMed]
- Satomi, M.; Vogel, B.F.; Venkateswaran, K.; Gram, L. Description of Shewanella glacialipiscicola sp. nov. and Shewanella algidipiscicola sp. nov., isolated from marine fish of the Danish Baltic Sea, and proposal that Shewanella affinis is a later heterotypic synonym of Shewanella colwelliana. Int. J. Syst. Evol. Microbiol. 2007, 57, 347–352. [Google Scholar] [CrossRef]
- Satomi, M.; Vogel, B.F.; Gram, L.; Venkateswaran, K. Shewanella hafniensis sp. nov. and Shewanella morhuae sp. nov., isolated from marine fish of the Baltic Sea. Int. J. Syst. Evol. Microbiol. 2006, 56, 243–249. [Google Scholar] [CrossRef] [PubMed]
- Prachumwat, A.; Wechprasit, P.; Srisala, J.; Kriangsaksri, R.; Flegel, T.W.; Thitamadee, S.; Sritunyalucksana, K. Shewanella khirikhana sp. nov.—A shrimp pathogen isolated from a cultivation pond exhibiting early mortality syndrome. Microb. Biotechnol. 2020, 13, 781–795. [Google Scholar] [CrossRef] [PubMed]
- Wechprasit, P.; Panphloi, M.; Thitamadee, S.; Sritunyalucksana, K.; Prachumwat, A. Complete Genome Sequence of Shewanella sp. Strain TH2012, Isolated from Shrimp in a Cultivation Pond Exhibiting Early Mortality Syndrome. Microbiol. Resour. Announc. 2019, 8, e01703-18. [Google Scholar] [CrossRef]
- Leonardo, M.R.; Moser, D.P.; Barbieri, E.; Brantner, C.A.; MacGregor, B.J.; Paster, B.J.; Stackebrandt, E.; Nealson, K.H. Shewanella pealeana sp. nov., a member of the microbial community associated with the accessory nidamental gland of the squid Loligo pealei. Int. J. Syst. Bacteriol. 1999, 49 Pt 4, 1341–1351. [Google Scholar] [CrossRef]
- Cha, Q.-Q.; Ren, X.-B.; Sun, Y.-Y.; He, X.-Y.; Su, H.-N.; Chen, X.-L.; Zhang, Y.-Z.; Xie, B.-B.; Zhao, L.-S.; Song, X.-Y.; et al. Shewanella polaris sp. nov., a psychrotolerant bacterium isolated from Arctic brown algae. Int. J. Syst. Evol. Microbiol. 2020, 70, 2096–2102. [Google Scholar] [CrossRef] [PubMed]
- Gai, Y.; Huang, Z.; Lai, Q.; Shao, Z. Shewanella intestini sp. nov., isolated from the intestine of abalone, Haliotis diversicolor. Int. J. Syst. Evol. Microbiol. 2017, 67, 1901–1905. [Google Scholar] [CrossRef] [PubMed]
- Satomi, M.; Oikawa, H.; Yano, Y. Shewanella marinintestina sp. nov., Shewanella schlegeliana sp. nov. and Shewanella sairae sp. nov., novel eicosapentaenoic-acid-producing marine bacteria isolated from sea-animal intestines. Int. J. Syst. Evol. Microbiol. 2003, 53, 491–499. [Google Scholar] [CrossRef] [PubMed]
- Brian-Jaisson, F.; Ortalo-Magné, A.; Guentas-Dombrowsky, L.; Armougom, F.; Blache, Y.; Molmeret, M. Identification of bacterial strains isolated from the Mediterranean Sea exhibiting different abilities of biofilm formation. Microb. Ecol. 2014, 68, 94–110. [Google Scholar] [CrossRef]
- Guillonneau, R.; Baraquet, C.; Bazire, A.; Molmeret, M. Multispecies Biofilm Development of Marine Bacteria Implies Complex Relationships Through Competition and Synergy and Modification of Matrix Components. Front. Microbiol. 2018, 9, 1960. [Google Scholar] [CrossRef]
- Caro-Quintero, A.; Auchtung, J.; Deng, J.; Brettar, I.; Höfle, M.; Tiedje, J.M.; Konstantinidis, K.T. Genome sequencing of five Shewanella baltica strains recovered from the oxic-anoxic interface of the Baltic Sea. J. Bacteriol. 2012, 194, 1236. [Google Scholar] [CrossRef]
- Deng, J.; Brettar, I.; Luo, C.; Auchtung, J.; Konstantinidis, K.T.; Rodrigues, J.L.; Höfle, M.; Tiedje, J.M. Stability, genotypic and phenotypic diversity of Shewanella baltica in the redox transition zone of the Baltic Sea. Environ. Microbiol. 2014, 16, 1854–1866. [Google Scholar] [CrossRef]
- Sung, H.-R.; Yoon, J.-H.; Ghim, S.-Y. Shewanella dokdonensis sp. nov., isolated from seawater. Int. J. Syst. Evol. Microbiol. 2012, 62, 1636–1643. [Google Scholar] [CrossRef] [PubMed]
- Reid, G.A.; Gordon, E.H.J. Phylogeny of marine and freshwater Shewanella: Reclassification of Shewanella putrefaciens NCIMB 400 as Shewanella frigidimarina. Int. J. Syst. Evol. Microbiol. 1999, 49, 189–191. [Google Scholar] [CrossRef]
- Reyes-Ramirez, F.; Dobbin, P.; Sawers, G.; Richardson David, J. Characterization of Transcriptional Regulation of Shewanella frigidimarina Fe(III)-Induced Flavocytochrome c Reveals a Novel Iron-Responsive Gene Regulation System. J. Bacteriol. 2003, 185, 4564–4571. [Google Scholar] [CrossRef]
- Kim, K.M.; Choe, H.; Kim, B.K.; Nasir, A. Complete genome of a metabolically-diverse marine bacterium Shewanella japonica KCTC 22435(T). Mar. Genom. 2017, 35, 39–42. [Google Scholar] [CrossRef]
- Biffinger, J.C.; Fitzgerald, L.A.; Ray, R.; Little, B.J.; Lizewski, S.E.; Petersen, E.R.; Ringeisen, B.R.; Sanders, W.C.; Sheehan, P.E.; Pietron, J.J.; et al. The utility of Shewanella japonica for microbial fuel cells. Bioresour. Technol. 2011, 102, 290–297. [Google Scholar] [CrossRef]
- Ivanova, E.P.; Sawabe, T.; Gorshkova, N.M.; Svetashev, V.I.; Mikhailov, V.V.; Nicolau, D.V.; Christen, R. Shewanella japonica sp. nov. Int. J. Syst. Evol. Microbiol. 2001, 51, 1027–1033. [Google Scholar] [CrossRef]
- Yun, B.-R.; Park, S.; Kim, M.-K.; Park, J.; Kim, S.B. Shewanella saliphila sp. nov., Shewanella ulleungensis sp. nov. and Shewanella litoralis sp. nov., isolated from coastal seawater. Int. J. Syst. Evol. Microbiol. 2018, 68, 2960–2966. [Google Scholar] [CrossRef] [PubMed]
- Bozal, N.; Montes, M.J.; Tudela, E.; Jiménez, F.; Guinea, J. Shewanella frigidimarina and Shewanella livingstonensis sp. nov. isolated from Antarctic coastal areas. Int. J. Syst. Evol. Microbiol. 2002, 52, 195–205. [Google Scholar] [CrossRef] [PubMed]
- Park, S.C.; Baik, K.S.; Kim, M.S.; Kim, D.; Seong, C.N. Shewanella marina sp. nov., isolated from seawater. Int. J. Syst. Evol. Microbiol. 2009, 59, 1888–1894. [Google Scholar] [CrossRef] [PubMed]
- Bae, S.S.; Jung, Y.-h.; Baek, K. Shewanella maritima sp. nov., a facultative anaerobic marine bacterium isolated from seawater, and emended description of Shewanella intestini. Int. J. Syst. Evol. Microbiol. 2020, 70, 1288–1293. [Google Scholar] [CrossRef]
- Makemson, J.C.; Fulayfil, N.R.; Landry, W.; Van Ert, L.M.; Wimpee, C.F.; Widder, E.A.; Case, J.F. Shewanella woodyi sp. nov., an exclusively respiratory luminous bacterium isolated from the Alboran Sea. Int. J. Syst. Bacteriol. 1997, 47, 1034–1039. [Google Scholar] [CrossRef]
- Park, H.Y.; Jeon, C.O. Shewanella aestuarii sp. nov., a marine bacterium isolated from a tidal flat. Int. J. Syst. Evol. Microbiol. 2013, 63, 4683–4690. [Google Scholar] [CrossRef]
- Baaziz, H.; Lemaire, O.N.; Jourlin-Castelli, C.; Iobbi-Nivol, C.; Méjean, V.; Alatou, R.; Fons, M. Draft Genome Sequence of Shewanella algidipiscicola H1, a Highly Chromate-Resistant Strain Isolated from Mediterranean Marine Sediments. Microbiol. Resour. Announc. 2018, 7, e00905-18. [Google Scholar] [CrossRef]
- Venkateswaran, K.; Dollhopf, M.E.; Aller, R.; Stackebrandt, E.; Nealson, K.H. Shewanella amazonensis sp. nov., a novel metal-reducing facultative anaerobe from Amazonian shelf muds. Int. J. Syst. Bacteriol. 1998, 48 Pt 3, 965–972. [Google Scholar] [CrossRef] [PubMed]
- Sucharita, K.; Sasikala, C.; Park, S.C.; Baik, K.S.; Seong, C.N.; Ramana, C.V. Shewanella chilikensis sp. nov., a moderately alkaliphilic gammaproteobacterium isolated from a lagoon. Int. J. Syst. Evol. Microbiol. 2009, 59, 3111–3115. [Google Scholar] [CrossRef]
- Yang, S.-H.; Lee, J.-H.; Ryu, J.-S.; Kato, C.; Kim, S.-J. Shewanella donghaensis sp. nov., a psychrophilic, piezosensitive bacterium producing high levels of polyunsaturated fatty acid, isolated from deep-sea sediments. Int. J. Syst. Evol. Microbiol. 2007, 57, 208–212. [Google Scholar] [CrossRef]
- Ivanova, E.P.; Sawabe, T.; Hayashi, K.; Gorshkova, N.M.; Zhukova, N.V.; Nedashkovskaya, O.I.; Mikhailov, V.V.; Nicolau, D.V.; Christen, R. Shewanella fidelis sp. nov., isolated from sediments and sea water. Int. J. Syst. Evol. Microbiol. 2003, 53, 577–582. [Google Scholar] [CrossRef] [PubMed]
- Verma, P.; Pandey, P.K.; Gupta, A.K.; Kim, H.J.; Baik, K.S.; Seong, C.N.; Patole, M.S.; Shouche, Y.S. Shewanella indica sp. nov., isolated from sediment of the Arabian Sea. Int. J. Syst. Evol. Microbiol. 2011, 61, 2058–2064. [Google Scholar] [CrossRef]
- Wang, M.-q.; Sun, L. Shewanella inventionis sp. nov., isolated from deep-sea sediment. Int. J. Syst. Evol. Microbiol. 2016, 66, 4947–4953. [Google Scholar] [CrossRef]
- Lee, M.H.; Yoon, J.H. Shewanella litorisediminis sp. nov., a gammaproteobacterium isolated from a tidal flat sediment. Antonie Van Leeuwenhoek 2012, 102, 591–599. [Google Scholar] [CrossRef]
- Liu, Y.; Shang, X.-X.; Yi, Z.-W.; Gu, L.; Zeng, R.-Y. Shewanella mangrovi sp. nov., an acetaldehyde-degrading bacterium isolated from mangrove sediment. Int. J. Syst. Evol. Microbiol. 2015, 65, 2630–2634. [Google Scholar] [CrossRef] [PubMed]
- Huang, J.; Sun, B.; Zhang, X. Electricity generation at high ionic strength in microbial fuel cell by a newly isolated Shewanella marisflavi EP1. Appl. Microbiol. Biotechnol. 2010, 85, 1141–1149. [Google Scholar] [CrossRef]
- Huang, J.; Ning, G.; Li, F.; Sheng, G.D. Biotransformation of 2,4-dinitrotoluene by obligate marine Shewanella marisflavi EP1 under anaerobic conditions. Bioresour. Technol. 2015, 180, 200–206. [Google Scholar] [CrossRef]
- Venkateswaran, K.; Moser, D.P.; Dollhopf, M.E.; Lies, D.P.; Saffarini, D.A.; MacGregor, B.J.; Ringelberg, D.B.; White, D.C.; Nishijima, M.; Sano, H.; et al. Polyphasic taxonomy of the genus Shewanella and description of Shewanella oneidensis sp. nov. Int. J. Syst. Evol. Microbiol. 1999, 49, 705–724. [Google Scholar] [CrossRef] [PubMed]
- Heidelberg, J.F.; Paulsen, I.T.; Nelson, K.E.; Gaidos, E.J.; Nelson, W.C.; Read, T.D.; Eisen, J.A.; Seshadri, R.; Ward, N.; Methe, B.; et al. Genome sequence of the dissimilatory metal ion–reducing bacterium Shewanella oneidensis. Nat. Biotechnol. 2002, 20, 1118–1123. [Google Scholar] [CrossRef] [PubMed]
- Xiao, X.; Wang, P.; Zeng, X.; Bartlett, D.H.; Wang, F. Shewanella psychrophila sp. nov. and Shewanella piezotolerans sp. nov., isolated from west Pacific deep-sea sediment. Int. J. Syst. Evol. Microbiol. 2007, 57, 60–65. [Google Scholar] [CrossRef] [PubMed]
- Wang, F.; Wang, J.; Jian, H.; Zhang, B.; Li, S.; Wang, F.; Zeng, X.; Gao, L.; Bartlett, D.H.; Yu, J.; et al. Environmental adaptation: Genomic analysis of the piezotolerant and psychrotolerant deep-sea iron reducing bacterium Shewanella piezotolerans WP3. PLoS ONE 2008, 3, e1937. [Google Scholar] [CrossRef]
- Xu, G.; Jian, H.; Xiao, X.; Wang, F. Complete genome sequence of Shewanella psychrophila WP2, a deep-sea bacterium isolated from west Pacific sediment. Mar. Genom. 2017, 35, 19–21. [Google Scholar] [CrossRef]
- Li, S.; Niu, Y.; Chen, H.; He, P. Complete genome sequence of an Arctic Ocean bacterium Shewanella sp. Arc9-LZ with capacity of synthesizing silver nanoparticles in darkness. Mar. Genom. 2021, 56, 100808. [Google Scholar] [CrossRef]
- Bujak, K.; Decewicz, P.; Rosinska, J.M.; Radlinska, M. Genome Study of a Novel Virulent Phage vB_SspS_KASIA and Mu-like Prophages of Shewanella sp. M16 Provides Insights into the Genetic Diversity of the Shewanella Virome. Int. J. Mol. Sci. 2021, 22, 11070. [Google Scholar] [CrossRef]
- Yang, Y.; Chen, J.; Qiu, D.; Zhou, J. Roles of UndA and MtrC of Shewanella putrefaciens W3-18-1 in iron reduction. BMC Microbiol. 2013, 13, 267. [Google Scholar] [CrossRef] [PubMed]
- Qiu, D.; Wei, H.; Tu, Q.; Yang, Y.; Xie, M.; Chen, J.; Pinkerton, M.H.; Liang, Y.; He, Z.; Zhou, J. Combined Genomics and Experimental Analyses of Respiratory Characteristics of Shewanella putrefaciens W3-18-1. Appl. Environ. Microbiol. 2013, 79, 5250–5257. [Google Scholar] [CrossRef]
- Bozal, N.; Montes, M.J.; Miñana-Galbis, D.; Manresa, A.; Mercadé, E. Shewanella vesiculosa sp. nov., a psychrotolerant bacterium isolated from an Antarctic coastal area. Int. J. Syst. Evol. Microbiol. 2009, 59, 336–340. [Google Scholar] [CrossRef]
- Nogi, Y.; Kato, C.; Horikoshi, K. Taxonomic studies of deep-sea barophilic Shewanella strains and description of Shewanella violacea sp. nov. Arch. Microbiol. 1998, 170, 331–338. [Google Scholar] [CrossRef]
- Chikuma, S.; Kasahara, R.; Kato, C.; Tamegai, H. Bacterial adaptation to high pressure: A respiratory system in the deep-sea bacterium Shewanella violacea DSS12. FEMS Microbiol. Lett. 2007, 267, 108–112. [Google Scholar] [CrossRef] [PubMed]
- Jensen, M.J.; Tebo, B.M.; Baumann, P.; Mandel, M.; Nealson, K.H. Characterization of Alteromonas hanedai (sp. nov.), a nonfermentative luminous species of marine origin. Curr. Microbiol. 1980, 3, 311–315. [Google Scholar] [CrossRef]
- Lee, J.-H.; Roh, Y.; Kim, K.-W.; Hur, H.-G. Organic Acid-Dependent Iron Mineral Formation by a Newly Isolated Iron-Reducing Bacterium, Shewanella sp. HN-41. Geomicrobiol. J. 2007, 24, 31–41. [Google Scholar] [CrossRef]
- Lee, J.H.; Kim, M.G.; Yoo, B.; Myung, N.V.; Maeng, J.; Lee, T.; Dohnalkova, A.C.; Fredrickson, J.K.; Sadowsky, M.J.; Hur, H.G. Biogenic formation of photoactive arsenic-sulfide nanotubes by Shewanella sp. strain HN-41. Proc. Natl. Acad. Sci. USA 2007, 104, 20410–20415. [Google Scholar] [CrossRef] [PubMed]
- Zhao, J.-S.; Manno, D.; Leggiadro, C.; Leggiadro, C.; O’Neil, C.; O’Neil, C.; Hawari, J. Shewanella halifaxensis sp. nov., a novel obligately respiratory and denitrifying psychrophile. Int. J. Syst. Evol. Microbiol. 2006, 56, 205–212. [Google Scholar] [CrossRef]
- Li, Z.; Song, F.; Chen, M. Complete Genome Sequence of Shewanella sp. Strain Lzh-2, an Algicidal Bacterial Strain Isolated from Lake Taihu, People’s Republic of China. Microbiol. Resour. Announc. 2021, 10, e00339-21. [Google Scholar] [CrossRef]
- Li, Z.; Lin, S.; Liu, X.; Tan, J.; Pan, J.; Yang, H. A freshwater bacterial strain, Shewanella sp. Lzh-2, isolated from Lake Taihu and its two algicidal active substances, hexahydropyrrolo[1,2-a]pyrazine-1,4-dione and 2, 3-indolinedione. Appl. Microbiol. Biotechnol. 2014, 98, 4737–4748. [Google Scholar] [CrossRef]
- Boles, B.W.; Murdoch, R.W.; Ohlsson, I.; Mikucki, J.A. Draft Genome Sequence of Shewanella sp. Strain BF02_Schw, Isolated from Blood Falls, a Feature Where Subglacial Brine Discharges to the Surface of Taylor Glacier, Antarctica. Microbiol. Resour. Announc. 2021, 10, e01141-20. [Google Scholar] [CrossRef]
- Sravan Kumar, R.; Sasi Jyothsna, T.S.; Sasikala, C.; Seong, C.N.; Lim, C.H.; Park, S.C.; Ramana, C.V. Shewanella fodinae sp. nov., isolated from a coal mine and from a marine lagoon. Int. J. Syst. Evol. Microbiol. 2010, 60, 1649–1654. [Google Scholar] [CrossRef]
- Saltikov, C.W.; Cifuentes, A.; Venkateswaran, K.; Newman, D.K. The ars detoxification system is advantageous but not required for As(V) respiration by the genetically tractable Shewanella species strain ANA-3. Appl. Environ. Microbiol. 2003, 69, 2800–2809. [Google Scholar] [CrossRef] [PubMed]
- Saltikov, C.W.; Wildman, R.A., Jr.; Newman, D.K. Expression dynamics of arsenic respiration and detoxification in Shewanella sp. strain ANA-3. J. Bacteriol. 2005, 187, 7390–7396. [Google Scholar] [CrossRef]
- Zargar, K.; Saltikov, C.W. Lysine-91 of the tetraheme c-type cytochrome CymA is essential for quinone interaction and arsenate respiration in Shewanella sp. strain ANA-3. Arch. Microbiol. 2009, 191, 797–806. [Google Scholar] [CrossRef] [PubMed]
- Qasim, M.S.; Lampi, M.; Heinonen, M.-M.K.; Garrido-Zabala, B.; Bamford, D.H.; Käkelä, R.; Roine, E.; Sarin, L.P. Cold-Active Shewanella glacialimarina TZS-4T nov. Features a Temperature-Dependent Fatty Acid Profile and Putative Sialic Acid Metabolism. Front. Microbiol. 2021, 12, 737641. [Google Scholar] [CrossRef]
- Cao, W.-R.; Li, X.; Sun, Y.-Y.; Jiang, M.-Y.; Xu, X.-D.; Li, Y.-J. Shewanella nanhaiensis sp. nov., a marine bacterium isolated from sediment of South China Sea, and emended descriptions of Shewanella woodyi, Shewanella hanedai and Shewanella canadensis. Int. J. Syst. Evol. Microbiol. 2021, 71, 005152. [Google Scholar] [CrossRef]
- Yu, N.Y.; Wagner, J.R.; Laird, M.R.; Melli, G.; Rey, S.; Lo, R.; Dao, P.; Sahinalp, S.C.; Ester, M.; Foster, L.J.; et al. PSORTb 3.0: Improved protein subcellular localization prediction with refined localization subcategories and predictive capabilities for all prokaryotes. Bioinformatics 2010, 26, 1608–1615. [Google Scholar] [CrossRef]
- Ishii, S.I.; Suzuki, S.; Norden-Krichmar, T.M.; Tenney, A.; Chain, P.S.G.; Scholz, M.B.; Nealson, K.H.; Bretschger, O. A novel metatranscriptomic approach to identify gene expression dynamics during extracellular electron transfer. Nat. Commun. 2013, 4, 1601. [Google Scholar] [CrossRef] [PubMed]
- Arndt, D.; Grant, J.R.; Marcu, A.; Sajed, T.; Pon, A.; Liang, Y.; Wishart, D.S. PHASTER: A better, faster version of the PHAST phage search tool. Nucleic Acids Res. 2016, 44, W16–W21. [Google Scholar] [CrossRef]
- Makarova, K.S.; Wolf, Y.I.; Iranzo, J.; Shmakov, S.A.; Alkhnbashi, O.S.; Brouns, S.J.J.; Charpentier, E.; Cheng, D.; Haft, D.H.; Horvath, P.; et al. Evolutionary classification of CRISPR–Cas systems: A burst of class 2 and derived variants. Nat. Rev. Microbiol. 2020, 18, 67–83. [Google Scholar] [CrossRef] [PubMed]
- Siguier, P.; Perochon, J.; Lestrade, L.; Mahillon, J.; Chandler, M. ISfinder: The reference centre for bacterial insertion sequences. Nucleic Acids Res. 2006, 34, D32–D36. [Google Scholar] [CrossRef]
- Néron, B.; Littner, E.; Haudiquet, M.; Perrin, A.; Cury, J.; Rocha, E.P.C. IntegronFinder 2.0: Identification and Analysis of Integrons across Bacteria, with a Focus on Antibiotic Resistance in Klebsiella. Microorganisms 2022, 10, 700. [Google Scholar] [CrossRef] [PubMed]
- Abby, S.S.; Néron, B.; Ménager, H.; Touchon, M.; Rocha, E.P.C. MacSyFinder: A Program to Mine Genomes for Molecular Systems with an Application to CRISPR-Cas Systems. PLoS ONE 2014, 9, e110726. [Google Scholar] [CrossRef] [PubMed]
- Cury, J.; Touchon, M.; Rocha, E.P.C. Integrative and conjugative elements and their hosts: Composition, distribution and organization. Nucleic Acids Res. 2017, 45, 8943–8956. [Google Scholar] [CrossRef] [PubMed]
- Bastian, M.; Heymann, S.; Jacomy, M. Gephi: An Open Source Software for Exploring and Manipulating Networks. In Proceedings of the International AAAI Conference on Web and Social Media, San Jose, CA, USA, 17–20 May 2009; Volume 3, pp. 361–362. [Google Scholar] [CrossRef]
- Fruchterman, T.M.J.; Reingold, E.M. Graph drawing by force-directed placement. Softw. Pract. Exp. 1991, 21, 1129–1164. [Google Scholar] [CrossRef]
- Hu, Y. Efficient and High Quality Force-Directed Graph Drawing. Math. J. 2005, 10, 37–71. [Google Scholar]
- Blondel, V.D.; Guillaume, J.-L.; Lambiotte, R.; Lefebvre, E. Fast unfolding of communities in large networks. J. Stat. Mech. Theory Exp. 2008, 2008, P10008. [Google Scholar] [CrossRef]
- Larsson, A. AliView: A fast and lightweight alignment viewer and editor for large datasets. Bioinformatics 2014, 30, 3276–3278. [Google Scholar] [CrossRef]
- Ronquist, F.; Teslenko, M.; van der Mark, P.; Ayres, D.L.; Darling, A.; Höhna, S.; Larget, B.; Liu, L.; Suchard, M.A.; Huelsenbeck, J.P. MrBayes 3.2: Efficient Bayesian phylogenetic inference and model choice across a large model space. Syst. Biol. 2012, 61, 539–542. [Google Scholar] [CrossRef]
- Garcia, P.S.; Jauffrit, F.; Grangeasse, C.; Brochier-Armanet, C. GeneSpy, a user-friendly and flexible genomic context visualizer. Bioinformatics 2019, 35, 329–331. [Google Scholar] [CrossRef]
- Chang, D.; Keinan, A. Principal Component Analysis Characterizes Shared Pathogenetics from Genome-Wide Association Studies. PLOS Comput. Biol. 2014, 10, e1003820. [Google Scholar] [CrossRef]
- Lovley, D.R.; Ueki, T.; Zhang, T.; Malvankar, N.S.; Shrestha, P.M.; Flanagan, K.A.; Aklujkar, M.; Butler, J.E.; Giloteaux, L.; Rotaru, A.E.; et al. Geobacter: The microbe electric’s physiology, ecology, and practical applications. Adv. Microb. Physiol. 2011, 59, 1–100. [Google Scholar] [CrossRef] [PubMed]
- Anderson, R.T.; Vrionis, H.A.; Ortiz-Bernad, I.; Resch, C.T.; Long, P.E.; Dayvault, R.; Karp, K.; Marutzky, S.; Metzler, D.R.; Peacock, A.; et al. Stimulating the in situ activity of Geobacter species to remove uranium from the groundwater of a uranium-contaminated aquifer. Appl. Environ. Microbiol. 2003, 69, 5884–5891. [Google Scholar] [CrossRef]
- Coates, J.D.; Phillips, E.J.; Lonergan, D.J.; Jenter, H.; Lovley, D.R. Isolation of Geobacter species from diverse sedimentary environments. Appl. Environ. Microbiol. 1996, 62, 1531–1536. [Google Scholar] [CrossRef] [PubMed]
- Snoeyenbos-West, O.L.; Nevin, K.P.; Anderson, R.T.; Lovley, D.R. Enrichment of Geobacter Species in Response to Stimulation of Fe(III) Reduction in Sandy Aquifer Sediments. Microb. Ecol. 2000, 39, 153–167. [Google Scholar] [CrossRef]
- Li, T.; Zhou, Q. The key role of Geobacter in regulating emissions and biogeochemical cycling of soil-derived greenhouse gases. Environ. Pollut. 2020, 266, 115135. [Google Scholar] [CrossRef] [PubMed]
- Lemaire, O.N.; Méjean, V.; Iobbi-Nivol, C. The Shewanella genus: Ubiquitous organisms sustaining and preserving aquatic ecosystems. FEMS Microbiol. Rev. 2020, 44, 155–170. [Google Scholar] [CrossRef]
- Lightfield, J.; Fram, N.R.; Ely, B. Across bacterial phyla, distantly-related genomes with similar genomic GC content have similar patterns of amino acid usage. PLoS ONE 2011, 6, e17677. [Google Scholar] [CrossRef]
- Bragg, J.G.; Hyder, C.L. Nitrogen versus carbon use in prokaryotic genomes and proteomes. Proc. Biol. Sci. 2004, 271 (Suppl. S5), S374–S377. [Google Scholar] [CrossRef]
- Dugdale, R.C. Nutrient limitation in the Sea: Dynamics, Identification and Significance. Limnol. Oceanogr. 1967, 12, 685–695. [Google Scholar] [CrossRef]
- Ely, B. Genomic GC content drifts downward in most bacterial genomes. PLoS ONE 2021, 16, e0244163. [Google Scholar] [CrossRef]
- Kotloski, N.J.; Gralnick, J.A. Flavin electron shuttles dominate extracellular electron transfer by Shewanella oneidensis. mBio 2013, 4, e00553-12. [Google Scholar] [CrossRef] [PubMed]
- Zou, L.; Zhu, F.; Long, Z.-E.; Huang, Y. Bacterial extracellular electron transfer: A powerful route to the green biosynthesis of inorganic nanomaterials for multifunctional applications. J. Nanobiotechnol. 2021, 19, 120. [Google Scholar] [CrossRef] [PubMed]
- Liu, D.; Dong, H.; Bishop, M.E.; Zhang, J.; Wang, H.; Xie, S.; Wang, S.; Huang, L.; Eberl, D.D. Microbial reduction of structural iron in interstratified illite-smectite minerals by a sulfate-reducing bacterium. Geobiology 2012, 10, 150–162. [Google Scholar] [CrossRef]
- Simonte, F.; Sturm, G.; Gescher, J.; Sturm-Richter, K. Extracellular Electron Transfer and Biosensors. In Bioelectrosynthesis; Harnisch, F., Holtmann, D., Eds.; Springer International Publishing: Cham, Switzerland, 2019; pp. 15–38. [Google Scholar]
- Shi, L.; Squier, T.C.; Zachara, J.M.; Fredrickson, J.K. Respiration of metal (hydr)oxides by Shewanella and Geobacter: A key role for multihaem c-type cytochromes. Mol. Microbiol. 2007, 65, 12–20. [Google Scholar] [CrossRef] [PubMed]
- Salgueiro, C.A.; Morgado, L.; Silva, M.A.; Ferreira, M.R.; Fernandes, T.M.; Portela, P.C. From iron to bacterial electroconductive filaments: Exploring cytochrome diversity using Geobacter bacteria. Coord. Chem. Rev. 2022, 452, 214284. [Google Scholar] [CrossRef]
- Holmes, D.E.; Zhou, J.; Smith, J.A.; Wang, C.; Liu, X.; Lovley, D.R. Different outer membrane c-type cytochromes are involved in direct interspecies electron transfer to Geobacter or Methanosarcina species. mLife 2022, 1, 272–286. [Google Scholar] [CrossRef]
- Lovley, D.R. Syntrophy Goes Electric: Direct Interspecies Electron Transfer. Annu. Rev. Microbiol. 2017, 71, 643–664. [Google Scholar] [CrossRef]
- Pereira, I.A.C.; Ramos, A.R.; Grein, F.; Marques, M.C.; Da Silva, S.M.; Venceslau, S.S. A Comparative Genomic Analysis of Energy Metabolism in Sulfate Reducing Bacteria and Archaea. Front. Microbiol. 2011, 2, 69. [Google Scholar] [CrossRef]
- Morgado, L.; Fernandes, A.P.; Dantas, J.M.; Silva, M.A.; Salgueiro, C.A. On the road to improve the bioremediation and electricity-harvesting skills of Geobacter sulfurreducens: Functional and structural characterization of multihaem cytochromes. Biochem. Soc. Trans. 2012, 40, 1295–1301. [Google Scholar] [CrossRef]
- Levar, C.E.; Chan, C.H.; Mehta-Kolte, M.G.; Bond, D.R. An inner membrane cytochrome required only for reduction of high redox potential extracellular electron acceptors. mBio 2014, 5, e02034. [Google Scholar] [CrossRef]
- Zacharoff, L.; Chan, C.H.; Bond, D.R. Reduction of low potential electron acceptors requires the CbcL inner membrane cytochrome of Geobacter sulfurreducens. Bioelectrochemistry 2016, 107, 7–13. [Google Scholar] [CrossRef]
- Aklujkar, M.; Coppi, M.V.; Leang, C.; Kim, B.C.; Chavan, M.A.; Perpetua, L.A.; Giloteaux, L.; Liu, A.; Holmes, D.E. Proteins involved in electron transfer to Fe(III) and Mn(IV) oxides by Geobacter sulfurreducens and Geobacter uraniireducens. Microbiology 2013, 159, 515–535. [Google Scholar] [CrossRef]
- Peng, L.; Zhang, X.T.; Yin, J.; Xu, S.Y.; Zhang, Y.; Xie, D.T.; Li, Z.L. Geobacter sulfurreducens adapts to low electrode potential for extracellular electron transfer. Electrochim. Acta 2016, 191, 743–749. [Google Scholar] [CrossRef]
- Butler, J.E.; Young, N.D.; Lovley, D.R. Evolution of electron transfer out of the cell: Comparative genomics of six Geobacter genomes. BMC Genom. 2010, 11, 40. [Google Scholar] [CrossRef]
- Lloyd, J.R.; Leang, C.; Hodges Myerson, A.L.; Coppi, M.V.; Cuifo, S.; Methe, B.; Sandler, S.J.; Lovley, D.R. Biochemical and genetic characterization of PpcA, a periplasmic c-type cytochrome in Geobacter sulfurreducens. Biochem. J. 2003, 369, 153–161. [Google Scholar] [CrossRef]
- Butler, J.E.; Kaufmann, F.; Coppi, M.V.; Núñez, C.; Lovley, D.R. MacA, a diheme c-type cytochrome involved in Fe(III) reduction by Geobacter sulfurreducens. J. Bacteriol. 2004, 186, 4042–4045. [Google Scholar] [CrossRef]
- Liu, Y.; Wang, Z.; Liu, J.; Levar, C.; Edwards, M.J.; Babauta, J.T.; Kennedy, D.W.; Shi, Z.; Beyenal, H.; Bond, D.R.; et al. A trans-outer membrane porin-cytochrome protein complex for extracellular electron transfer by Geobacter sulfurreducens PCA. Environ. Microbiol. Rep. 2014, 6, 776–785. [Google Scholar] [CrossRef]
- Jiménez Otero, F.; Chan, C.H.; Bond, D.R. Identification of Different Putative Outer Membrane Electron Conduits Necessary for Fe(III) Citrate, Fe(III) Oxide, Mn(IV) Oxide, or Electrode Reduction by Geobacter sulfurreducens. J. Bacteriol. 2018, 200, e00347-18. [Google Scholar] [CrossRef]
- Mehta, T.; Coppi, M.V.; Childers, S.E.; Lovley, D.R. Outer Membrane c-Type Cytochromes Required for Fe(III) and Mn(IV) Oxide Reduction in Geobacter sulfurreducens. Appl. Environ. Microbiol. 2005, 71, 8634–8641. [Google Scholar] [CrossRef]
- Leang, C.; Qian, X.; Mester, T.; Lovley, D.R. Alignment of the c-Type Cytochrome OmcS along Pili of Geobacter sulfurreducens. Appl. Environ. Microbiol. 2010, 76, 4080–4084. [Google Scholar] [CrossRef]
- Wang, F.; Gu, Y.; O’Brien, J.P.; Yi, S.M.; Yalcin, S.E.; Srikanth, V.; Shen, C.; Vu, D.; Ing, N.L.; Hochbaum, A.I.; et al. Structure of Microbial Nanowires Reveals Stacked Hemes that Transport Electrons over Micrometers. Cell 2019, 177, 361–369.e10. [Google Scholar] [CrossRef]
- Nevin, K.P.; Kim, B.C.; Glaven, R.H.; Johnson, J.P.; Woodard, T.L.; Methé, B.A.; Didonato, R.J.; Covalla, S.F.; Franks, A.E.; Liu, A.; et al. Anode biofilm transcriptomics reveals outer surface components essential for high density current production in Geobacter sulfurreducens fuel cells. PLoS ONE 2009, 4, e5628. [Google Scholar] [CrossRef]
- Holmes, D.E.; Chaudhuri, S.K.; Nevin, K.P.; Mehta, T.; Methé, B.A.; Liu, A.; Ward, J.E.; Woodard, T.L.; Webster, J.; Lovley, D.R. Microarray and genetic analysis of electron transfer to electrodes in Geobacter sulfurreducens. Environ. Microbiol. 2006, 8, 1805–1815. [Google Scholar] [CrossRef]
- Zacharoff, L.A.; Morrone, D.J.; Bond, D.R. Geobacter sulfurreducens Extracellular Multiheme Cytochrome PgcA Facilitates Respiration to Fe(III) Oxides But Not Electrodes. Front. Microbiol. 2017, 8, 2481. [Google Scholar] [CrossRef]
- Afkar, E.; Reguera, G.; Schiffer, M.; Lovley, D.R. A novel Geobacteraceae-specific outer membrane protein J (OmpJ) is essential for electron transport to Fe(III) and Mn(IV) oxides in Geobacter sulfurreducens. BMC Microbiol. 2005, 5, 41. [Google Scholar] [CrossRef]
- Rollefson, J.B.; Stephen, C.S.; Tien, M.; Bond, D.R. Identification of an extracellular polysaccharide network essential for cytochrome anchoring and biofilm formation in Geobacter sulfurreducens. J. Bacteriol. 2011, 193, 1023–1033. [Google Scholar] [CrossRef]
- Huang, L.; Liu, X.; Ye, Y.; Chen, M.; Zhou, S. Evidence for the coexistence of direct and riboflavin-mediated interspecies electron transfer in Geobacter co-culture. Environ. Microbiol. 2020, 22, 243–254. [Google Scholar] [CrossRef]
- Myers, C.R.; Myers, J.M. Cloning and sequence of cymA, a gene encoding a tetraheme cytochrome c required for reduction of iron(III), fumarate, and nitrate by Shewanella putrefaciens MR-1. J. Bacteriol. 1997, 179, 1143–1152. [Google Scholar] [CrossRef]
- Shi, L.; Rosso, K.M.; Clarke, T.A.; Richardson, D.J.; Zachara, J.M.; Fredrickson, J.K. Molecular Underpinnings of Fe(III) Oxide Reduction by Shewanella oneidensis MR-1. Front. Microbiol. 2012, 3, 50. [Google Scholar] [CrossRef]
- Sturm, G.; Richter, K.; Doetsch, A.; Heide, H.; Louro, R.O.; Gescher, J. A dynamic periplasmic electron transfer network enables respiratory flexibility beyond a thermodynamic regulatory regime. ISME J. 2015, 9, 1802–1811. [Google Scholar] [CrossRef]
- Beliaev, A.S.; Saffarini, D.A. Shewanella putrefaciens mtrB encodes an outer membrane protein required for Fe(III) and Mn(IV) reduction. J. Bacteriol. 1998, 180, 6292–6297. [Google Scholar] [CrossRef]
- Beliaev, A.S.; Saffarini, D.A.; McLaughlin, J.L.; Hunnicutt, D. MtrC, an outer membrane decahaem c cytochrome required for metal reduction in Shewanella putrefaciens MR-1. Mol. Microbiol. 2001, 39, 722–730. [Google Scholar] [CrossRef]
- Ross, D.E.; Ruebush, S.S.; Brantley, S.L.; Hartshorne, R.S.; Clarke, T.A.; Richardson, D.J.; Tien, M. Characterization of protein-protein interactions involved in iron reduction by Shewanella oneidensis MR-1. Appl. Environ. Microbiol. 2007, 73, 5797–5808. [Google Scholar] [CrossRef]
- Pirbadian, S.; Barchinger, S.E.; Leung, K.M.; Byun, H.S.; Jangir, Y.; Bouhenni, R.A.; Reed, S.B.; Romine, M.F.; Saffarini, D.A.; Shi, L.; et al. Shewanella oneidensis MR-1 nanowires are outer membrane and periplasmic extensions of the extracellular electron transport components. Proc. Natl. Acad. Sci. USA 2014, 111, 12883–12888. [Google Scholar] [CrossRef]
- Marsili, E.; Baron, D.B.; Shikhare, I.D.; Coursolle, D.; Gralnick, J.A.; Bond, D.R. Shewanella secretes flavins that mediate extracellular electron transfer. Proc. Natl. Acad. Sci. USA 2008, 105, 3968–3973. [Google Scholar] [CrossRef]
- Edwards, M.J.; White, G.F.; Norman, M.; Tome-Fernandez, A.; Ainsworth, E.; Shi, L.; Fredrickson, J.K.; Zachara, J.M.; Butt, J.N.; Richardson, D.J.; et al. Redox Linked Flavin Sites in Extracellular Decaheme Proteins Involved in Microbe-Mineral Electron Transfer. Sci. Rep. 2015, 5, 11677. [Google Scholar] [CrossRef]
- Min, D.; Cheng, L.; Zhang, F.; Huang, X.-N.; Li, D.-B.; Liu, D.-F.; Lau, T.-C.; Mu, Y.; Yu, H.-Q. Enhancing Extracellular Electron Transfer of Shewanella oneidensis MR-1 through Coupling Improved Flavin Synthesis and Metal-Reducing Conduit for Pollutant Degradation. Environ. Sci. Technol. 2017, 51, 5082–5089. [Google Scholar] [CrossRef]
- Li, H.; Xu, D.; Li, Y.; Feng, H.; Liu, Z.; Li, X.; Gu, T.; Yang, K. Extracellular Electron Transfer Is a Bottleneck in the Microbiologically Influenced Corrosion of C1018 Carbon Steel by the Biofilm of Sulfate-Reducing Bacterium Desulfovibrio vulgaris. PLoS ONE 2015, 10, e0136183. [Google Scholar] [CrossRef]
- Zhang, P.; Xu, D.; Li, Y.; Yang, K.; Gu, T. Electron mediators accelerate the microbiologically influenced corrosion of 304 stainless steel by the Desulfovibrio vulgaris biofilm. Bioelectrochemistry 2015, 101, 14–21. [Google Scholar] [CrossRef]
- Carr, V.R.; Shkoporov, A.; Hill, C.; Mullany, P.; Moyes, D.L. Probing the Mobilome: Discoveries in the Dynamic Microbiome. Trends Microbiol. 2021, 29, 158–170. [Google Scholar] [CrossRef]
- Attrill, E.L.; Łapińska, U.; Westra, E.R.; Harding, S.V.; Pagliara, S. Slow growing bacteria survive bacteriophage in isolation. ISME Commun. 2023, 3, 95. [Google Scholar] [CrossRef]
- Suttle, C.A. Marine viruses--major players in the global ecosystem. Nat. Rev. Microbiol. 2007, 5, 801–812. [Google Scholar] [CrossRef] [PubMed]
- Braga, L.P.P.; Spor, A.; Kot, W.; Breuil, M.-C.; Hansen, L.H.; Setubal, J.C.; Philippot, L. Impact of phages on soil bacterial communities and nitrogen availability under different assembly scenarios. Microbiome 2020, 8, 52. [Google Scholar] [CrossRef]
- Barrangou, R.; Fremaux, C.; Deveau, H.; Richards, M.; Boyaval, P.; Moineau, S.; Romero, D.A.; Horvath, P. CRISPR provides acquired resistance against viruses in prokaryotes. Science 2007, 315, 1709–1712. [Google Scholar] [CrossRef] [PubMed]
- Lopez-Sanchez, M.-J.; Sauvage, E.; Da Cunha, V.; Clermont, D.; Ratsima Hariniaina, E.; Gonzalez-Zorn, B.; Poyart, C.; Rosinski-Chupin, I.; Glaser, P. The highly dynamic CRISPR1 system of Streptococcus agalactiae controls the diversity of its mobilome. Mol. Microbiol. 2012, 85, 1057–1071. [Google Scholar] [CrossRef]
- Samson, J.E.; Magadan, A.H.; Moineau, S. The CRISPR-Cas Immune System and Genetic Transfers: Reaching an Equilibrium. Microbiol. Spectr. 2015, 3, 209–218. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Zheng, K.; Wang, M.; Ma, K.; Ren, L.; Guo, R.; Ma, L.; Zhang, H.; Liu, Y.; Xiong, Y.; et al. Shewanella phage encoding a putative anti-CRISPR-like gene represents a novel potential viral family. Microbiol. Spectr. 2024, 12, e0336723. [Google Scholar] [CrossRef]
- Pawluk, A.; Staals, R.H.; Taylor, C.; Watson, B.N.; Saha, S.; Fineran, P.C.; Maxwell, K.L.; Davidson, A.R. Inactivation of CRISPR-Cas systems by anti-CRISPR proteins in diverse bacterial species. Nat. Microbiol. 2016, 1, 16085. [Google Scholar] [CrossRef]
- Hwang, S.; Maxwell, K.L. Meet the Anti-CRISPRs: Widespread Protein Inhibitors of CRISPR-Cas Systems. CRISPR J. 2019, 2, 23–30. [Google Scholar] [CrossRef]
- Cury, J.; Jové, T.; Touchon, M.; Néron, B.; Rocha, E.P. Identification and analysis of integrons and cassette arrays in bacterial genomes. Nucleic Acids Res. 2016, 44, 4539–4550. [Google Scholar] [CrossRef]
- Hall, R.M. Integrons and gene cassettes: Hotspots of diversity in bacterial genomes. Ann. N. Y. Acad. Sci. 2012, 1267, 71–78. [Google Scholar] [CrossRef] [PubMed]
- Berg, D.E.; Howe, M.M. (Eds.) Mobile DNA; American Society for Microbiology Publications: Washington, DC, USA, 1989. [Google Scholar]
- Siguier, P.; Gourbeyre, E.; Varani, A.; Ton-Hoang, B.; Chandler, M. Everyman’s Guide to Bacterial Insertion Sequences. Microbiol. Spectr. 2015, 3, MDNA3-0030-2014. [Google Scholar] [CrossRef]
- Pradel, N.; Ji, B.; Gimenez, G.; Talla, E.; Lenoble, P.; Garel, M.; Tamburini, C.; Patrick, F.; Lebrun, R.; Bertin, P.; et al. The First Genomic and Proteomic Characterization of a Deep-Sea Sulfate Reducer: Insights into the Piezophilic Lifestyle of Desulfovibrio piezophilus. PLoS ONE 2013, 8, e55130. [Google Scholar] [CrossRef] [PubMed]
- Heidelberg, J.F.; Seshadri, R.; Haveman, S.A.; Hemme, C.L.; Paulsen, I.T.; Kolonay, J.F.; Eisen, J.A.; Ward, N.; Methe, B.; Brinkac, L.M.; et al. The genome sequence of the anaerobic, sulfate-reducing bacterium Desulfovibrio vulgaris Hildenborough. Nat. Biotechnol. 2004, 22, 554–559. [Google Scholar] [CrossRef] [PubMed]
- Siguier, P.; Gourbeyre, E.; Chandler, M. Bacterial insertion sequences: Their genomic impact and diversity. FEMS Microbiol. Rev. 2014, 38, 865–891. [Google Scholar] [CrossRef] [PubMed]
- Ding, Y.H.; Hixson, K.K.; Aklujkar, M.A.; Lipton, M.S.; Smith, R.D.; Lovley, D.R.; Mester, T. Proteome of Geobacter sulfurreducens grown with Fe(III) oxide or Fe(III) citrate as the electron acceptor. Biochim. Biophys. Acta 2008, 1784, 1935–1941. [Google Scholar] [CrossRef]
- Ferreira, M.R.; Salgueiro, C.A. Biomolecular Interaction Studies Between Cytochrome PpcA From Geobacter sulfurreducens and the Electron Acceptor Ferric Nitrilotriacetate (Fe-NTA). Front. Microbiol. 2018, 9, 2741. [Google Scholar] [CrossRef]
- Simon, J.; Hederstedt, L. Composition and function of cytochrome c biogenesis System II. FEBS J. 2011, 278, 4179–4188. [Google Scholar] [CrossRef]
- Lee, P.T.; Hsu, A.Y.; Ha, H.T.; Clarke, C.F. A C-methyltransferase involved in both ubiquinone and menaquinone biosynthesis: Isolation and identification of the Escherichia coli ubiE gene. J. Bacteriol. 1997, 179, 1748–1754. [Google Scholar] [CrossRef]
- Franza, T.; Gaudu, P. Quinones: More than electron shuttles. Res. Microbiol. 2022, 173, 103953. [Google Scholar] [CrossRef]
- Pokkuluri, P.R.; Londer, Y.Y.; Yang, X.; Duke, N.E.; Erickson, J.; Orshonsky, V.; Johnson, G.; Schiffer, M. Structural characterization of a family of cytochromes c(7) involved in Fe(III) respiration by Geobacter sulfurreducens. Biochim. Biophys. Acta 2010, 1797, 222–232. [Google Scholar] [CrossRef]
- Morgado, L.; Bruix, M.; Pessanha, M.; Londer, Y.Y.; Salgueiro, C.A. Thermodynamic characterization of a triheme cytochrome family from Geobacter sulfurreducens reveals mechanistic and functional diversity. Biophys. J. 2010, 99, 293–301. [Google Scholar] [CrossRef] [PubMed]
- Hannigan, R.; Dorval, E.; Jones, C. The rare earth element chemistry of estuarine surface sediments in the Chesapeake Bay. Chem. Geol. 2010, 272, 20–30. [Google Scholar] [CrossRef]
- Gilmour, C.C.; Elias, D.A.; Kucken, A.M.; Brown, S.D.; Palumbo, A.V.; Schadt, C.W.; Wall, J.D. Sulfate-reducing bacterium Desulfovibrio desulfuricans ND132 as a model for understanding bacterial mercury methylation. Appl. Environ. Microbiol. 2011, 77, 3938–3951. [Google Scholar] [CrossRef]
- Zhao, J.S.; Manno, D.; Beaulieu, C.; Paquet, L.; Hawari, J. Shewanella sediminis sp. nov., a novel Na+-requiring and hexahydro-1,3,5-trinitro-1,3,5-triazine-degrading bacterium from marine sediment. Int. J. Syst. Evol. Microbiol. 2005, 55, 1511–1520. [Google Scholar] [CrossRef]
- Zhao, J.-S.; Manno, D.; Thiboutot, S.; Ampleman, G.; Hawari, J. Shewanella canadensis sp. nov. and Shewanella atlantica sp. nov., manganese dioxide- and hexahydro-1,3,5-trinitro-1,3,5-triazine-reducing, psychrophilic marine bacteria. Int. J. Syst. Evol. Microbiol. 2007, 57, 2155–2162. [Google Scholar] [CrossRef]
- Ravenhall, M.; Škunca, N.; Lassalle, F.; Dessimoz, C. Inferring horizontal gene transfer. PLoS Comput. Biol. 2015, 11, e1004095. [Google Scholar] [CrossRef] [PubMed]
- Cuecas, A.; Kanoksilapatham, W.; Gonzalez, J.M. Evidence of horizontal gene transfer by transposase gene analyses in Fervidobacterium species. PLoS ONE 2017, 12, e0173961. [Google Scholar] [CrossRef]
- Postgate, J.R.; Campbell, L.L. Classification of Desulfovibrio species, the nonsporulating sulfate-reducing bacteria. Bacteriol. Rev. 1966, 30, 732–738. [Google Scholar] [CrossRef]
- Schwalb, C.; Chapman, S.K.; Reid, G.A. The tetraheme cytochrome CymA is required for anaerobic respiration with dimethyl sulfoxide and nitrite in Shewanella oneidensis. Biochemistry 2003, 42, 9491–9497. [Google Scholar] [CrossRef]
- Murphy, J.N.; Saltikov, C.W. The cymA gene, encoding a tetraheme c-type cytochrome, is required for arsenate respiration in Shewanella species. J. Bacteriol. 2007, 189, 2283–2290. [Google Scholar] [CrossRef] [PubMed]
- Fu, H.; Jin, M.; Ju, L.; Mao, Y.; Gao, H. Evidence for function overlapping of CymA and the cytochrome 1 complex in the hewanella oneidensis nitrate and nitrite respiration. Environ. Microbiol. 2014, 16, 3181–3195. [Google Scholar] [CrossRef] [PubMed]
- Dulay, H.; Tabares, M.; Kashefi, K.; Reguera, G. Cobalt Resistance via Detoxification and Mineralization in the Iron-Reducing Bacterium Geobacter sulfurreducens. Front. Microbiol. 2020, 11, 600463. [Google Scholar] [CrossRef] [PubMed]
- Mollaei, M.; Timmers, P.H.A.; Suarez-Diez, M.; Boeren, S.; van Gelder, A.H.; Stams, A.J.M.; Plugge, C.M. Comparative proteomics of Geobacter sulfurreducens PCA(T) in response to acetate, formate and/or hydrogen as electron donor. Environ. Microbiol. 2021, 23, 299–315. [Google Scholar] [CrossRef]
- Cordova, C.D.; Schicklberger, M.F.; Yu, Y.; Spormann, A.M. Partial functional replacement of CymA by SirCD in Shewanella oneidensis MR-1. J. Bacteriol. 2011, 193, 2312–2321. [Google Scholar] [CrossRef]
- Gherardini, P.F.; Wass, M.N.; Helmer-Citterich, M.; Sternberg, M.J. Convergent evolution of enzyme active sites is not a rare phenomenon. J. Mol. Biol. 2007, 372, 817–845. [Google Scholar] [CrossRef]
- Zhou, W.; Nakhleh, L. Convergent evolution of modularity in metabolic networks through different community structures. BMC Evol. Biol. 2012, 12, 181. [Google Scholar] [CrossRef]
- Aono, E.; Baba, T.; Ara, T.; Nishi, T.; Nakamichi, T.; Inamoto, E.; Toyonaga, H.; Hasegawa, M.; Takai, Y.; Okumura, Y.; et al. Complete genome sequence and comparative analysis of Shewanella violacea, a psychrophilic and piezophilic bacterium from deep sea floor sediments. Mol. BioSyst. 2010, 6, 1216–1226. [Google Scholar] [CrossRef]
- Brettar, I.; Christen, R.; Höfle, M.G. Shewanella denitrificans sp. nov., a vigorously denitrifying bacterium isolated from the oxic-anoxic interface of the Gotland Deep in the central Baltic Sea. Int. J. Syst. Evol. Microbiol. 2002, 52, 2211–2217. [Google Scholar] [CrossRef]
- Schmehl, M.; Jahn, A.; Meyer zu Vilsendorf, A.; Hennecke, S.; Masepohl, B.; Schuppler, M.; Marxer, M.; Oelze, J.; Klipp, W. Identification of a new class of nitrogen fixation genes in Rhodobacter capsulatus: A putative membrane complex involved in electron transport to nitrogenase. Mol. Gen. Genet. 1993, 241, 602–615. [Google Scholar] [CrossRef]
- Reyes-Prieto, A.; Barquera, B.; Juárez, O. Origin and evolution of the sodium-pumping NADH: Ubiquinone oxidoreductase. PLoS ONE 2014, 9, e96696. [Google Scholar] [CrossRef]
- Biegel, E.; Schmidt, S.; González, J.M.; Müller, V. Biochemistry, evolution and physiological function of the Rnf complex, a novel ion-motive electron transport complex in prokaryotes. Cell. Mol. Life Sci. 2011, 68, 613–634. [Google Scholar] [CrossRef] [PubMed]

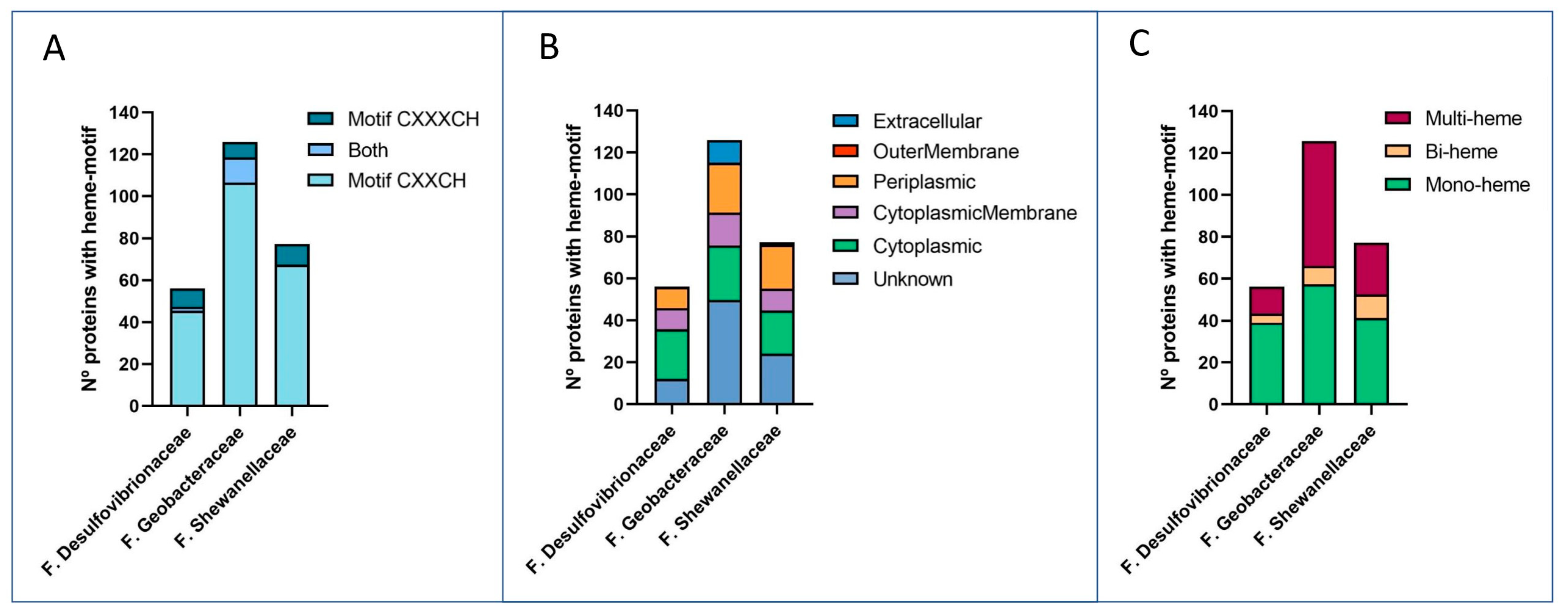
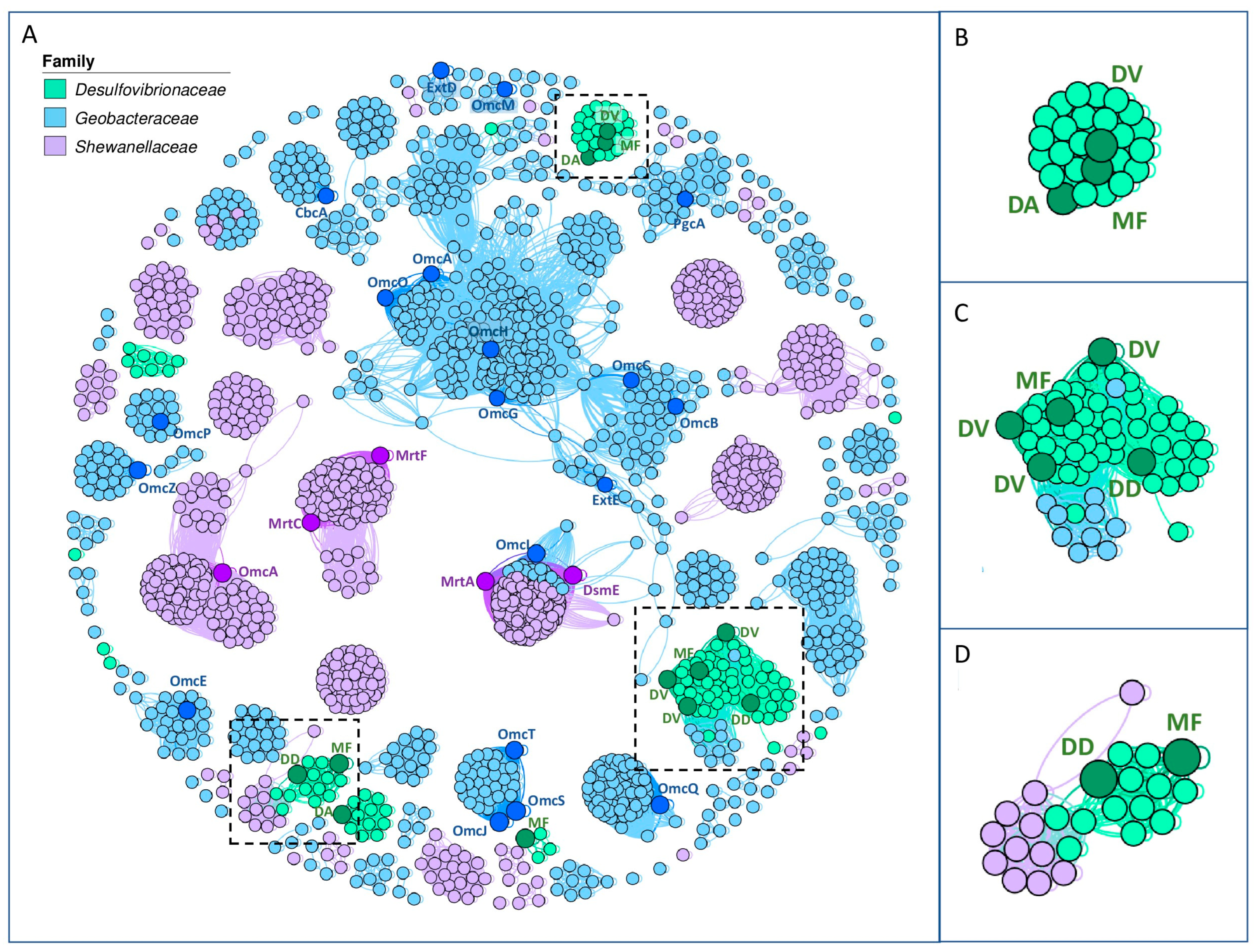

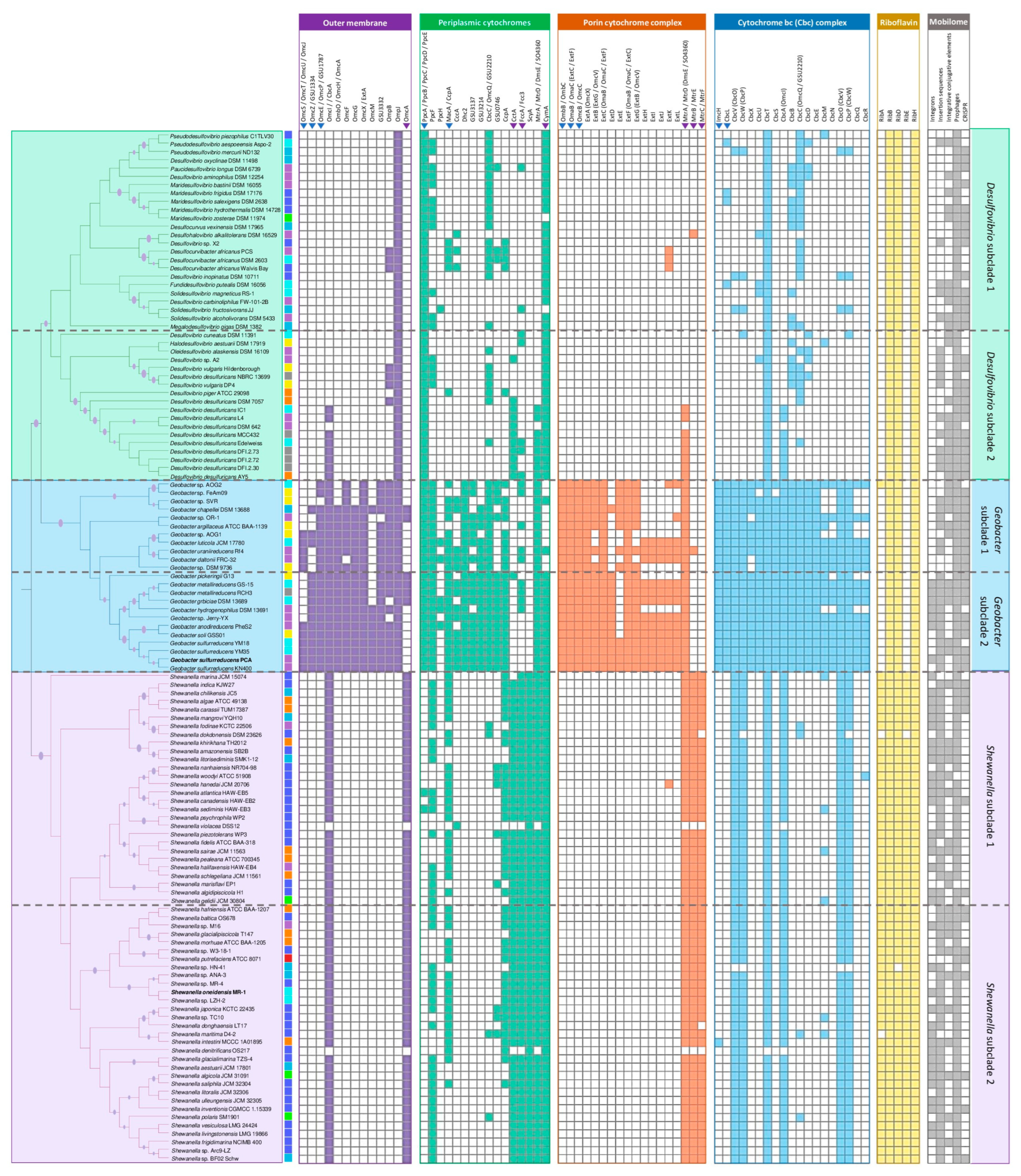




Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gonzalez, V.; Abarca-Hurtado, J.; Arancibia, A.; Claverías, F.; Guevara, M.R.; Orellana, R. Novel Insights on Extracellular Electron Transfer Networks in the Desulfovibrionaceae Family: Unveiling the Potential Significance of Horizontal Gene Transfer. Microorganisms 2024, 12, 1796. https://doi.org/10.3390/microorganisms12091796
Gonzalez V, Abarca-Hurtado J, Arancibia A, Claverías F, Guevara MR, Orellana R. Novel Insights on Extracellular Electron Transfer Networks in the Desulfovibrionaceae Family: Unveiling the Potential Significance of Horizontal Gene Transfer. Microorganisms. 2024; 12(9):1796. https://doi.org/10.3390/microorganisms12091796
Chicago/Turabian StyleGonzalez, Valentina, Josefina Abarca-Hurtado, Alejandra Arancibia, Fernanda Claverías, Miguel R. Guevara, and Roberto Orellana. 2024. "Novel Insights on Extracellular Electron Transfer Networks in the Desulfovibrionaceae Family: Unveiling the Potential Significance of Horizontal Gene Transfer" Microorganisms 12, no. 9: 1796. https://doi.org/10.3390/microorganisms12091796