
Performance of a Differentiation of Infected from Vaccinated Animals (DIVA) Classical Swine Fever Virus (CSFV) Serum and Oral Fluid Erns Antibody AlphaLISA Assay

 ,
,  , , and
, , and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Experimental Design
2.2. Biological Samples
2.3. Virus Neutralization Test (VNT)
2.4. CSFV Erns Antigen
2.5. CSFV Erns AlphaLISA Bead Conjugation
2.6. CSFV Erns IgG AlphaLISA Optimization
2.7. Statistical Analysis
3. Results
3.1. CSFV Antibody Responses
3.2. Evaluation of Test Diagnostic Performance
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Postel, A.; Smith, D.B.; Becher, P. Proposed update to the taxonomy of pestiviruses: Eight additional species with the genus Pestivirus, family Flaviviridae. Viruses 2021, 13, 1542. [Google Scholar] [CrossRef] [PubMed]
- Smith, D.B.; Meyers, G.; Bukh, J.; Gould, E.A.; Monath, T.; Muerhoff, A.S.; Pletnev, A.; Rico-Hesse, R.; Stapleton, J.T.; Simmonds, P.; et al. Proposed revision to the taxonomy of the genus Pestivirus, family Flaviviridae. J. Gen. Virol. 2017, 98, 2106–2112. [Google Scholar] [CrossRef] [PubMed]
- de Schweinitz, E.A.; Dorset, M.A. A Form of Hog Cholera Not Caused the Hog Cholera Bacillus; U.S. Bureau of Animal Industry Circular no. 41; U.S. Department of Agriculture, Government Printing Office: Washington, DC, USA, 1903. [Google Scholar]
- Stegeman, A.; Elbers, A.; de Smit, H.; Moser, H.; Smak, J.; Pluimers, F. The 1997–1998 epidemic of classical swine fever in the Netherlands. Vet. Microbiol. 2000, 73, 183–196. [Google Scholar] [CrossRef] [PubMed]
- Paton, D. The reappearance of classical swine fever in England in 2000. In Trends in Emerging Viral Infections of Swine; Morilla, A., Yoon, K.-J., Zimmerman, J., Eds.; Iowa State Press: Ames, IA, USA, 2002; pp. 153–158. [Google Scholar]
- Sandvik, T.; Crooke, H.; Drew, T.W.; Blome, S.; Greiser-Wilke, I.; Moennig, V.; Gous, T.A.; Gers, S.; Kitching, J.A.; Bührmann, G.; et al. Classical swine fever in South Africa after 87 years’ absence. Vet. Rec. 2005, 157, 267. [Google Scholar] [CrossRef] [PubMed]
- David, D.; Edri, N.; Yakobson, B.A.; Bombarov, V.; King, R.; Davidson, I.; Pozzi, P.; Hadani, Y.; Bellaiche, M.; Schmeiser, S.; et al. Emergence of classical swine fever virus in Israel in 2009. Vet. J. 2011, 190, e146–e149. [Google Scholar] [CrossRef] [PubMed]
- Postel, A.; Austermann-Busch, S.; Petrov, A.; Moennig, V.; Becher, P. Epidemiology, diagnosis and control of classical swine fever: Recent developments and future challenges. Transbound. Emerg. Dis. 2018, 65 (Suppl. S1), 248–261. [Google Scholar] [CrossRef] [PubMed]
- van Oirschot, J.T.; De Waal, C.A. An ELISA to distinguish between Aujeszky’s disease vaccinated and infected pigs. Vet. Rec. 1987, 121, 305–306. [Google Scholar] [CrossRef] [PubMed]
- van Oirschot, J.T.; Gielkens, A.L.; Moormann, R.J.; Berns, A.J. Marker vaccines, virus protein-specific antibody assays and the control of Aujeszky’s disease. Vet. Microbiol. 1990, 23, 85–101. [Google Scholar] [CrossRef]
- van Oirschot, J.T. Diva vaccines that reduce virus transmission. J. Biotechnol. 1999, 73, 195–205. [Google Scholar] [CrossRef]
- Blome, S.; Moβ, C.; Reimann, I.; König, P.; Beer, M. Classical swine fever vaccines—State-of-the art. Vet. Microbiol. 2017, 206, 10–20. [Google Scholar] [CrossRef]
- Rümenapf, T.; Unger, G.; Strauss, J.H.; Thiel, H.J. Processing of the envelope glycoproteins of pestiviruses. J. Virol. 1993, 67, 3288–3294. [Google Scholar] [CrossRef] [PubMed]
- Meyers, G.; Thiel, H.J. Molecular characterization of pestiviruses. Adv. Virus Res. 1996, 47, 53–118. [Google Scholar] [CrossRef] [PubMed]
- Tautz, N.; Elbers, K.; Stoll, D.; Meyers, G.; Thiel, H.J. Serine protease of pestiviruses: Determination of cleavage sites. J. Virol. 1997, 71, 5415–5422. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Cao, H.W.; Wu, Z.J.; Cui, Y.D. A review of molecular characterization of classical swine fever virus (CSFV). Isr. J. Vet. Med. 2011, 66, 89–95. [Google Scholar]
- Blome, S.; Wernike, K.; Reimann, I.; König, P.; Moβ, C.; Beer, M. A decade of research into classical swine fever marker vaccine CP7_E2alf (Suvaxyn® CSF Marker): A review of vaccine properties. Vet. Res. 2017, 48, 51. [Google Scholar] [CrossRef] [PubMed]
- Floegel-Niesmann, G. Classical swine fever (CSF) marker vaccine. Trial III. Evaluation of discriminatory ELISAs. Vet. Microbiol. 2001, 83, 121–136. [Google Scholar] [CrossRef] [PubMed]
- Schroeder, S.; von Rosen, T.; Blome, S.; Loeffen, W.; Haegeman, A.; Koenen, F.; Uttenthal, A. Evaluation of classical swine fever virus antibody detection assays with an emphasis on the differentiation of infected from vaccinated animals. Rev. Sci. Tech. 2012, 31, 997–1010. [Google Scholar] [CrossRef]
- Pannhorst, K.; Frohlich, A.; Staubach, C.; Meyer, D.; Blome, S.; Becher, P. Evaluation of an Ern-based enzyme-linked immunosorbent assay to distinguish Classical swine fever virus-infected pigs from pigs vaccinated with CP7_E2alf. J. Vet. Diagn. Investig. 2015, 27, 449–460. [Google Scholar] [CrossRef]
- Meyer, D.; Fritsche, S.; Luo, Y.; Engemann, C.; Blome, S.; Beyerbach, M.; Chang, C.-Y.; Qiu, H.-J.; Becher, P.; Poste, A. The double-antigen ELISA concept for early detection of Erns-specific classical swine fever virus antibodies and application as an accompanying test for differentiation of infected from marker vaccinated animals. Transbound. Emerg. Dis. 2017, 64, 2013–2022. [Google Scholar] [CrossRef]
- Beaudet, L.; Rodriguez-Suarez, R.; Venne, M.-H.; Caron, M.; Bédard, J.; Brechler, V.; Parent, S.; Bielefeld-Sévigny, M. AlphaLISA® immunoassays: The no-wash alternative to ELISAs for research and drug discovery. Nat. Methods 2008, 5, 8–9. [Google Scholar] [CrossRef]
- Kimpston-Burkgren, K.; Mora-Diaz, J.C.; Roby, P.; Bjustrom-Kraft, J.; Main, R.; Bosse, R.; Gimenez-Lirola, L.G. Characterization of the humoral immune response to porcine epidemic diarrhea virus infection under experimental and field conditions using an AlphaLISA® platform. Pathogens 2020, 9, 233. [Google Scholar] [CrossRef] [PubMed]
- Chen, D.; Wang, D.; Wang, C.; Wei, F.; Zhao, H.; Lin, X.; Wu, S. Application of an AlphaLISA® for rapid sensitive detection of African swine fever virus in porcine serum. Appl. Microbiol. Biotechnol. 2021, 105, 4751–4759. [Google Scholar] [CrossRef] [PubMed]
- Panyasing, Y.; Kedkovid, R.; Thanawongnuwech, R.; Kittawornrat, A.; Ji, J.; Giménez-Lirola, L.; Zimmerman, J. Effective surveillance for early classical swine fever virus detection will utilize both virus and antibody detection capabilities. Vet. Microbiol. 2018, 216, 72–78. [Google Scholar] [CrossRef] [PubMed]
- Panyasing, Y.; Thanawongnuwech, R.; Ji, J.; Gimenez-Lirola, L.; Zimmerman, J. Detection of classical swine fever virus (CSFV) E2 and Erns antibody (IgG, IgA) in oral fluid specimens from inoculated (ALD strain) or vaccinated (LOM strain) pigs. Vet. Microbiol. 2018, 224, 70–77. [Google Scholar] [CrossRef] [PubMed]
- Robin, X.; Turck, N.; Hainard, A.; Tiberti, N.; Lisacek, F.; Sanchez, J.-C.; Müller, M. pROC: An open-source package for R and S+ to analyze and compare ROC curves. BMC Bioinform. 2011, 12, 77. [Google Scholar] [CrossRef] [PubMed]
- Du, Y. Choosing Cutoff Values for Correlated Continuous Diagnostic Data to Estimate Sensitivity and Specificity. Ph.D. Thesis, Iowa State University, Ames, IA, USA, 2018. [Google Scholar]
- Moennig, V. Introduction to classical swine fever: Virus, disease, and control policy. Vet Microbiol. 2000, 73, 93–102. [Google Scholar] [CrossRef] [PubMed]
- König, M.; Lengsfeld, T.; Pauly, T.; Stark, R.; Thiel, H.J. Classical swine fever virus: Independent induction of protective immunity by two structural glycoproteins. J. Virol. 1995, 69, 6479–6486. [Google Scholar] [CrossRef] [PubMed]
- Henao-Diaz, A.; Gimenez-Lirola, L.; Baum, D.H.; Zimmerman, J. Guidelines for oral fluid-based surveillance of viral pathogens in swine. Porc. Health Manag. 2020, 6, 28. [Google Scholar] [CrossRef]
- Reimann, I.; Depner, K.; Trapp, S.; Beer, M. An avirulent chimeric Pestivirus with altered cell tropism protects pigs against lethal infection with classical swine fever virus. Virology 2004, 322, 143–157. [Google Scholar] [CrossRef]
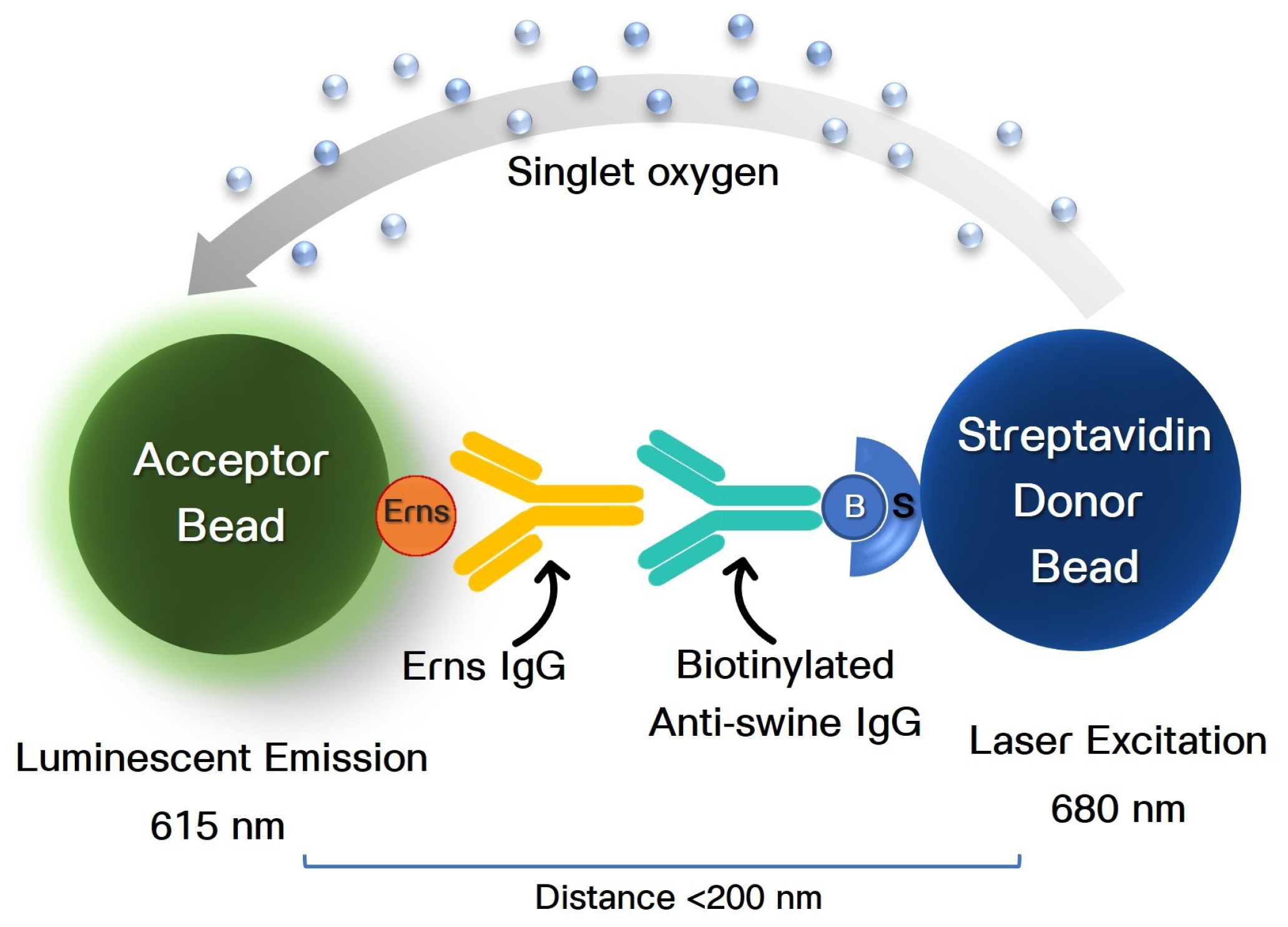


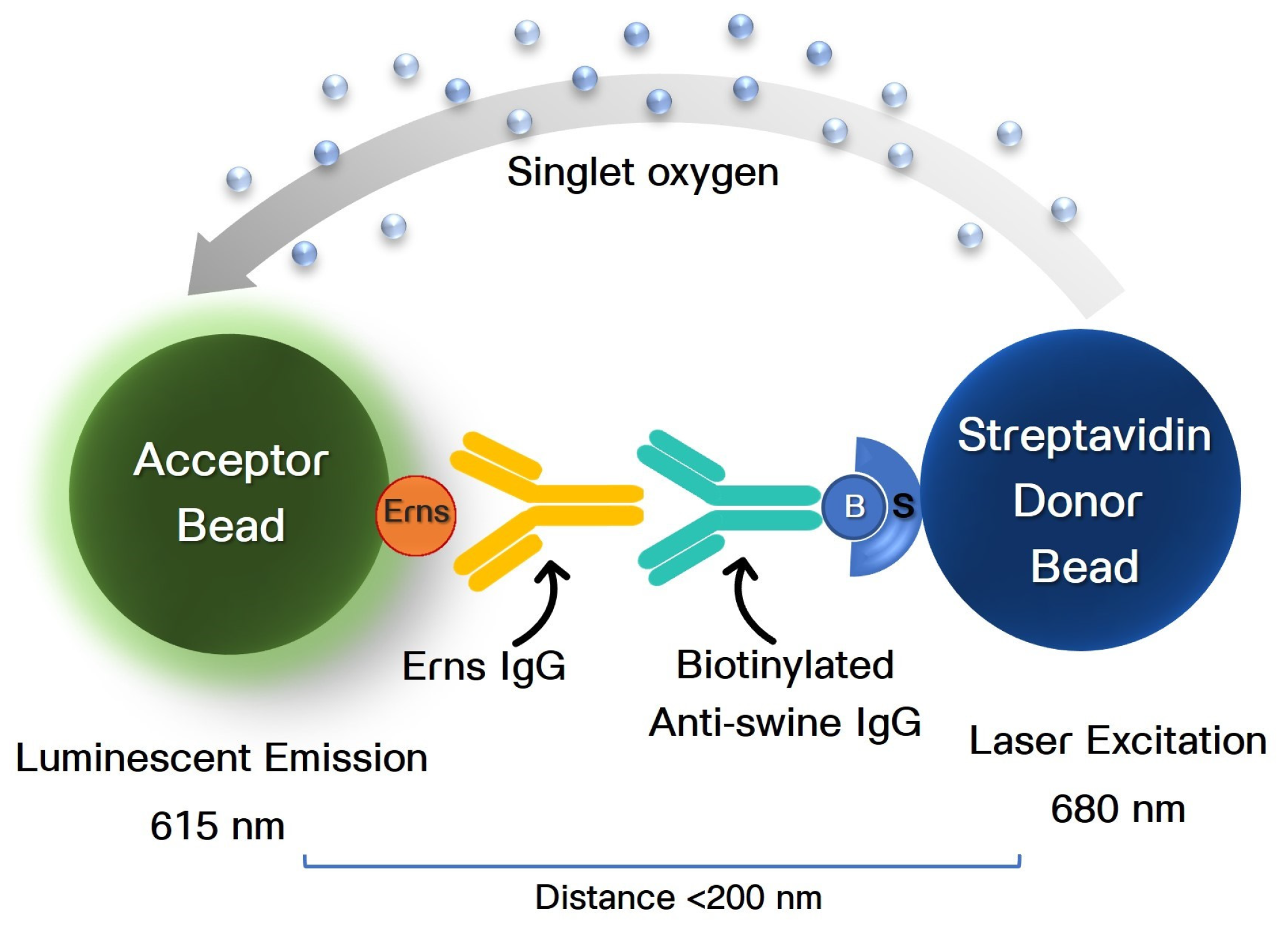{kind=link}
{kind=link}
| Group and CSFV Status | Specimen (No. Samples) | Virus Neutralization Geometric Mean Titer (log2 x) (95% CI) | Erns IgG AlphaLISA Mean Sample-to-Positive (S/P) Ratio (95% CI) |
|---|---|---|---|
| 1. Negative (unvaccinated and uninoculated). | Serum (n = 160) | negative | −0.01 a (−0.01, 0.00) |
| OF from individual pigs (n = 160) | • | −0.00 a (−0.00, 0.00) | |
| 2. Inoculated CSFV ALD wild-type strain. | Serum (n = 100) | 6.17 (5.82, 6.52) | 7.48 b (6.55, 8.40) |
| OF from individual pigs (n = 184) | • | 7.29 b (5.90, 8.68) | |
| 3. Vaccinated CSFV LOM live vaccine. | Serum (n = 100) | 4.01 (3.79, 4.23) | 1.93 c (1.76, 2.11) |
| OF from individual pigs (n = 184) | • | 1.89 c (1.80, 1.99) | |
| 4. Vaccinated CSF marker live vaccine. | Serum (n = 360) | 5.09 (4.91, 5.27) | 0.05 a (0.04, 0.06) |
| OF from pens of pigs (n = 30) | • | 0.01 a (−0.01, 0.04) |
| Cutoff (S/P) | Serum | Oral Fluids | ||
|---|---|---|---|---|
| Dx Se (95% CI) | Dx Sp (95% CI) | Dx Se (95% CI) | Dx Sp (95% CI) | |
| ≥0.1 | 100 (100, 100) * | 88.3 (85.1, 90.8) | 99.7 (99.2, 99.9) | 100 (100, 100) * |
| ≥0.2 | 100 (100, 100) * | 94.8 (93.0, 96.2) | 99.7 (99.4, 99.9) | 100 (100, 100) * |
| ≥0.3 | 97.9 (96.3, 98.8) | 99.8 (98.6, 99.9) | 99.7 (99.5, 99.9) | 100 (100, 100) * |
| ≥0.4 | 97.9 (96.5, 98.8) | 99.8 (99.7, 99.9) | 99.7 (99.5, 99.8) | 100 (100, 100) * |
| ≥0.5 | 97.4 (95.7, 98.4) | 99.8 (99.7, 99.9) | 99.5 (99.1, 99.7) | 100 (100, 100) * |
| ≥0.6 | 97.4 (95.8, 98.4) | 99.8 (99.7, 99.9) | 97.0 (95.2, 98.1) | 100 (100, 100) * |
| ≥0.7 | 97.4 (95.9, 98.3) | 100 (100, 100) * | 95.4 (92.9, 97.0) | 100 (100, 100) * |
| ≥0.8 | 97.4 (96.0, 98.3) | 100 (100, 100) * | 93.5 (90.2, 95.7) | 100 (100, 100) * |
| ≥0.9 | 97.4 (96.0, 98.3) | 100 (100, 100) * | 92.1 (88.5, 94.7) | 100 (100, 100) * |
| ≥1.0 | 97.4 (96.1, 98.3) | 100 (100, 100) * | 91.3 (87.5, 94.0) | 100 (100, 100) * |
| Group and CSFV Status | Specimen (No. Samples) | VNT | Erns IgG AlphaLISA by Cutoff | ||
|---|---|---|---|---|---|
| S/P ≥ 0.3 | S/P ≥ 0.5 | S/P ≥ 0.7 | |||
| 1. Negative (unvaccinated and uninoculated). | Serum (n = 160) | 0.0 (0.0, 2.3) | 0.0 (0.0, 2.3) | 0.0 (0.0, 2.3) | 0.0 (0.0, 2.3) |
| OF from individual pigs (n = 160) | • | 0.0 (0.0, 2.3) | 0.0 (0.0, 2.3) | 0.0 (0.0, 2.3) | |
| 2. Inoculated CSFV ALD wild-type strain. | Serum (n = 100) | 100 (96.4, 100) | 98.0 (93.0, 99.8) | 98.0 (93.0, 99.8) | 98.0 (93.0, 99.8) |
| OF from individual pigs (n = 184) | • | 100 (98.0, 100) | 100 (98.0, 100) | 92.9 (88.2, 96.2) | |
| 3. Vaccinated CSFV LOM live vaccine. | Serum (n = 100) | 100 (97.4, 100) | 96.4 (91.9, 98.3) | 96.4 (91.9, 98.3) | 96.4 (91.9, 98.3) |
| OF from individual pigs (n = 184) | • | 99.5 (97.0, 100) | 99.5 (97.0, 100) | 96.7 (93.0, 98.8) | |
| 4. Vaccinated CSF marker live vaccine. | Serum (n = 360) | 100 (99.0, 100) | 1.1 (0.3, 2.2) | 0.3 (0.0, 1.5) | 0.0 (0.0, 1.0) |
| OF from pens of pigs (n = 30) | • | 0.0 (0.0, 11.6) | 0.0 (0.0, 11.6) | 0.0 (0.0, 11.6) | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Panyasing, Y.; Gimenez-Lirola, L.; Thanawongnuwech, R.; Prakobsuk, P.; Kawilaphan, Y.; Kittawornrat, A.; Cheng, T.-Y.; Zimmerman, J. Performance of a Differentiation of Infected from Vaccinated Animals (DIVA) Classical Swine Fever Virus (CSFV) Serum and Oral Fluid Erns Antibody AlphaLISA Assay. Animals 2023, 13, 3802. https://doi.org/10.3390/ani13243802
Panyasing Y, Gimenez-Lirola L, Thanawongnuwech R, Prakobsuk P, Kawilaphan Y, Kittawornrat A, Cheng T-Y, Zimmerman J. Performance of a Differentiation of Infected from Vaccinated Animals (DIVA) Classical Swine Fever Virus (CSFV) Serum and Oral Fluid Erns Antibody AlphaLISA Assay. Animals. 2023; 13(24):3802. https://doi.org/10.3390/ani13243802
Chicago/Turabian StylePanyasing, Yaowalak, Luis Gimenez-Lirola, Roongroje Thanawongnuwech, Phakawan Prakobsuk, Yanee Kawilaphan, Apisit Kittawornrat, Ting-Yu Cheng, and Jeffrey Zimmerman. 2023. "Performance of a Differentiation of Infected from Vaccinated Animals (DIVA) Classical Swine Fever Virus (CSFV) Serum and Oral Fluid Erns Antibody AlphaLISA Assay" Animals 13, no. 24: 3802. https://doi.org/10.3390/ani13243802
APA StylePanyasing, Y., Gimenez-Lirola, L., Thanawongnuwech, R., Prakobsuk, P., Kawilaphan, Y., Kittawornrat, A., Cheng, T. -Y., & Zimmerman, J. (2023). Performance of a Differentiation of Infected from Vaccinated Animals (DIVA) Classical Swine Fever Virus (CSFV) Serum and Oral Fluid Erns Antibody AlphaLISA Assay. Animals, 13(24), 3802. https://doi.org/10.3390/ani13243802