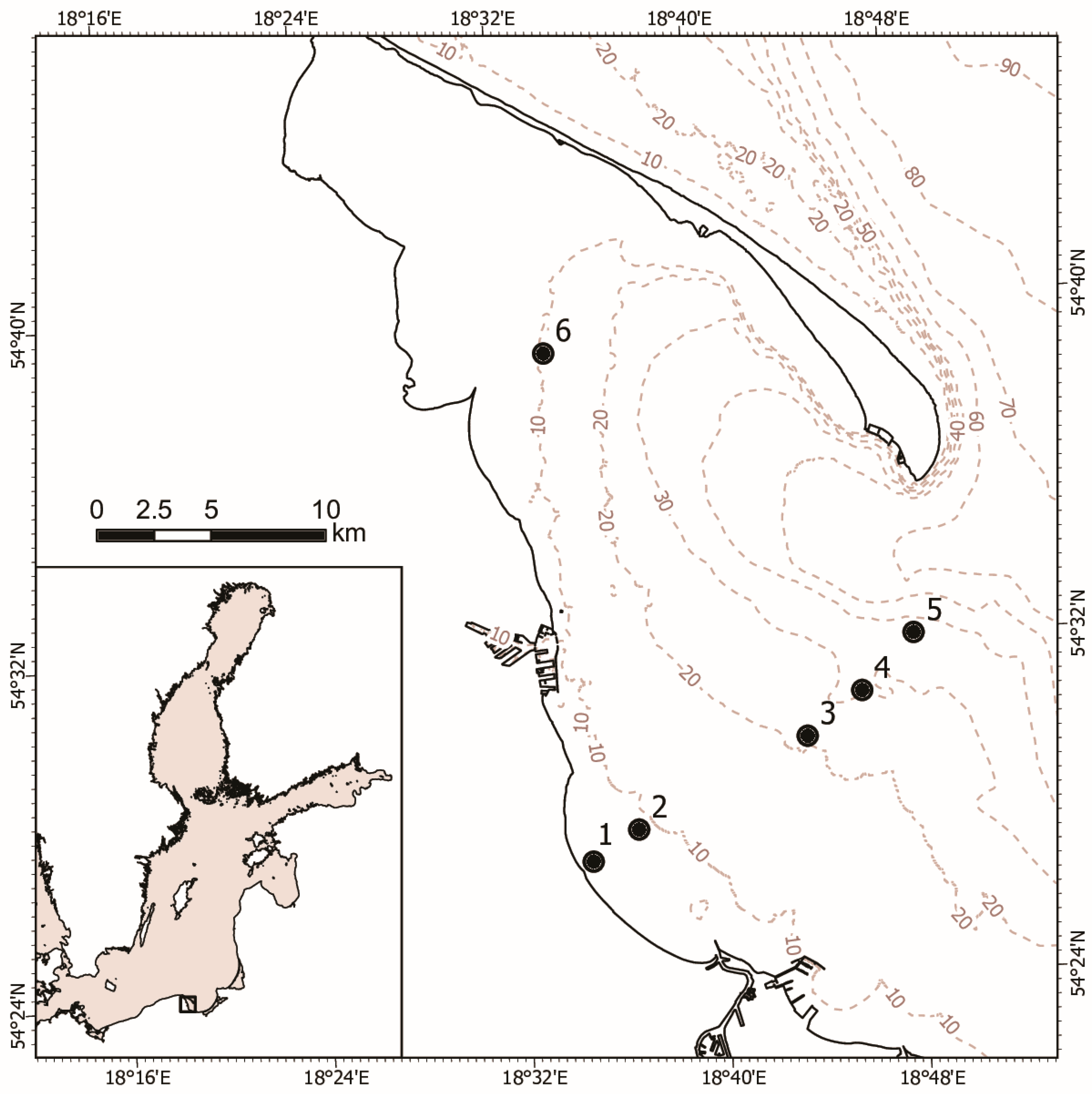
1. Introduction
The Baltic Sea is a unique ecosystem, and due to its inland character, large drainage area, and limited exchange of sea water with the Atlantic it is very sensitive to ongoing natural and anthropogenic (climate change, pollution, eutrophication, and overfishing) changes. The coastal zone is especially vulnerable, and in similarity to other regions of the Baltic Sea, exhibits little variety in the number of animal species, which is the result of eutrophication and the degradation of the environment. Despite this, it is considered among the marine habitats with the highest biological productivity. It plays an important ecological role by offering a variety of habitat types for many species, giving shelter to animals, and functioning as nursery areas and feeding grounds for many marine fishes and crustaceans.
In marine pelagic food webs, zooplankton plays a key role as an important link in energy transfer between primary producers and higher tier consumers, strongly influencing fish production. Zooplankton of the Gulf of Gdańsk typically consist of euryhaline and eurythermic taxa, among copepods mainly species from genera Acartia and Temora longicornis, as well as the less abundant, but ecologically important, Pseudocalanus acuspes. They are preferred prey items for commercially important fishes like sprat and herring as well as larval cod.
In order to properly asses the role of zooplankton in the marine food web, zooplankton secondary production and mortality rates need to be estimated. It is a useful tool to obtain knowledge of marine productivity, quantifying, behavior, distribution, migration patterns, and transfers between food web components [
1,
2,
3]. Many studies have focused on vital rates of copepods in the laboratory [
4,
5,
6,
7,
8,
9,
10], yet only few studies exist on its population dynamics in the field [
11,
12,
13,
14,
15,
16,
17].
The aim of this study was to describe the secondary production and mortality rates of three dominant calanoid copepod species—
Acartia spp.,
Temora longicornis, and
Pseudocalanus acuspes—in the southern Baltic Sea with relation to hydrographic water conditions. The data will be used for upgrading the copepod population model for the Baltic Sea [
10,
16,
18,
19,
20].
4. Discussion
The aim of our study was a description of seasonal and interannual patterns of secondary production and mortality rates for the main southern Baltic copepods. The main factors determining zooplankton production are temperature and food availability [
29]. Therefore, we decided to investigate seasonal and interannual fluctuations of secondary production of three copepod taxa:
Acartia spp.,
T. longicornis, and
P. acuspes in the southern Baltic. This is even more important in the context of increasing water temperatures observed in the Baltic related to global warming [
30]. Higher water temperature leads to shorter generation time and smaller body size of copepods, also causing individuals to reach reproductive age quicker, and causing rapid increases in density [
31]. However, different copepod species have their individual temperature optimums, at which their development is most optimal [
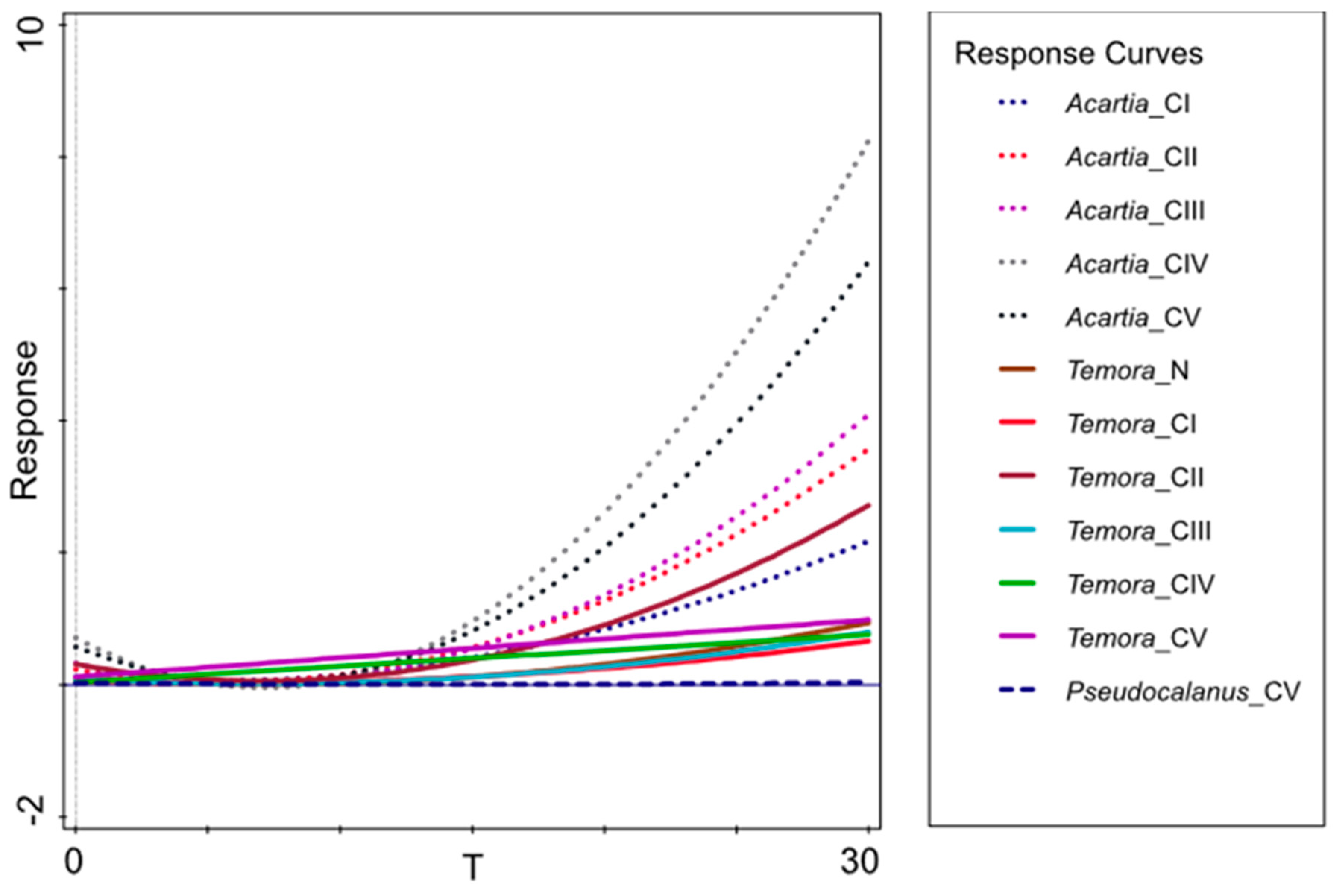
10]. Therefore, when estimating the rate of secondary production of these copepods, we used a
Di function that takes into account the individual temperature optimums. We are also aware that accurate estimates, most approximate to the natural state of the secondary production based on mathematical expressions, require a method that combines a variety of factors. Because of that our estimations do not fully reflect actual, real as in the natural environment, secondary production values. However, they allow for an approximation of these values and enable recognition of trends or anomalies occurring in the studied ecosystem.
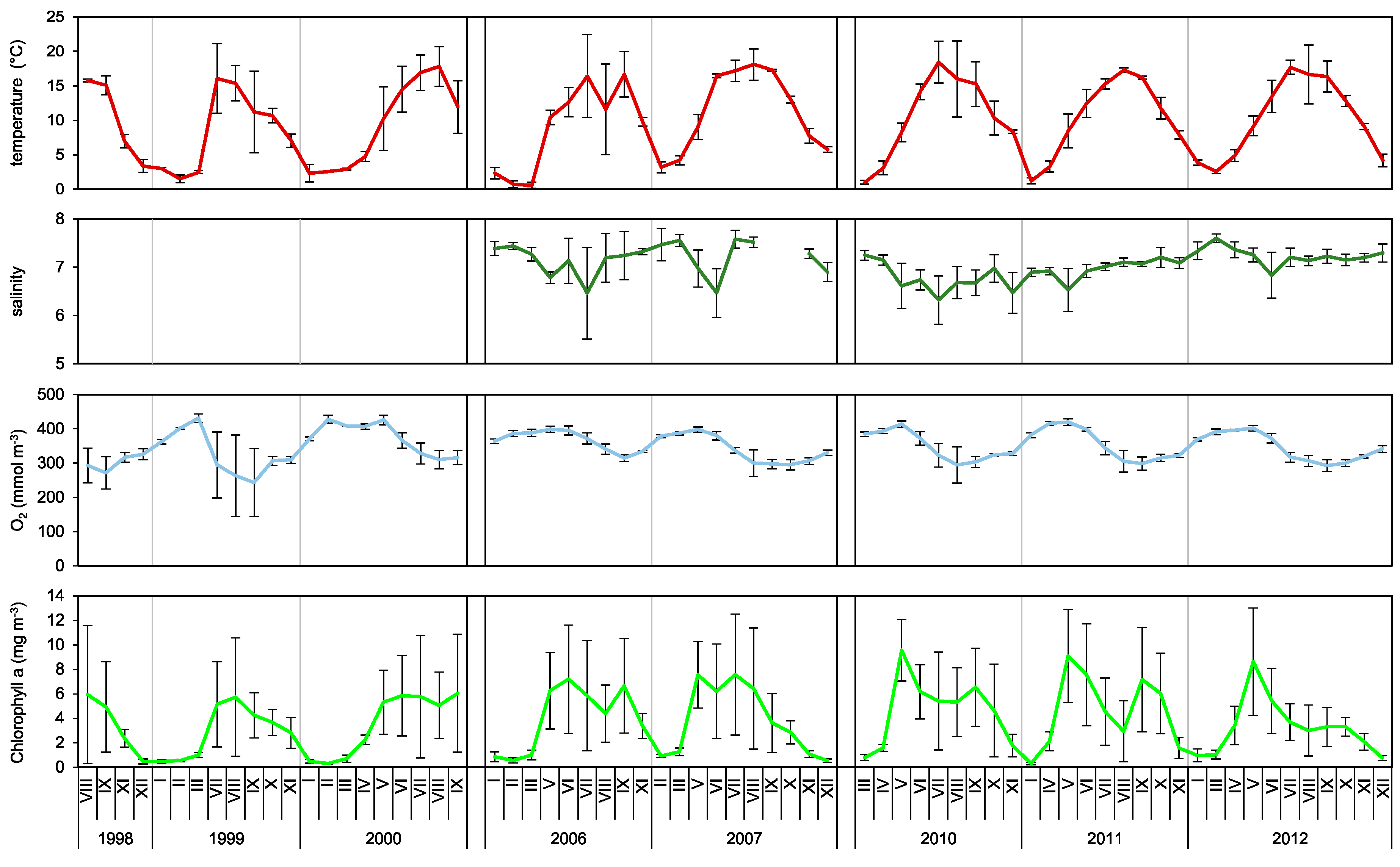
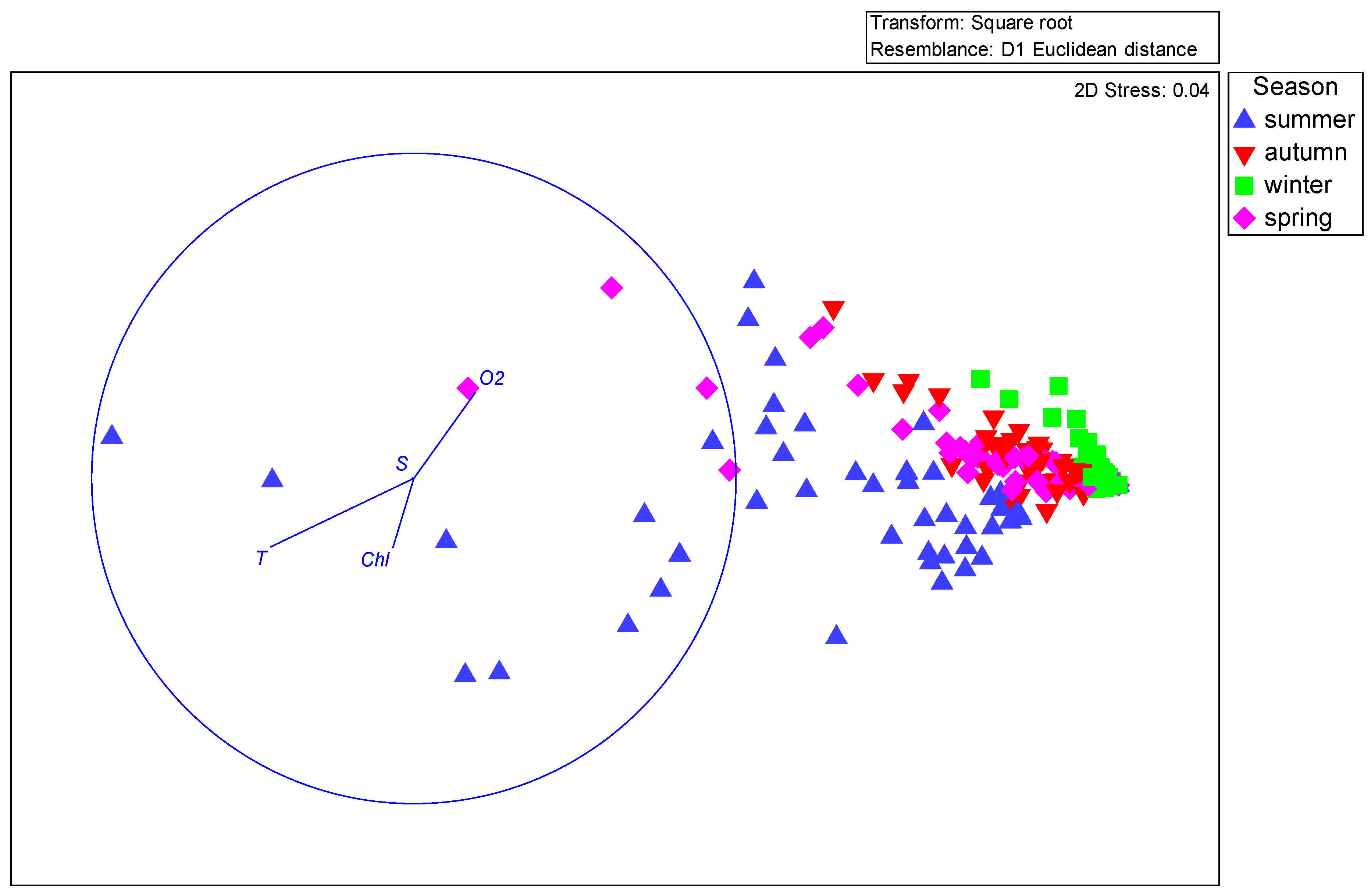
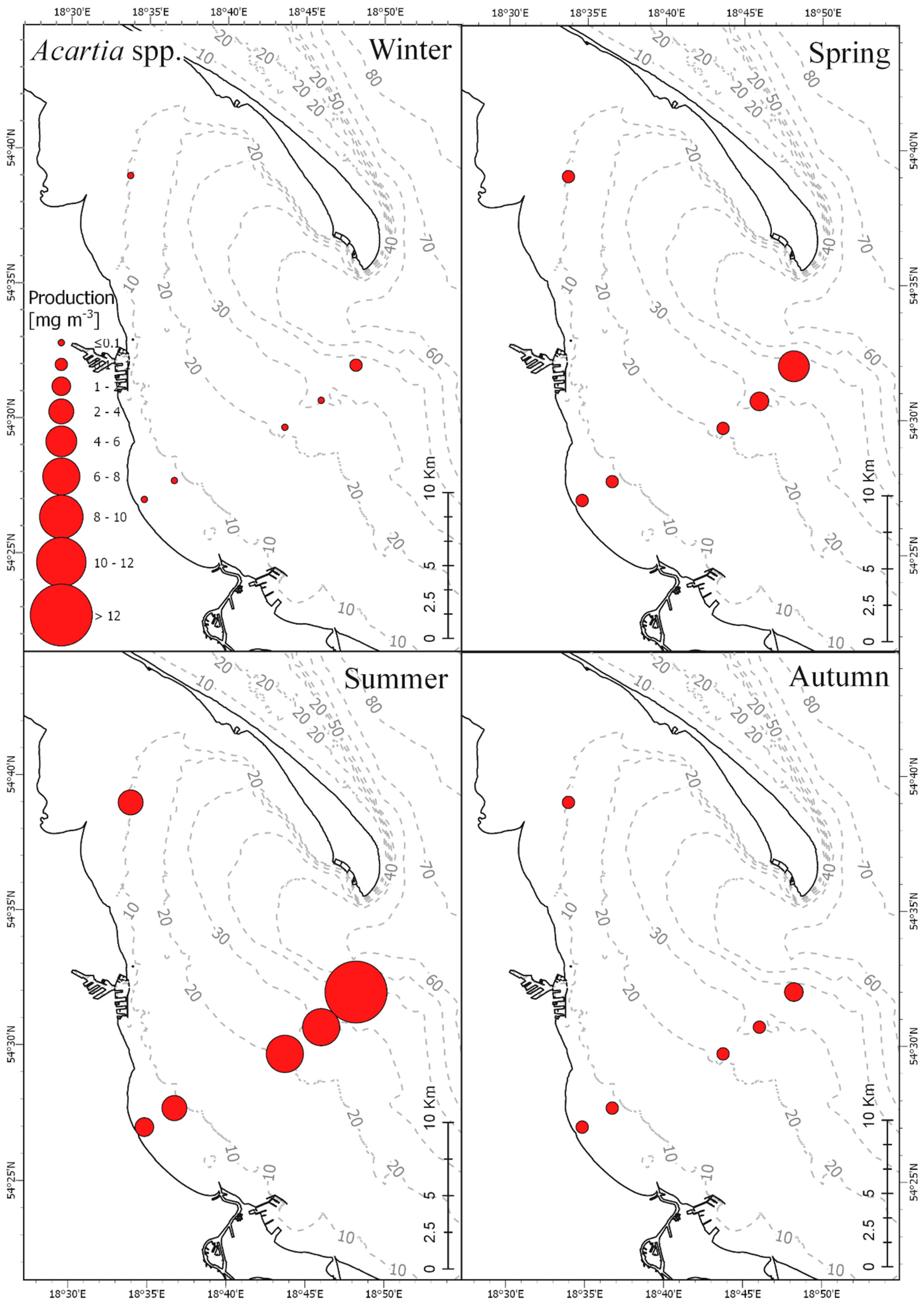
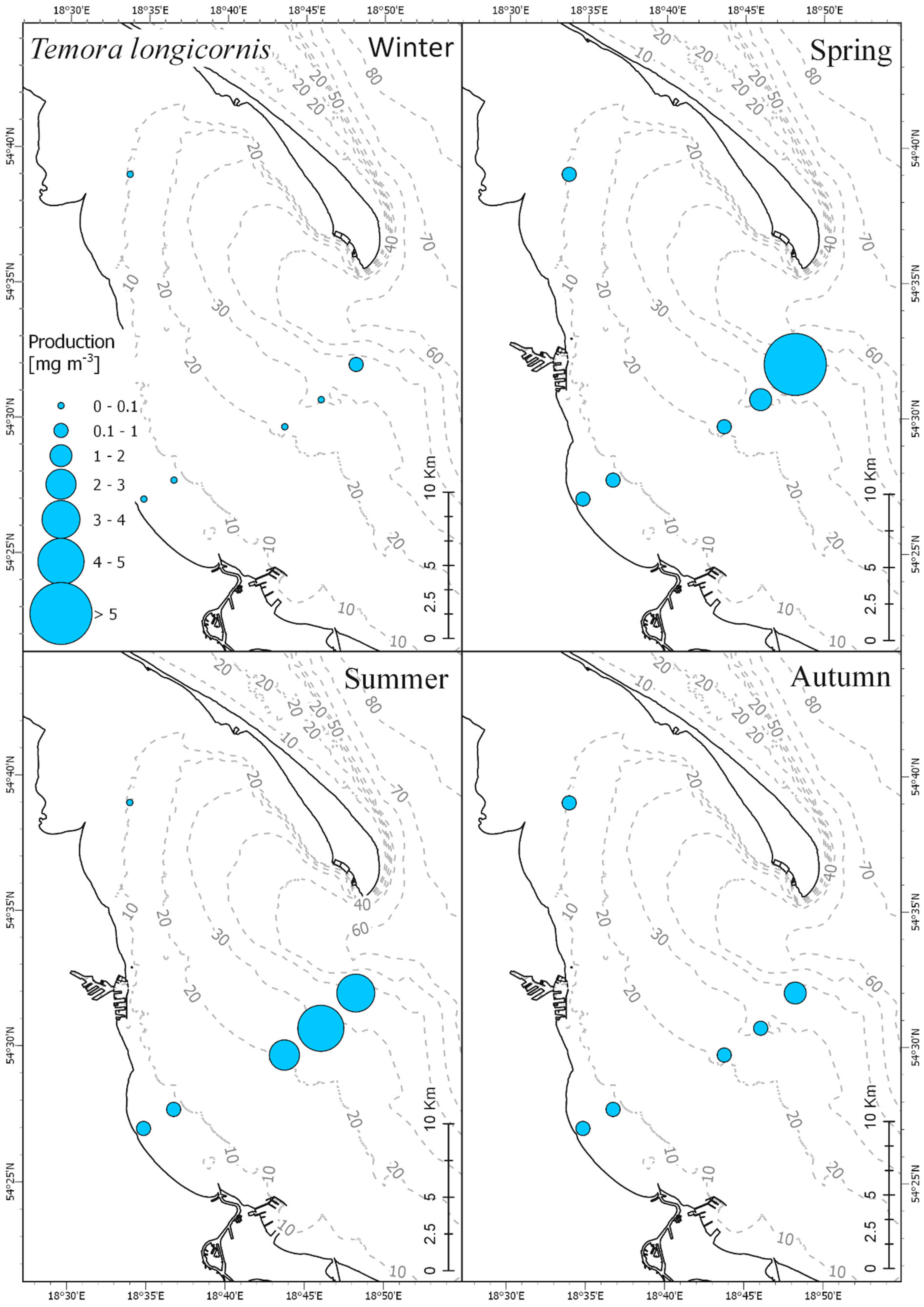
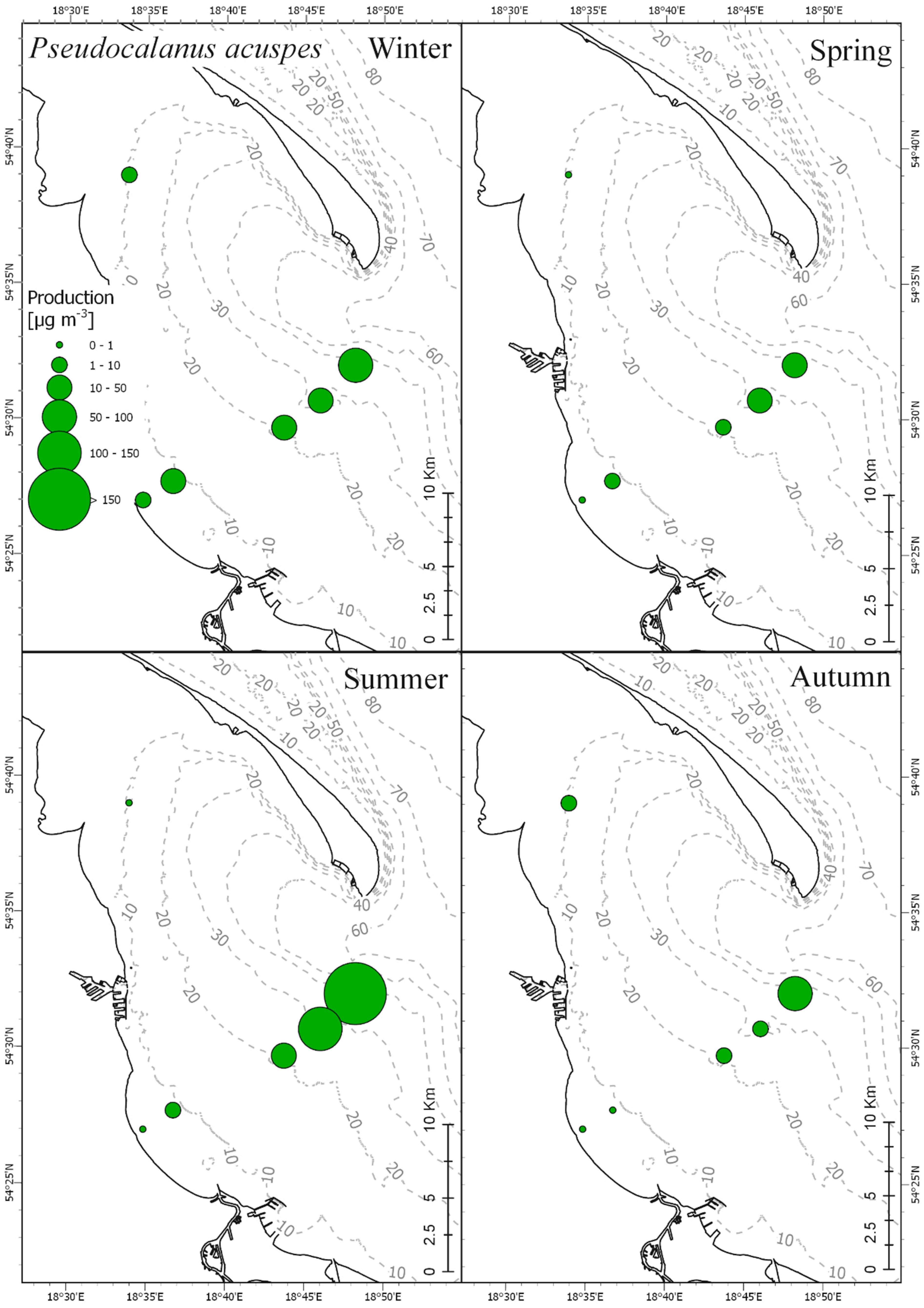
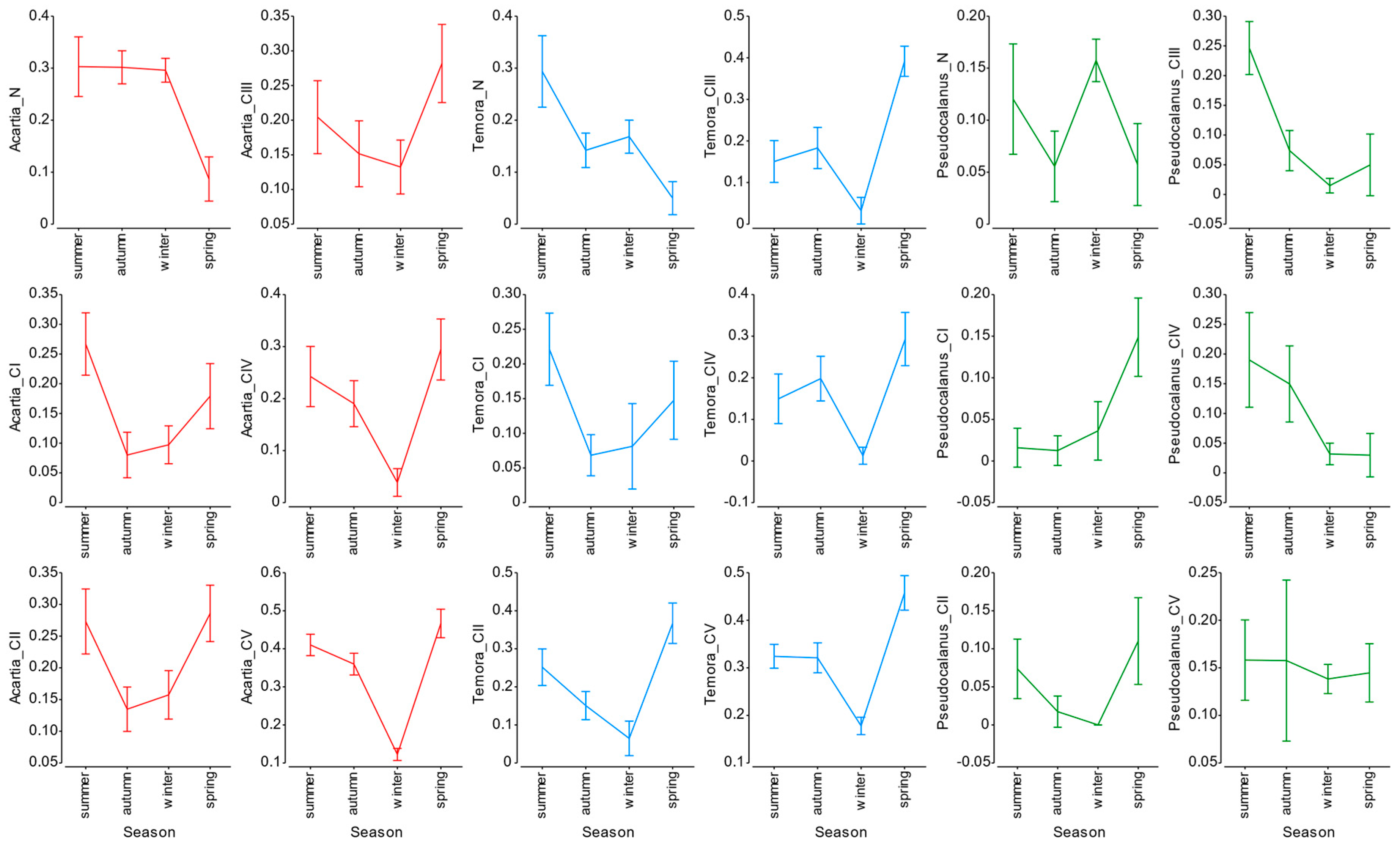
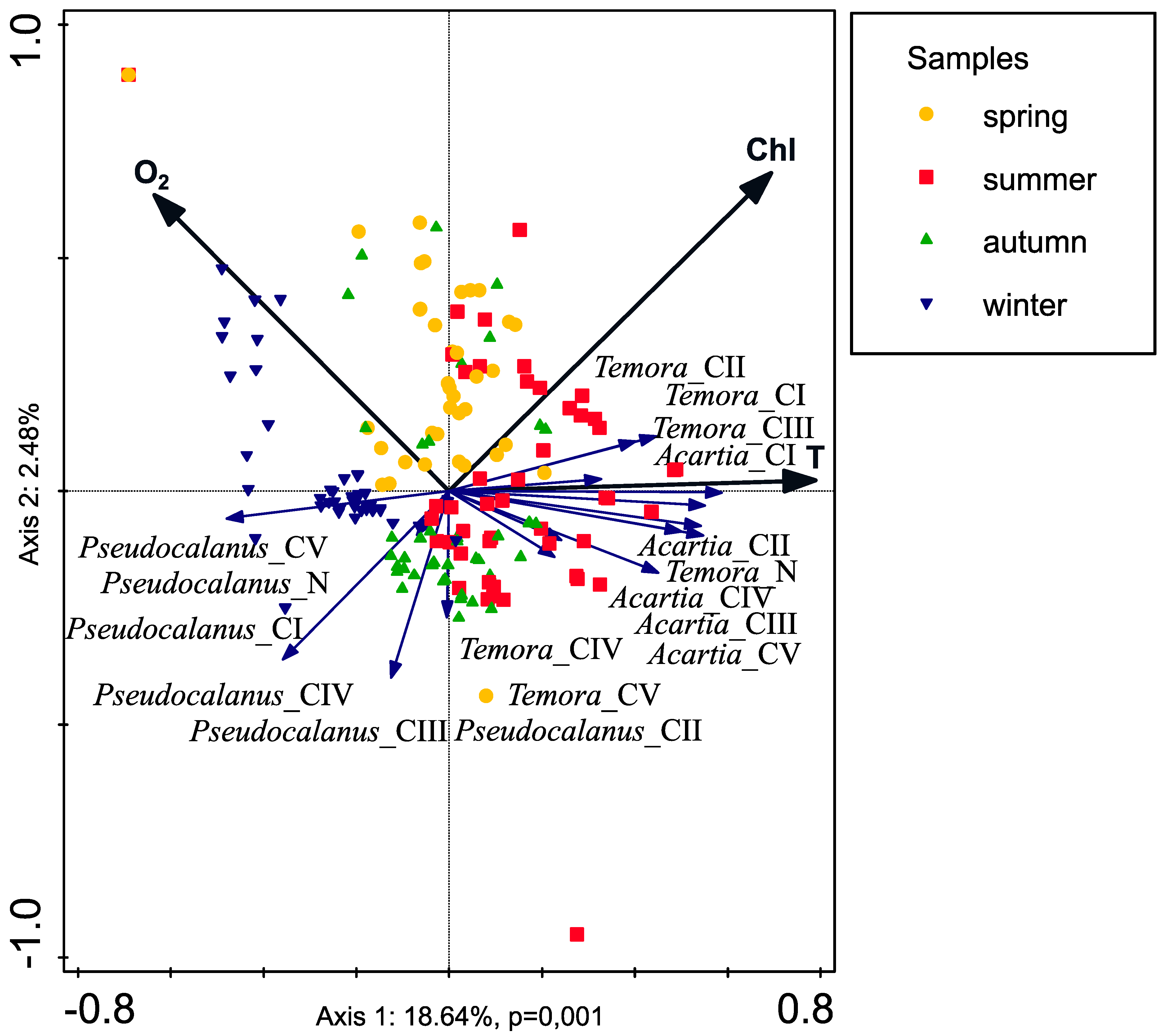
Results obtained in our research showed clear correlation between seasonal production fluctuations in the Gulf of Gdańsk and the hydrological conditions, mainly water temperature (
Figure 21). The highest correlation was recorded during summer, mainly for the young copepodite stages of
Acartia spp. as well as nauplii and copepodites of
T. longicornis. This is consistent with research carried out by Koski et al. [
32] in the North Sea, which also indicates that the production coefficient is significantly positively correlated with the average water temperature. However, research from the Western Scheldt Estuary [
33], showed that neither the biomass nor the secondary copepod production was associated with chlorophyll concentration, and the temperature seemed to have a significant impact only on the predominance of certain copepods. In contrary to those two taxa,
P. acuspes showed mostly negative correlation of secondary production with temperature. This species in the central Baltic is associated with the deeper, more saline, colder water layer [
34,
35,
36]. This was visible in high mean values of secondary production of nauplii and older copepodites (CIV and CV) noted during winter seasons (
Figure 8). We can therefore clearly state that, similarly to other water basins, water temperature is one of the main factors controlling not only biomass and abundance [
17,
37] but also secondary production of main copepod taxa in the Gulf of Gdańsk. Temperature was responsible for 11.8% of variability observed in RDA. In addition to temperature, variability of secondary production was also to some extent explained by the concentration of dissolved oxygen (5%) and chlorophyll
a (4.5%) (
Table 6).
Our results indicate that the water salinity does not have a statistically significant effect on abundance of copepodite developmental stages (
Table 6). There was also no correlation for both production and mortality rates. These results differed from literature data from other brackish [
38] or low salinity water bodies [
36,
39]. For example, in the Lake Waihola both elevated water temperature and salinity affected population dynamics of
Boeckella hamata. Additionally, higher temperature and salinity, which were related to the increase of egg production, were offset by the higher mortality rates of reproductive females [
38]. Nagaraj, on the other hand, described that the combination of temperature and salinity had a greater impact on mortality of
Eurytemor affinis than each of these factors separately [
40]. In the German Southern Bay (North Sea) [
36], due to the high summer temperature, higher salinity, and sufficient food concentration,
Pseudocalanus elongatus have three to four generations per year; similarly in the northern part of the North Sea three generations are usually described [
41]. While Evans [
42] described four to six generations of that species in coastal waters of Northumberland (also North Sea). However, in the Southern Baltic, due to lower salinity only one full generation per year for
P. acuspes is observed [
43]. Such a discrepancy of results is a result of the females’ physiological abilities, due to slower growth and differences in the body size which are related to lower salinity. This is a well-known phenomenon concerning many organisms from the Baltic Sea, including copepods;
T. longicornis and
P. acuspes occurring in the North Sea reach significantly larger body sizes than those recorded in the Baltic Sea.
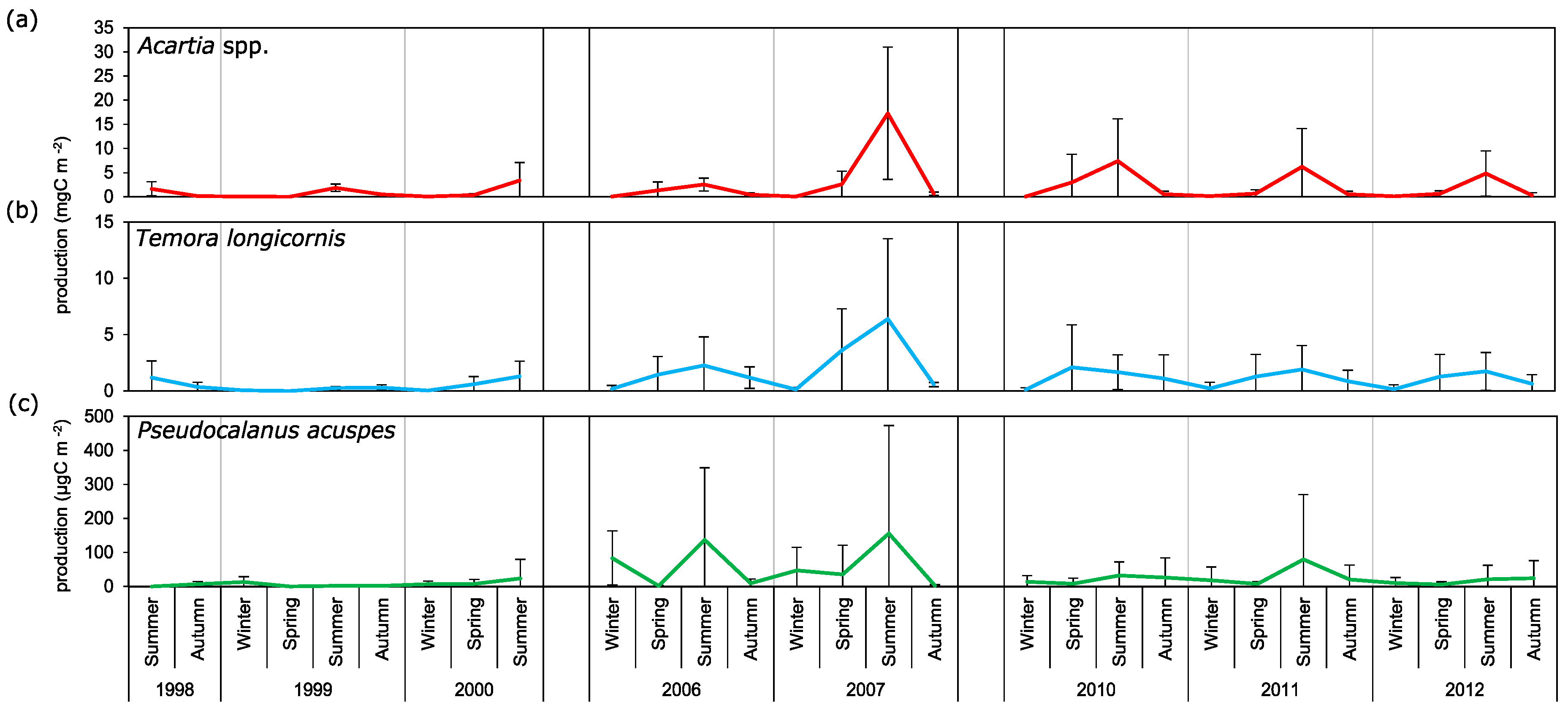
Comparison of the estimated values of secondary production of crustaceans from other regions shows that that copepod production in the Gulf of Gdańsk is relatively low. The maximum estimated average value of
P. acuspes was 0.2 mgC m
−2 day
−1 in the summer of 2007, while Renz [
44] described it as 26.2 mg C m
−2 day
−1 in June. Such a large difference in the obtained values is probably due to differences in hydrological factors leading to differences in metabolic rates [
36]. Renz and Hirche [
36] have shown that the rate of North Sea
P. elongatus population development is ~3–5 times higher and the growth rate is up to 10 times higher than the
P. acuspes population from the Baltic Sea, which translates directly into the higher production of
P. elongatus. Results reported by Fransz [
45] from the North Sea show that the secondary production of
T. longicornis and
Acartia clausi fluctuated between 22 and 16 mgC m
−2 between May and September. In the brackish waters of the Western Scheldt Estuary, the estimated maximum average secondary production of
Acartia tonsa oscillated around 25 mgC m
−3 in August; the second peak was recorded in September with the value of 8 mgC m
−3. The presence of maxima of secondary production in these months is consistent with the seasonality of production observed for these species from genus Acartia in the Gulf of Gdańsk. Production values, as in the case of
P. acuspes, were, however, lower than those recorded in the North Sea. Differences in the value of secondary production can also be caused by the use of different calculation methods and wet mass converters for the copepod from various sources of literature.
In the perspective of the observed progressing warming of the Baltic Sea, which is particularly noticeable in its northern region (the air temperature in the spring increased by ~1.5 °C over the period of 1871 to 2011 [
46]. The ecosystem of the southern Baltic, which is much more productive and biodiverse, is more susceptible to the negative effects of such changes. Due to the currently observed restructuring of unicellular plankton and the shift of phenological phases [
47], further comprehensive research on biological production in ecosystems is needed, combining the research on primary phytoplankton production and the production of zooplankton. Observed progressing delaying of the maximum of secondary production in relation to spring bloom of phytoplankton may lead to serious consequences for the whole organic production and higher trophic levels in the ecosystem.
Zooplankton mortality estimates are still not a well-developed parameter in determining population dynamics. Therefore, selecting a proper methodology to describe this phenomenon can be challenging, and the main uncertainty results from inherent difficulties in measuring this process in the natural environment [
48].
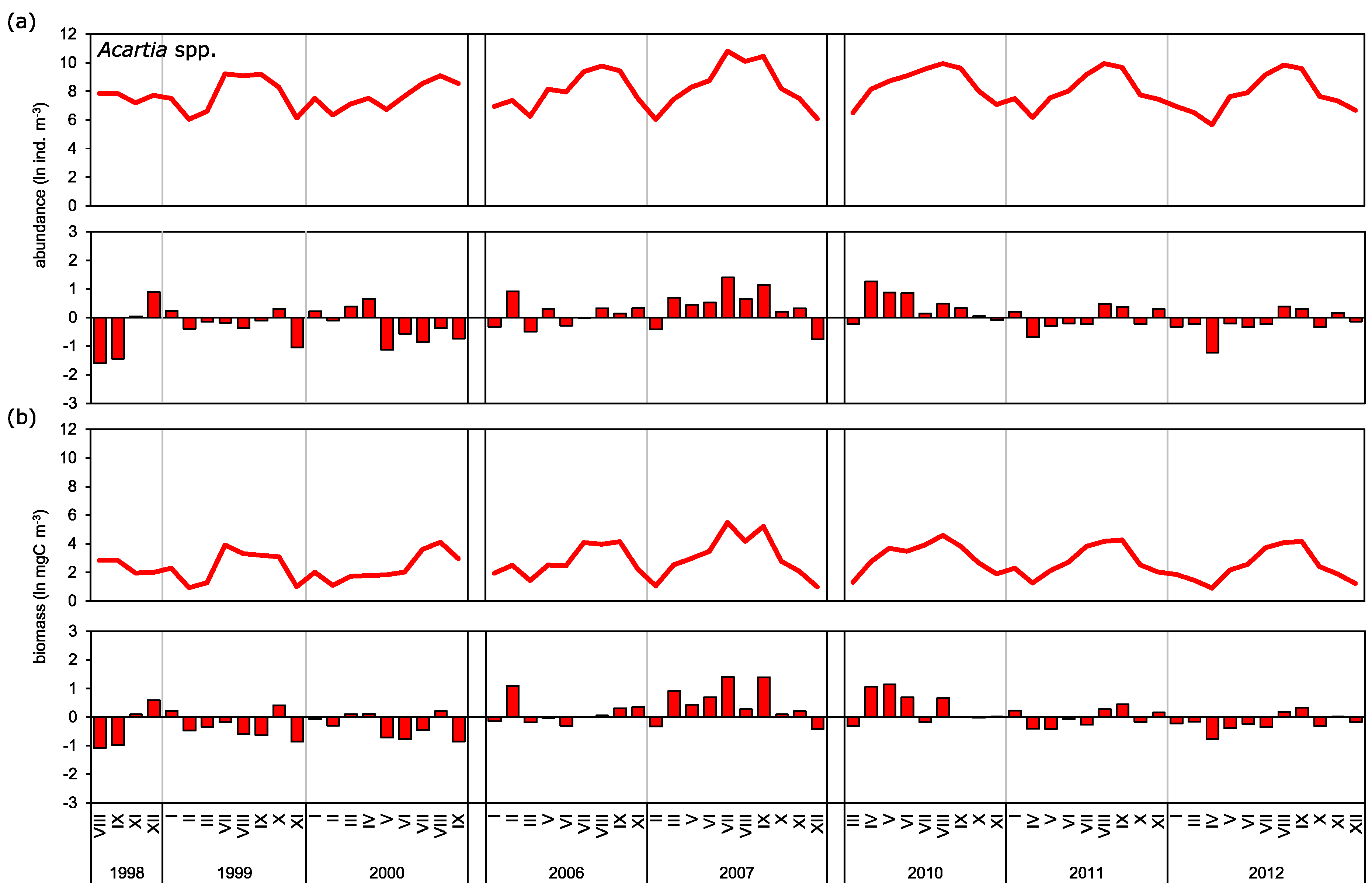
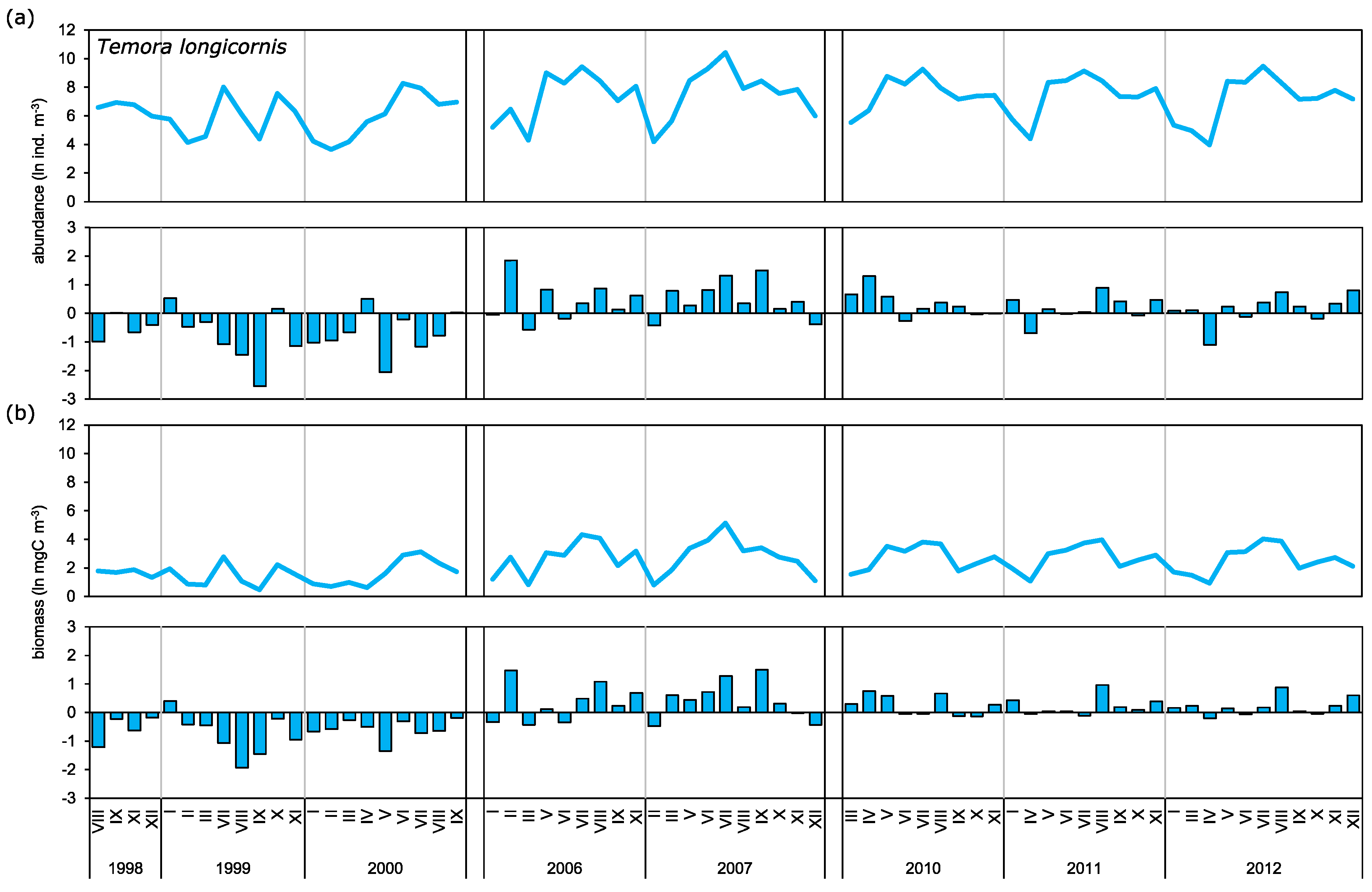
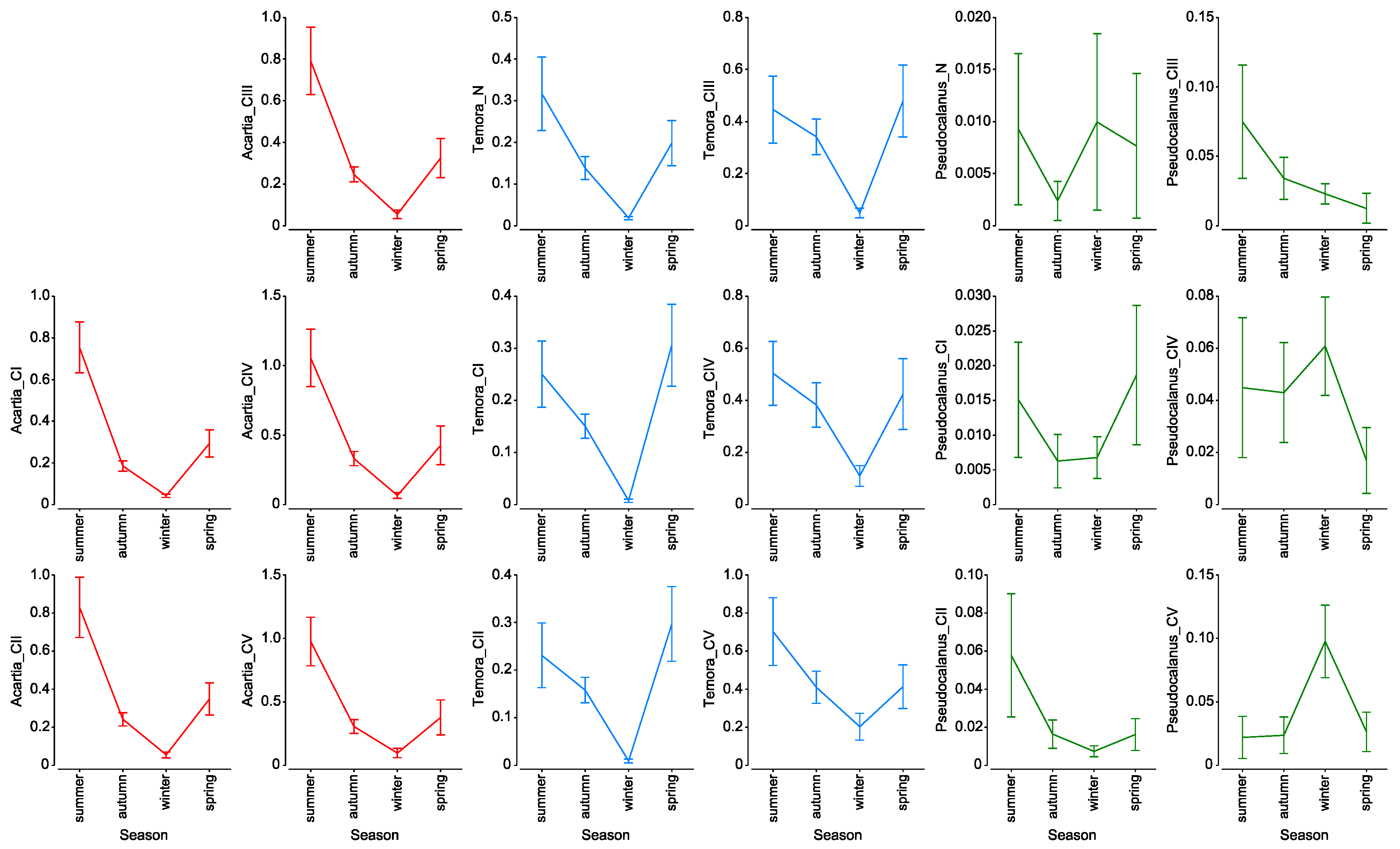
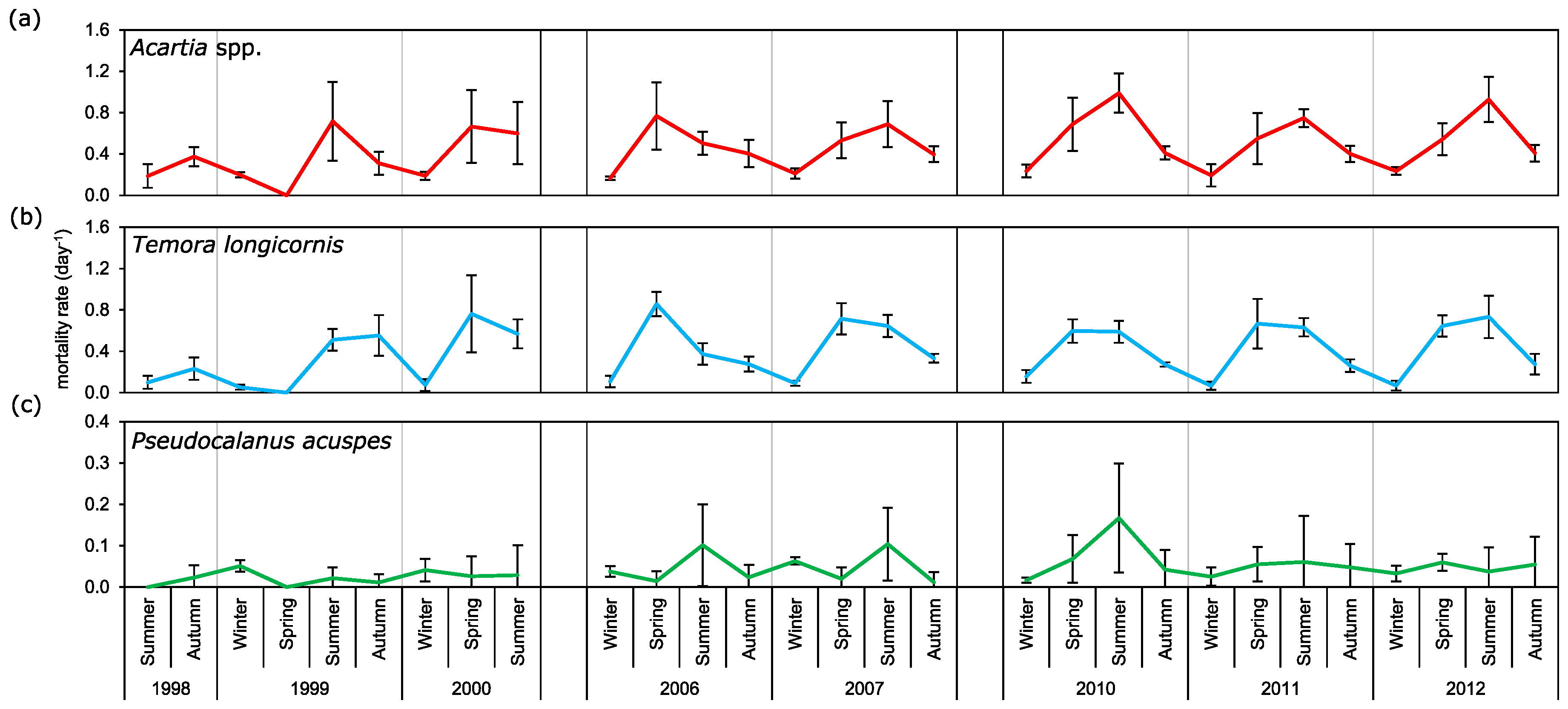
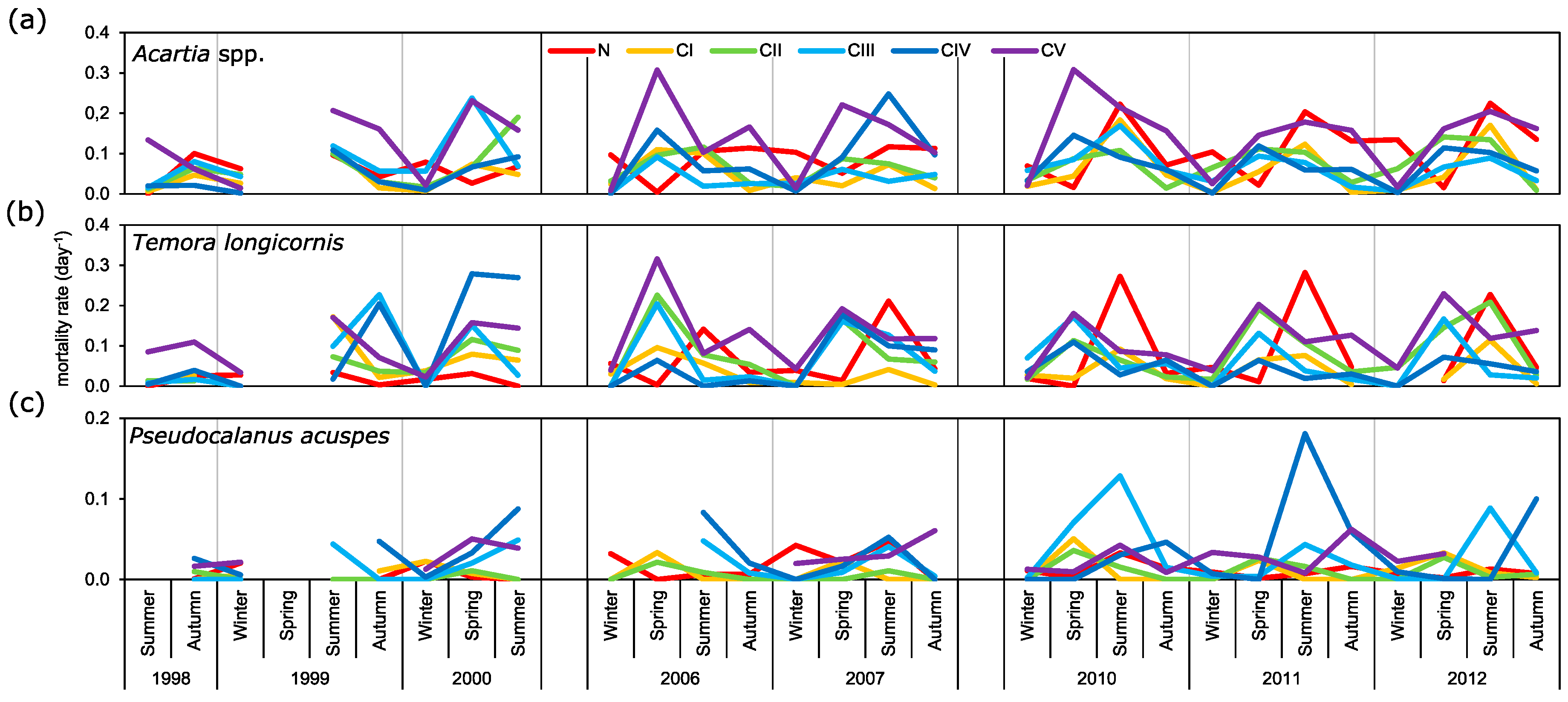
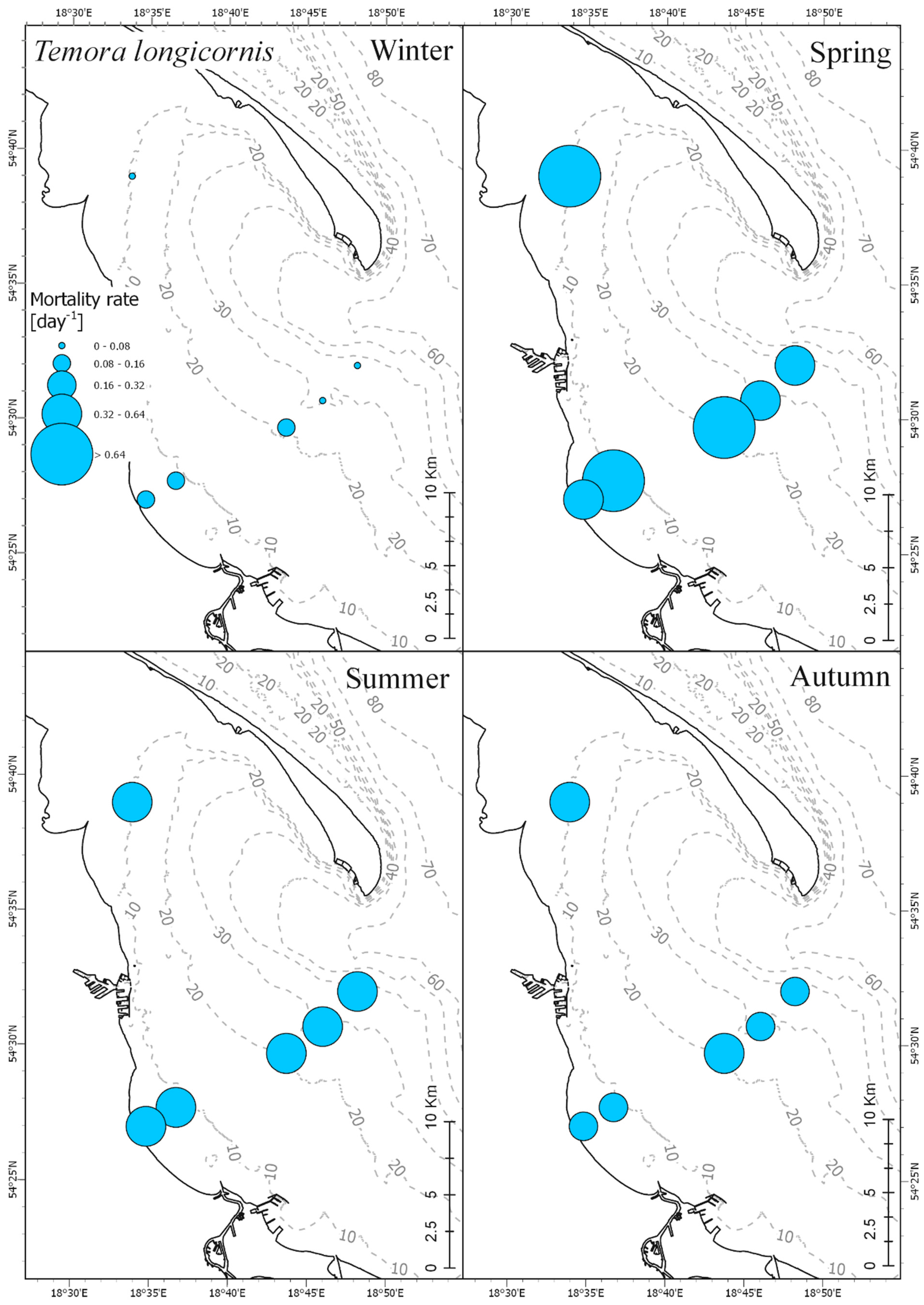
It is widely accepted that the daily copepod mortality decreases with consecutive developmental stages or with an increase in size [
49,
50]. Based on these assumptions we wanted to describe mortality rates of main copepod taxa from the Gulf of Gdańsk at particular seasons of the year. The results obtained for
Acartia spp. and
T. longicornis, however, show differences in mortality for particular stages from that described by the above-mentioned authors. The highest mortality rate for
Acartia spp. was observed for the oldest copepodites (CV) with maximum values noted in spring: ~0.3 day
−1. For
T. longicornis, high rates were observed for CIII, CIV, and CV, within a range of 0.15 day
−1 to 0.28 day
−1. High variability in mortality estimates between copepod species and developmental stages in both spatial and seasonal distribution were also observed by other authors [
15,
51,
52].
In our research we observed cyclical changes in mortality, with the peak falling in the spring and summer season. Maud et al. [
53], on the other hand, recorded the highest mortality rates in summer and autumn, with the lowest—as in our research—in winter. Differences in seasonality of mortality in different regions may result from differences in the main cause of mortality. Mortality of copepods can be caused by predatory (consumptive) or physicochemical and biological factors as factors causing nonconsumptive mortality. Consumptive mortality may be associated with the abundance of predators [
54], and is described in the literature as usually occurring in the autumn season, when the abundance of predators is the highest. This type of mortality described for
Calanus helgolandicus constituted an average of 89% of the total mortality for this species [
53]. In the Baltic Sea, copepods are a valuable source of food for the commercially important fish species sprat and herring, but jellyfish can also have a significant predatory impact. The gelatinous zooplankton was the main cause of mortality variability observed in deep coastal sampling station located near the southwest of Plymouth, UK [
55]. Nonconsumer mortality may result from death caused by age [
56], diseases, and parasitism [
57], exposure to environmental pollution [
58], and physiological stress [
59]. Field and laboratory studies show that nonconsumptive factors can account for 25–33% of the total death rate among adult copepods [
60].
The species from genus Acartia had the highest abundance and biomass among the three investigated taxa, while
P. acuspes was far less abundant than the other two taxa. Such proportions between these taxa are quite typical for the coastal region of the southern Baltic.
P. acuspes tends to dominate offshore areas of the Baltic Sea, is much less abundant in the coastal zones, and it is rarely present above the thermocline, especially during the warm season [
61,
62].
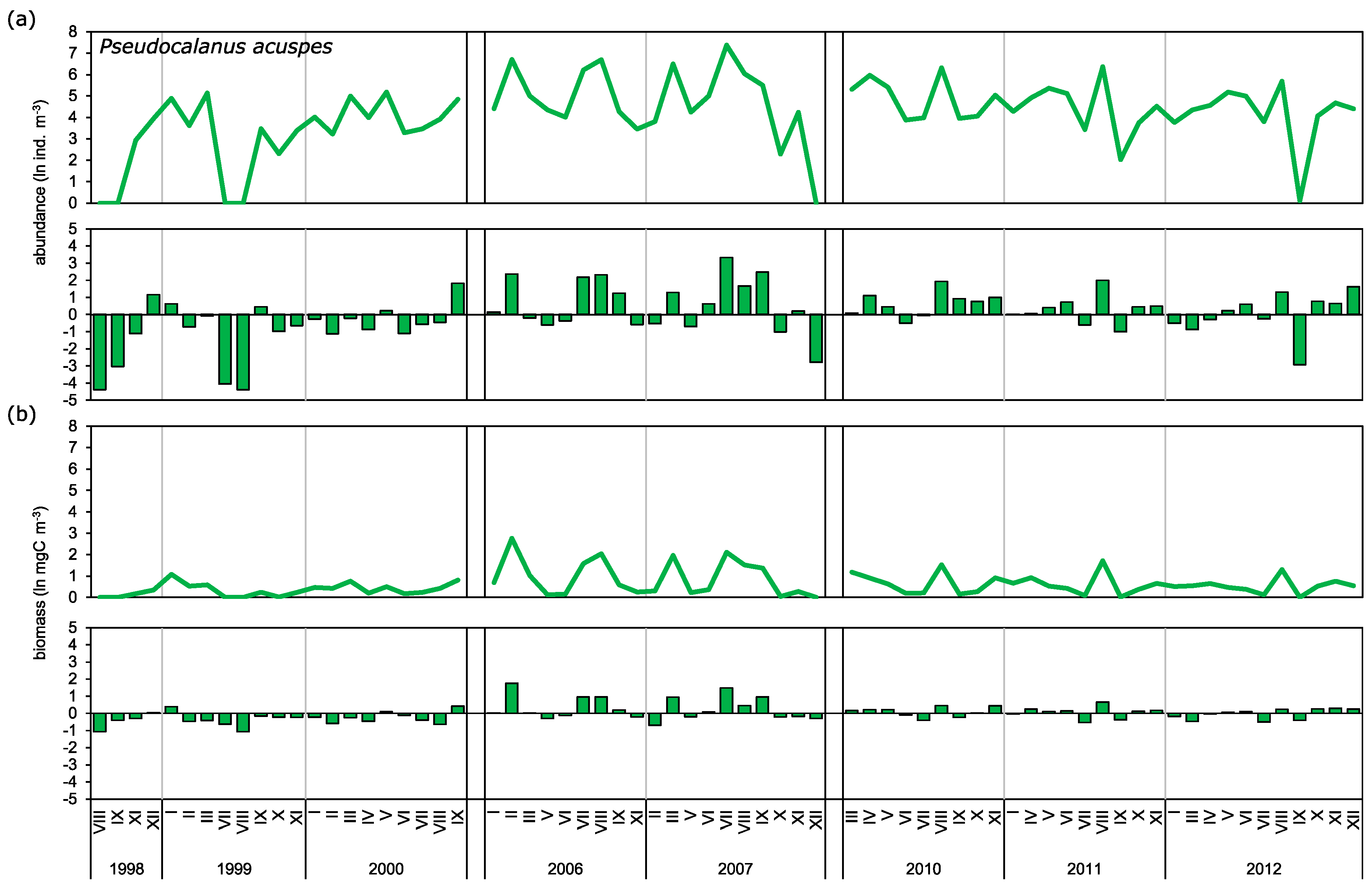
Observed long-term biomass means for
Acartia spp. were in a similar range as those reported for this region by Möllmann [
30], while for
T. longicornis, and especially
P. acuspes, they were much lower. This was probably due to our sampling stations being located mostly in the inner, coastal part of the Gulf of Gdańsk. Möllmann [
30] also showed mostly negative anomalies for both
T. longicornis and
P. acuspes as well as the mostly neutral anomaly of
Acartia spp. during the late 1990s. This is consistent with our findings, which also showed strong positive anomalies for those taxa during the first decade of the 2000s.
Obtained data show seasonal fluctuations of copepod abundance, biomass, production, and mortality rates, as well the correlation with environmental variables influencing their development and mortality. The results of this work will be useful for future extended evaluation of copepod production with relation to environmental variables in brackish waters.

 ,
, 





















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}