
The Effects of Dietary Crude Protein Level on Ammonia Emissions from Slurry from Lactating Holstein-Friesian Cows as Measured in Open-Circuit Respiration Chambers


Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Diets and Feeding of Animals for Slurry Production
2.2. Faeces and Urine Collection
2.3. Slurry Sample Preparation
2.4. Slurry Allocation to Respiration Chambers and NH3 Emission Measurements
2.5. Laboratory Analysises
2.6. Statistical Analysis
3. Results and Discussion
3.1. Urine and Faeces Characteristics
3.2. Dietary CP Level and NH3 Emissions from Slurry
3.3. Chamber Temperature and NH3 Emissions from Slurry
3.4. Relationships of NH3 Emissions from Slurry and Dietary CP Level, Total Urinary N, and Faeces N
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Smith, H.; Jones, L.; Thistlethwaite, G.; Raoult, J.; Richardson, J.; Richmond, B.; Zhang, H. Air Pollutant Inventories for England, Scotland, Wales and Northern Ireland: 1990–2018; The Department for Environment, Food & Rural Affairs, The Scottish Government, The Welsh Government and The Northern Ireland Department for Agriculture, Environment and Rural Affairs, 2020. Available online: https://uk-air.defra.gov.uk/assets/documents/reports/cat09/1910031755_DA_Air_Pollutant_Inventories_1990-2017_Issue_1.1.pdf (accessed on 9 May 2022).
- Dragosits, U.; Dore, A.J.; Sheppard, L.J.; Vieno, M.; Tang, Y.S.; Theobald, M.R.; Sutton, M.A. Sources, Dispersion and Fate of Atmospheric Ammonia. In Nitrogen in the Environment: Sources, Problems, and Management; Hatfield, J.L., Follett, R.F., Eds.; Academic Press: Amsterdam, The Netherlands, 2008; pp. 333–393. [Google Scholar]
- Sheppard, L.J.; Leith, I.D.; Mizunuma, T.; Neil Cape, J.; Crossley, A.; Leeson, S.; Sutton, M.A.; van Dijk, N.; Fowler, D. Dry deposition of ammonia gas drives species change faster than wet deposition of ammonium ions: Evidence from a long-term field manipulation. Glob. Chang. Biol. 2011, 17, 3589–3607. [Google Scholar] [CrossRef]
- Anderson, J.O.; Thundiyil, J.G.; Stolbach, A. Clearing the air: A review of the effects of particulate matter air pollution on human health. J. Toxicol. Clin. Toxicol. 2012, 8, 166–175. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Horn, H.H.; Newton, G.L.; Kunkle, W.E. Ruminant nutrition from an environmental perspective: Factors affecting whole-farm nutrient balance. Anim. Sci. J. 1996, 74, 3082–3102. [Google Scholar] [CrossRef] [PubMed]
- Singurindy, O.; Molodovskaya, M.; Richards, B.K.; Steenhuis, T.S. Nitrous oxide emission at low temperatures from manure-amended soils under corn (Zea mays L.). Agric. Ecosyst. Environ. 2009, 132, 74–81. [Google Scholar] [CrossRef]
- Chadwick, D.; Sommer, S.; Thorman, R.; Fangueiro, D.; Cardenas, L.; Amon, B.; Misselbrook, T. Manure management: Implications for greenhouse gas emissions. Anim. Feed Sci. Technol. 2011, 166, 514–531. [Google Scholar] [CrossRef]
- Holm-Nielsen, J.B.; Al Seadi, T.; Oleskowicz-Popiel, P. The future of anaerobic digestion and biogas utilization. Bioresour. Technol. 2009, 100, 5478–5484. [Google Scholar] [CrossRef]
- Todd, R.W.; Cole, N.A.; Clark, R.N.; Flesch, T.K.; Harper, L.A.; Baek, B.H. Ammonia emissions from a beef cattle feedyard on the southern High Plains. Atmos. Environ. 2008, 42, 6797–6805. [Google Scholar] [CrossRef]
- Varel, V.H. Use of urease inhibitors to control nitrogen loss from livestock waste. Bioresour. Technol. 1997, 62, 11–17. [Google Scholar] [CrossRef]
- Petersen, S.O.; Sommer, S.G.; Aaes, O.; Søegaard, K. Ammonia losses from urine and dung of grazing cattle: Effect of N intake. Atmos. Environ. 1998, 32, 295–300. [Google Scholar] [CrossRef]
- Dijkstra, J.; Oenema, O.; Van Groenigen, J.W.; Spek, J.W.; Van Vuuren, A.M.; Bannink, A. Diet effects on urine composition of cattle and N2O emissions. Animal 2013, 7, 292–302. [Google Scholar] [CrossRef] [Green Version]
- Hristov, A.N.; Heyler, K.; Schurman, E.; Griswold, K.; Topper, P.; Hile, M.; Ishler, V.; Fabian-Wheeler, E.; Dinh, S. CASE STUDY: Reducing dietary protein decreased the ammonia emitting potential of manure from commercial dairy farms. Prof. Anim. Sci. 2015, 31, 68–79. [Google Scholar] [CrossRef]
- Burgos, S.A.; Embertson, N.M.; Zhao, Y.; Mitloehner, F.M.; DePeters, E.J.; Fadel, J.G. Prediction of ammonia emission from dairy cattle manure based on milk urea nitrogen: Relation of milk urea nitrogen to ammonia emissions. J. Dairy Sci. 2010, 93, 2377–2386. [Google Scholar] [CrossRef] [PubMed]
- Edouard, N.; Charpiot, A.; Robin, P.; Lorinquer, E.; Dollé, J.B.; Faverdin, P. Influence of diet and manure management on ammonia and greenhouse gas emissions from dairy barns. Animal 2019, 13, 2903–2912. [Google Scholar] [CrossRef] [PubMed]
- Spiehs, M.J.; Varel, V.H. Nutrient excretion and odorant production in manure from cattle fed corn wet distillers grains with solubles. J. Anim. Sci. 2009, 87, 2977–2984. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yan, T.; Frost, J.P.; Agnew, R.E.; Binnie, R.C.; Mayne, C.S. Relationships among manure nitrogen output and dietary and animal factors in lactating dairy cows. J. Dairy Sci. 2006, 89, 3981–3991. [Google Scholar] [CrossRef]
- Angelidis, A.; Crompton, L.; Misselbrook, T.; Yan, T.; Reynolds, C.K.; Stergiadis, S. Evaluation and prediction of nitrogen use efficiency and outputs in faeces and urine in beef cattle. Agric. Ecosyst. Environ. 2019, 280, 1–15. [Google Scholar] [CrossRef]
- Cichota, R.; Snow, V.O. Ammonia volatilisation from grazed pastures. Report Prepared for Dairy Australia and the New Zealand Agricultural Greenhouse Gas Research Centre—NZAGRC. 2012. Available online: https://www.nzagrc.org.nz/publications/ammonia-volatilisation-from-grazed-pastures (accessed on 9 May 2022).
- Sommer, S.G.; Hutchings, N.J. Ammonia emission from field applied manure and its reduction. Eur. J. Agron. 2001, 15, 1–15. [Google Scholar] [CrossRef]
- Fangueiro, D.; Hjorth, M.; Gioelli, F. Acidification of animal slurry—A review. J. Environ. Manag. 2015, 149, 46–56. [Google Scholar] [CrossRef]
- Katongole, C.B.; Yan, T. Effect of varying dietary crude protein level on feed intake, nutrient digestibility, milk production, and nitrogen use efficiency by lactating Holstein-Friesian cows. Animals 2020, 10, 2439. [Google Scholar] [CrossRef]
- Thomas, C. Feed into Milk: A New Applied Feeding System for Dairy Cows: An Advisory Manual; Nottingham University Press: Nottingham, UK, 2004. [Google Scholar]
- Van Soest, P.V.; Robertson, J.B.; Lewis, B. Methods for dietary fiber, neutral detergent fiber, and nonstarch polysaccharides in relation to animal nutrition. J. Dairy Sci. 1991, 74, 3583–3597. [Google Scholar] [CrossRef]
- Lee, C.; Hristov, A.N.; Heyler, K.S.; Cassidy, T.W.; Long, M.; Corl, B.A.; Karnati, S.K.R. Effects of dietary protein concentration and coconut oil supplementation on nitrogen utilization and production in dairy cows. J. Dairy Sci. 2011, 94, 5544–5557. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bach, A.; Calsamiglia, S.; Stern, M.D. Nitrogen metabolism in the rumen. J. Dairy Sci. 2005, 88, E9–E21. [Google Scholar] [CrossRef] [Green Version]
- Valadares, R.; Broderick, G.; Filho, S.V.; Clayton, M. Effect of replacing Alfalfa silage with high moisture corn on ruminal protein synthesis estimated from excretion of total purine derivatives. J. Dairy Sci. 1999, 82, 2686–2696. [Google Scholar] [CrossRef]
- Colmenero, J.O.; Broderick, G.A. Effect of dietary crude protein concentration on milk production and nitrogen utilization in lactating dairy cows. J. Dairy Sci. 2006, 89, 1704–1712. [Google Scholar] [CrossRef] [Green Version]
- Van Vliet, P.C.J.; Reijs, J.W.; Bloem, J.; Dijkstra, J.; De Goede, R.G.M. Effects of cow diet on the microbial community and organic matter and nitrogen content of feces. J. Dairy Sci. 2007, 90, 5146–5158. [Google Scholar] [CrossRef] [Green Version]
- Koenig, K.M.; McGinn, S.M.; Beauchemin, K.A. Ammonia emissions and performance of backgrounding and finishing beef feedlot cattle fed barley-based diets varying in dietary crude protein concentration and rumen degradability. J. Anim. Sci. 2013, 91, 2278–2294. [Google Scholar] [CrossRef]
- Mobley, H.L.T.; Hausinger, R.P. Microbial ureases: Significance, regulation and molecular characterization. Microbiol. Rev. 1989, 53, 85–108. [Google Scholar] [CrossRef]
- Reynolds, C.K.; Kristensen, N.B. Nitrogen recycling through the gut and the nitrogen economy of ruminants: An asynchronous symbiosis. J. Anim. Sci. 2008, 86, E293–E305. [Google Scholar] [CrossRef] [Green Version]
- O’Connell, J.M.; Callan, J.J.; O’Doherty, J.V. The effect of dietary crude protein level, cereal type and exogenous enzyme supplementation on nutrient digestibility, nitrogen excretion, faecal volatile fatty acid concentration and ammonia emissions from pigs. Anim. Feed Sci. Technol. 2006, 127, 73–88. [Google Scholar] [CrossRef]
- Spek, J.W.; Dijkstra, J.; Van Duinkerken, G.; Bannink, A. A review of factors influencing milk urea concentration and its relationship with urinary urea excretion in lactating dairy cattle. J. Agric. Sci. 2013, 151, 407–423. [Google Scholar] [CrossRef] [Green Version]
- Arriaga, H.; Salcedo, G.; Martínez-Suller, L.; Calsamiglia, S.; Merino, P. Effect of dietary crude protein modification on ammonia and nitrous oxide concentration on a tie-stall dairy barn floor. J. Dairy Sci. 2010, 93, 3158–3165. [Google Scholar] [CrossRef] [PubMed]
- Dai, X.; Karring, H. A determination and comparison of urease activity in feces and fresh manure from pig and cattle in relation to ammonia production and pH changes. PLoS ONE 2014, 9, e110402. [Google Scholar] [CrossRef] [PubMed]
- Moraes, L.E.; Burgos, S.A.; DePeters, E.J.; Zhang, R.; Fadel, J.G. Urea hydrolysis in dairy cattle manure under different temperature, urea, and pH conditions. J. Dairy Sci. 2017, 100, 2388–2394. [Google Scholar] [CrossRef] [PubMed]
- Rochette, P.; Angers, D.A.; Chantigny, M.H.; MacDonald, J.D.; Gasser, M.O.; Bertrand, N. Reducing ammonia volatilization in a no-till soil by incorporating urea and pig slurry in shallow bands. Nutr. Cycl. Agroecosyst. 2009, 84, 71–80. [Google Scholar] [CrossRef]
- Chantigny, M.H.; MacDonald, J.D.; Beaupré, C.; Rochette, P.; Angers, D.A.; Massé, D.; Parent, L.E. Ammonia volatilization following surface application of raw and treated liquid swine manure. Nutr. Cycl. Agroecosyst. 2009, 85, 275–286. [Google Scholar] [CrossRef]
- Moal, J.F.; Martinez, J.; Guiziou, F.; Coste, C.M. Ammonia volatilization following surface-applied pig and cattle slurry in France. J. Agric. Sci. 1995, 125, 245–252. [Google Scholar] [CrossRef]
- Carozzi, M.; Ferrara, R.M.; Fumagalli, M.; Sanna, M.; Chiodini, M.; Perego, A.; Chierichetti, A.; Brenna, S.; Rana, G.; Acutis, M. Field-scale ammonia emissions from surface spreading of dairy slurry in Po Valley. Ital. J. Agrometeorol. 2012, 4, 25–34. [Google Scholar]
- Sommer, S.G.; Genermont, S.; Cellier, P.; Hutchings, N.J.; Olesen, J.E.; Morvan, T. Processes controlling ammonia emission from livestock slurry in the field. Eur. J. Agron. 2003, 19, 465–486. [Google Scholar] [CrossRef]
- Christensen, M.L.; Hjorth, M.; Keiding, K. Characterization of pig slurry with reference to flocculation and separation. Water Res. 2009, 43, 773–783. [Google Scholar] [CrossRef]
- Hjorth, M.; Christensen, K.V.; Christensen, M.L.; Sommer, S.G. Solid-liquid separation of animal slurry in theory and practice. A review. Agron. Sustain. Dev. 2011, 30, 153–180. [Google Scholar] [CrossRef] [Green Version]
- Liu, M.; Giard, D.; Barrington, S. Ammonium dissociation for swine and dairy cattle manures. J. Environ. Prot. Sci. 2013, 4, 6–15. [Google Scholar] [CrossRef] [Green Version]
- Vaddella, V.K.; Ndegwa, P.M.; Jiang, A. An empirical model of ammonium ion dissociation in liquid dairy manure. Trans. ASABE 2011, 54, 1119–1126. [Google Scholar] [CrossRef] [Green Version]
- Arogo, J.; Westerman, P.W.; Liang, Z.S. Comparing ammonium ion dissociation constant in swine anaerobic lagoon liquid and deionized water. Trans. ASABE 2003, 46, 1415–1419. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
| Dietary CP Content (Formulated), g/kg DM | ||||
|---|---|---|---|---|
| 141 | 151 | 177 | 201 | |
| Ingredient, % of TMR DM | ||||
| Ryegrass silage | 47.3 | 47.9 | 47.7 | 47.5 |
| Concentrate mix | 52.7 | 52.1 | 52.3 | 52.5 |
| Chemical composition 1 | ||||
| Dry matter (DM), g/kg | 507 | 513 | 510 | 508 |
| Crude protein (CP), g/kg DM | 141 | 151 | 177 | 201 |
| Rumen-degradable protein (RDP) 2, g/kg CP | 103 | 109 | 127 | 144 |
| Rumen-undegradable protein (RUP) 2, g/kg CP | 39 | 41 | 49 | 56 |
| Neutral detergent fibre (NDF), g/kg DM | 411 | 408 | 399 | 388 |
| Acid detergent fibre (ADF), g/kg DM | 218 | 218 | 214 | 209 |
| Water-soluble carbohydrates) WSC, g/kg DM | 81 | 82 | 86 | 90 |
| Ash, g/kg DM | 78 | 78 | 80 | 81 |
| Dietary CP Level, g/kg DM | p-Value | |||||||
|---|---|---|---|---|---|---|---|---|
| 141 | 151 | 177 | 201 | SEM | Diet | Linear | Quadratic | |
| Urine | ||||||||
| Weight, kg/cow/day | 16.7 b | 16.8 b | 21.6 ab | 23.2 a | 1.85 | 0.039 | 0.006 | 0.805 |
| Total urine N, g/L | 8.0 b | 8.5 b | 9.5 b | 12.0 a | 0.53 | 0.0001 | <0.0001 | 0.179 |
| Faeces | ||||||||
| Weight, kg/cow/day | 34.9 | 36.3 | 46.3 | 39.4 | 4.66 | 0.343 | 0.275 | 0.180 |
| Total faeces N, g/kg (fresh) | 4.9 | 4.8 | 5.1 | 5.2 | 0.36 | 0.812 | 0.393 | 0.942 |
| DM, g/kg | 149.5 | 145.7 | 140.8 | 152.9 | 7.24 | 0.654 | 0.770 | 0.242 |
| NDF, g/kg DM | 545.7 | 559.6 | 570.2 | 534.3 | 10.1 | 0.108 | 0.412 | 0.082 |
| ADF, g/kg DM | 286.3 | 299.1 | 298.7 | 279.9 | 6.50 | 0.111 | 0.303 | 0.094 |
| Total slurry 1 | ||||||||
| Weight, kg/cow/day | 51.6 b | 53.1 b | 67.9 a | 62.6 ab | 5.48 | 0.043 | 0.038 | 0.070 |
| Dietary CP Level, g/kg DM | p-Value | |||||||
|---|---|---|---|---|---|---|---|---|
| 141 | 151 | 177 | 201 | SEM | Diet | Linear | Quadratic | |
| 0–24 h | ||||||||
| NH3, g/cow | 21.3 c | 29.5 bc | 42.2 b | 63.4 a | 4.97 | 0.0002 | <0.0001 | 0.538 |
| 24–48 h | ||||||||
| NH3, g/cow | 9.4 c | 13.8 bc | 23.5 ab | 33.1 a | 3.80 | 0.003 | 0.0003 | 0.992 |
| 0–48 h 1 | ||||||||
| NH3, g/cow | 30.6 c | 43.4 bc | 65.8 b | 96.6 a | 7.91 | 0.0002 | <0.0001 | 0.712 |
| Average daily emissions | ||||||||
| NH3, g/cow/day | 15.3 c | 21.7 bc | 32.9 b | 48.3 a | 3.96 | 0.0002 | <0.0001 | 0.712 |
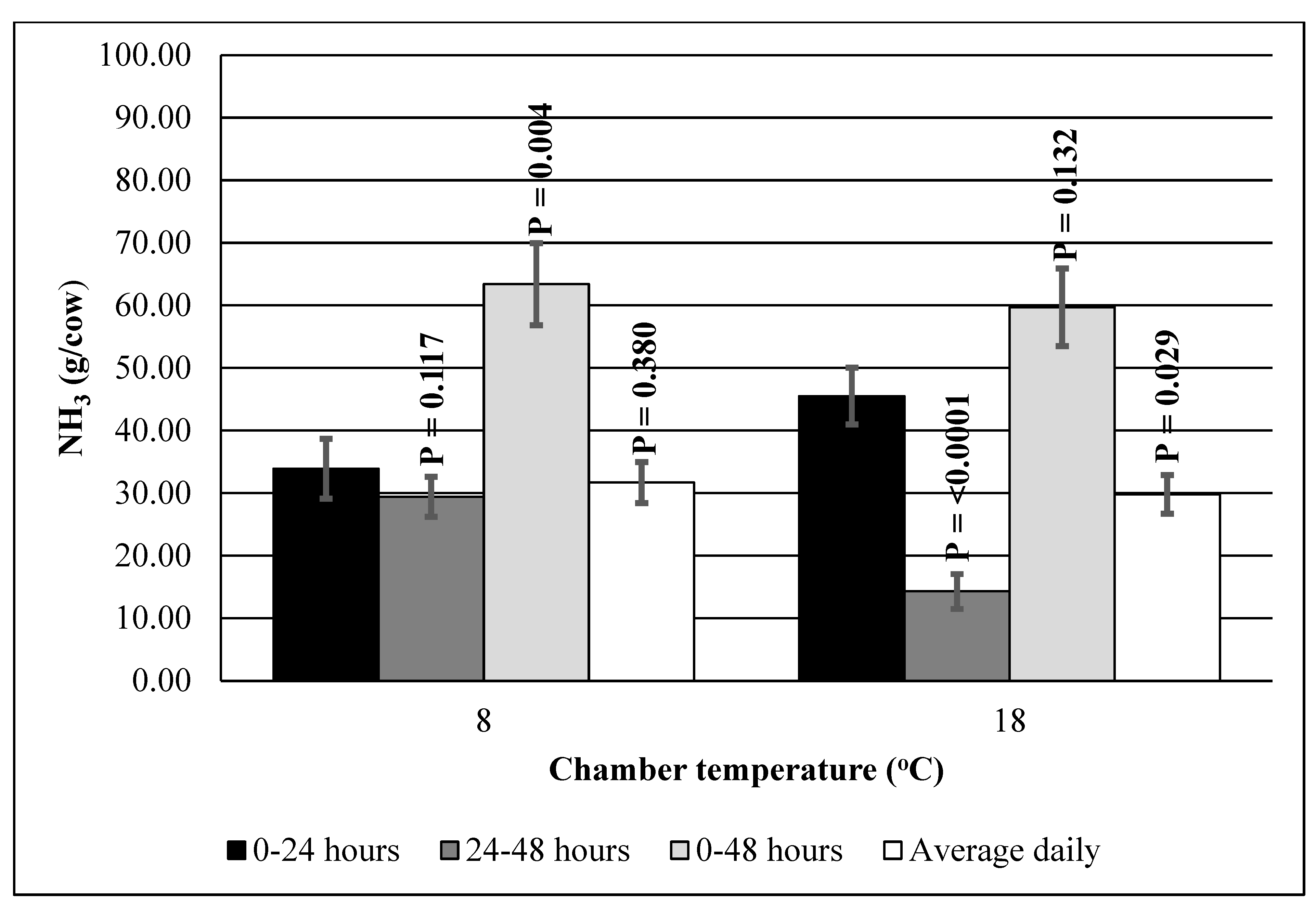
| Chamber Temperature, °C | ||||
|---|---|---|---|---|
| 8 | 18 | SEM | p-Value | |
| Measurement time interval | ||||
| 0–24 h | ||||
| NH3, g/cow | 33.9 b | 45.4 a | 4.78 | 0.009 |
| 24–48 h | ||||
| NH3, g/cow | 29.4 a | 14.3 b | 3.20 | 0.001 |
| 0–48 h | ||||
| NH3, g/cow | 63.4 | 59.7 | 6.56 | 0.475 |
| Average daily emissions | ||||
| NH3, g/cow/day | 31.7 | 29.8 | 3.28 | 0.475 |
| Linear Equation | R2 | |||||||
|---|---|---|---|---|---|---|---|---|
| NH3, g/Cow/Day = | ||||||||
| (1) | −60.09 (11.1) | + | 0.54 (0.07) × CP a | 0.67 | ||||
| (2) | −30.98 (8.86) | + | 6.55 (0.91) × UN b | 0.62 | ||||
| (3) | +30.04 (19.4) | + | 0.32 (3.77) × FN c | 0.0002 | ||||
| (4) | −56.07 (10.9) | + | 0.37 (0.11) × CP a | + | 2.66 (1.44) × UN b | 0.72 | ||
| (5) | −47.2 (14.0) | + | 0.56 (0.06) × CP a | − | 3.13 (2.12) × FN c | 0.71 | ||
| (6) | −31.7 (15.0) | + | 6.55 (0.92) × UN b | + | 0.14 (2.36) × FN c | 0.62 | ||
| (7) | −47.4 (13.7) | + | 0.41 (0.12) × CP a | + | 2.24 (1.49) × UN b | − | 2.25 (2.15) × FN c | 0.73 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Katongole, C.B.; Yan, T. The Effects of Dietary Crude Protein Level on Ammonia Emissions from Slurry from Lactating Holstein-Friesian Cows as Measured in Open-Circuit Respiration Chambers. Animals 2022, 12, 1243. https://doi.org/10.3390/ani12101243
Katongole CB, Yan T. The Effects of Dietary Crude Protein Level on Ammonia Emissions from Slurry from Lactating Holstein-Friesian Cows as Measured in Open-Circuit Respiration Chambers. Animals. 2022; 12(10):1243. https://doi.org/10.3390/ani12101243
Chicago/Turabian StyleKatongole, Constantine Bakyusa, and Tianhai Yan. 2022. "The Effects of Dietary Crude Protein Level on Ammonia Emissions from Slurry from Lactating Holstein-Friesian Cows as Measured in Open-Circuit Respiration Chambers" Animals 12, no. 10: 1243. https://doi.org/10.3390/ani12101243