
Worry Modifies the Relationship between Locus Coeruleus Activity and Emotional Mnemonic Discrimination

 , , , ,
, , , , 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Participants
2.2. Measurements and Tests
2.2.1. Neuropsychological Testing and Questionnaires
2.2.2. Worry Questionnaire
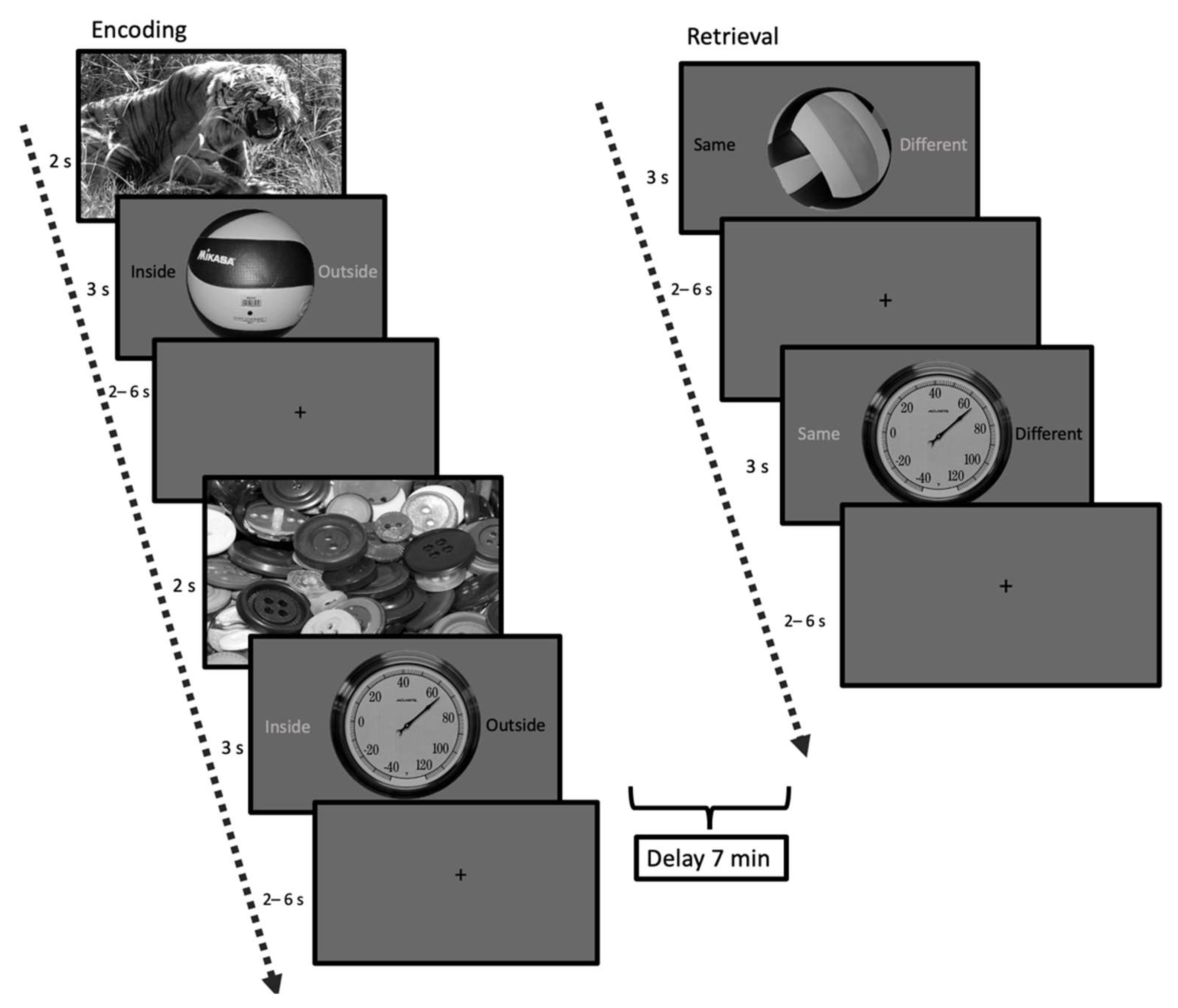
2.2.3. Emotional Mnemonic Discrimination Memory Task
2.3. Imaging
7T Imaging Parameters
2.4. Preprocessing of MRI Data
2.4.1. Anatomical Preprocessing
2.4.2. Preprocessing of BOLD fMRI Data
2.4.3. ROI Value Extraction
2.5. Statistical Analyses
- Memory performance ~ age + sex + depression score + worry score
- 2.
- Memory performance ~ age + sex + worry score + ROI contrast activity
- 3.
- Memory performance ~ age + sex + worry score × ROI contrast activity
3. Results
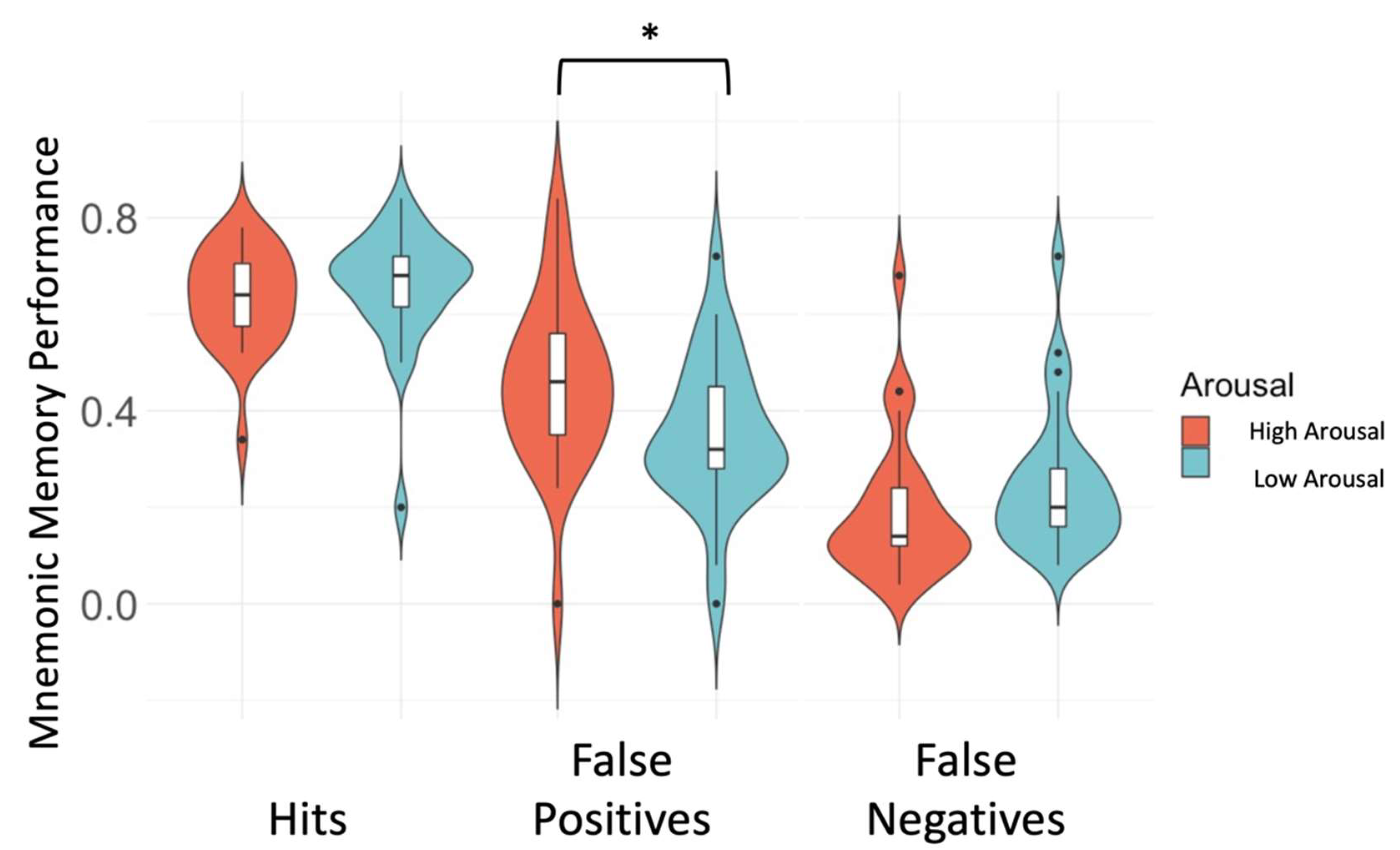
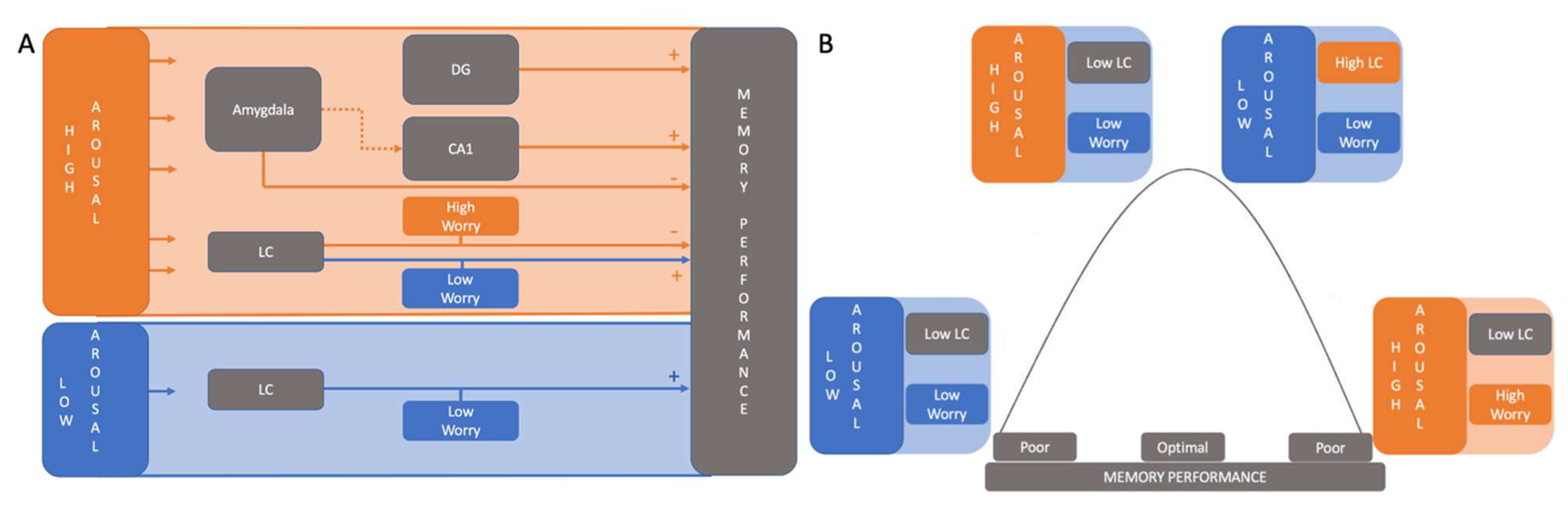
3.1. High Arousal Modulates Mnemonic Discrimination Performance
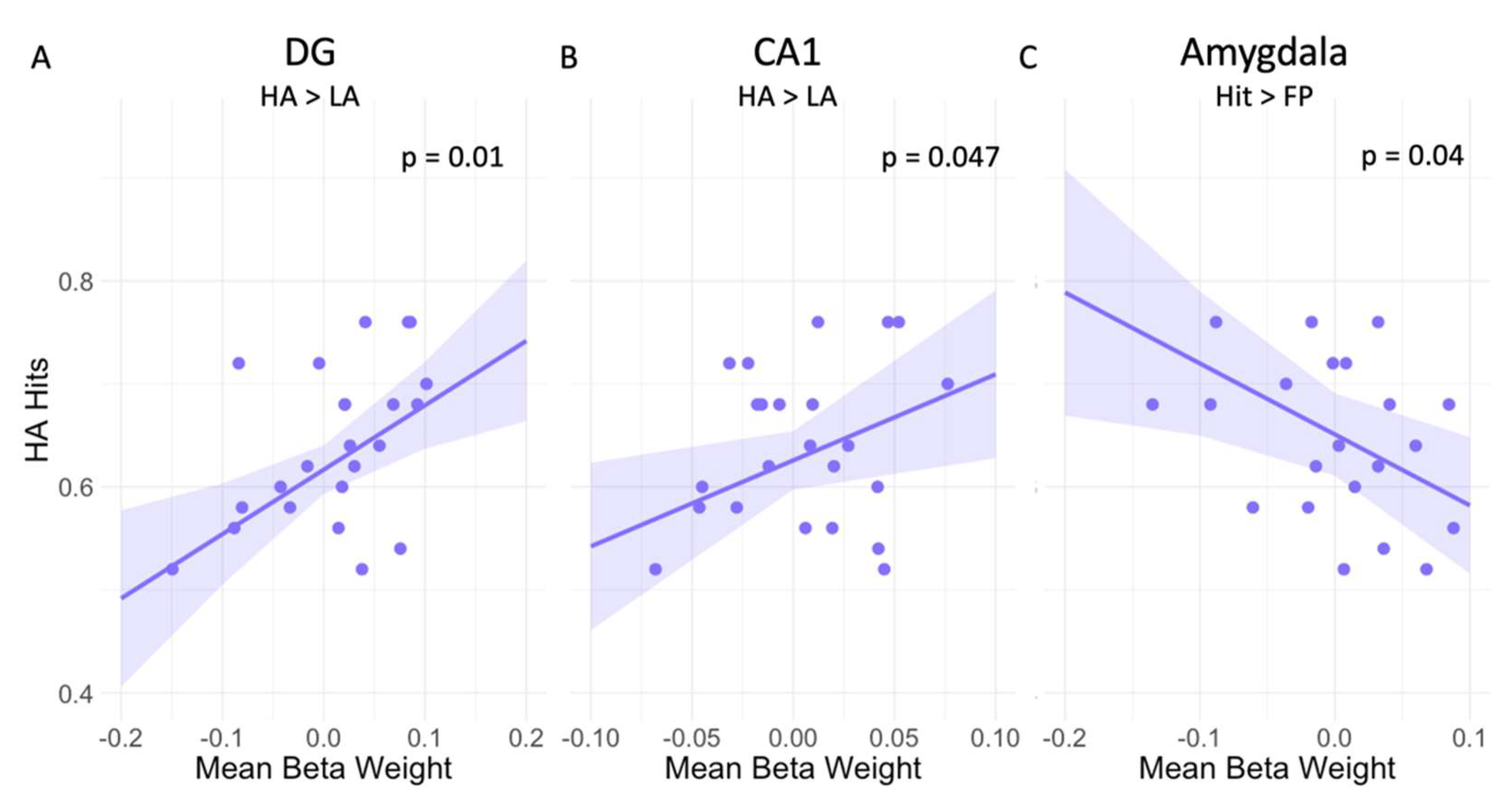
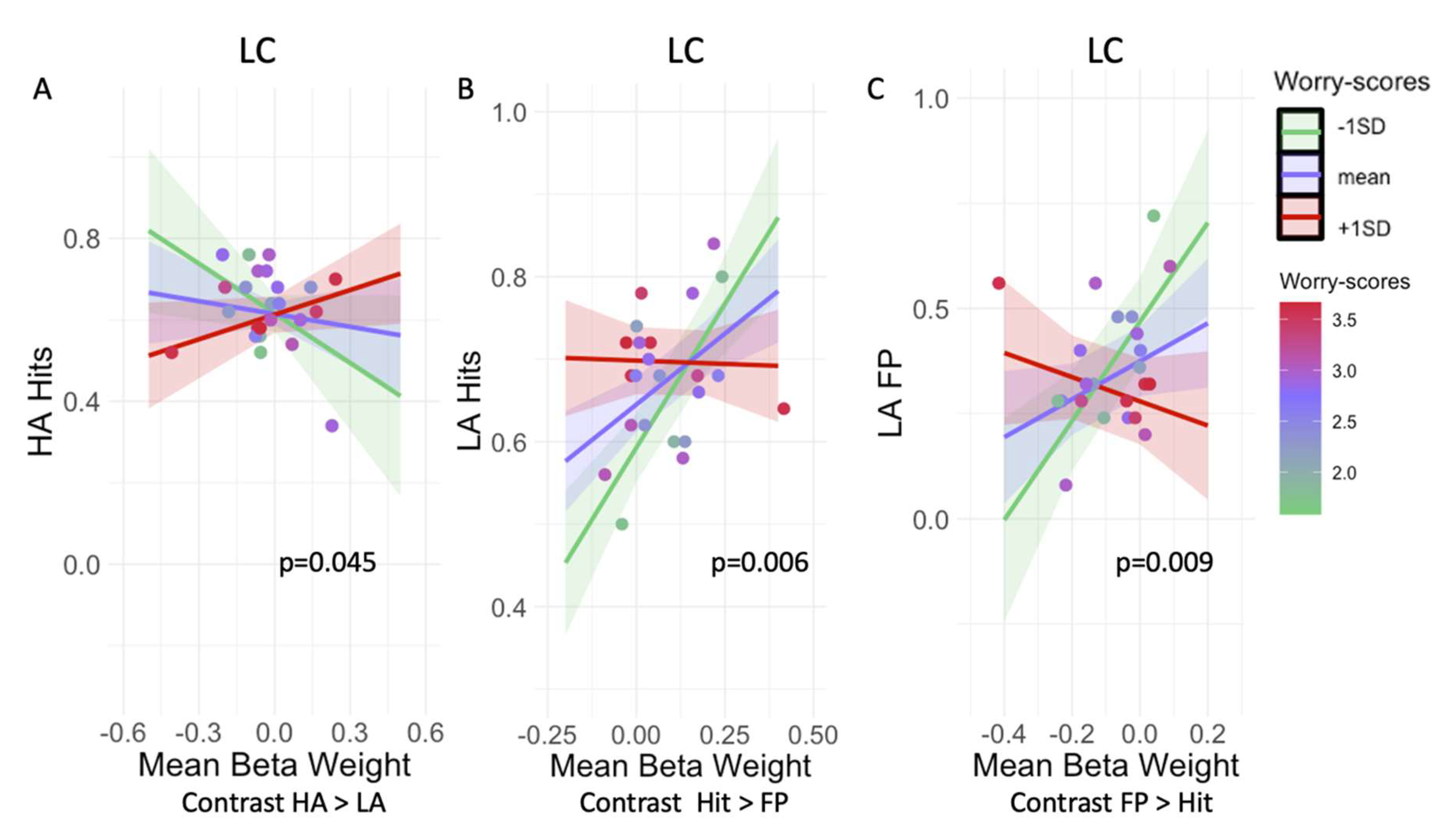
3.2. Worry Modulates the Relationship between LC Activation and Mnemonic Discrimination Performance
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Roozendaal, B.; McGaugh, J.L. Memory modulation. Behav. Neurosci. 2011, 125, 797–824. [Google Scholar] [CrossRef] [PubMed]
- McGaugh, J.L. The amygdala modulates the consolidation of memories of emotionally arousing experiences. Annu. Rev. Neurosci. 2004, 27, 1–28. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Segal, S.K.; Cahill, L. Endogenous noradrenergic activation and memory for emotional material in men and women. Psychoneuroendocrinology 2009, 34, 1263–1271. [Google Scholar] [CrossRef] [PubMed]
- Segal, S.K.; Stark, S.M.; Kattan, D.; Stark, C.E.; Yassa, M.A. Norepinephrine-mediated emotional arousal facilitates subsequent pattern separation. Neurobiol. Learn. Mem. 2012, 97, 465–469. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aston-Jones, G.; Cohen, J.D. An integrative theory of locus coeruleus-norepinephrine function: Adaptive gain and optimal performance. Annu. Rev. Neurosci. 2005, 28, 403–450. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Berridge, C.W.; Waterhouse, B.D. The locus coeruleus–noradrenergic system: Modulation of behavioral state and state-dependent cognitive processes. Brain Res. Rev. 2003, 42, 33–84. [Google Scholar] [CrossRef]
- Jacobs, H.I.; Wiese, S.; van de Ven, V.; Gronenschild, E.H.; Verhey, F.R.; Matthews, P.M. Relevance of parahippocampal-locus coeruleus connectivity to memory in early dementia. Neurobiol. Aging 2015, 36, 618–626. [Google Scholar] [CrossRef] [PubMed]
- Mather, M.; Harley, C.W. The Locus Coeruleus: Essential for Maintaining Cognitive Function and the Aging Brain. Trends Cogn. Sci. 2016, 20, 214–226. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Winklewski, P.J.; Radkowski, M.; Wszedybyl-Winklewska, M.; Demkow, U. Stress Response, Brain Noradrenergic System and Cognition. In Respiratory System Diseases; Springer: Berlin/Heidelberg, Germany, 2017. [Google Scholar]
- Aston-Jones, G.; Cohen, J.D. Adaptive gain and the role of the locus coeruleus-norepinephrine system in optimal performance. J. Comp. Neurol. 2005, 493, 99–110. [Google Scholar] [CrossRef] [PubMed]
- Sara, S.J. The locus coeruleus and noradrenergic modulation of cognition. Nat. Rev. Neurosci. 2009, 10, 211–223. [Google Scholar] [CrossRef] [PubMed]
- Harley, C. Noradrenergic and locus coeruleus modulation of the perforant path-evoked potential in rat dentate gyrus supports a role for the locus coeruleus in attentional and memorial processes. Neurobiology of the Locus Coeruleus. Prog. Brain Res. 1991, 88, 307–321. [Google Scholar] [PubMed]
- Harley, C.W. A role for norepinephrine in arousal, emotion and learning?: Limbic modulation by norepinephrine and the Kety hypothesis. Prog. Neuro-Psychopharmacol. Biol. Psychiatry 1987, 11, 419–458. [Google Scholar] [CrossRef]
- Leal, S.L.; Tighe, S.K.; Jones, C.K.; Yassa, M.A. Pattern separation of emotional information in hippocampal dentate and CA3. Hippocampus 2014, 24, 1146–1155. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leal, S.L.; Tighe, S.K.; Yassa, M.A. Asymmetric effects of emotion on mnemonic interference. Neurobiol. Learn. Mem. 2014, 111, 41–48. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yassa, M.A.; Stark, C.E. Pattern separation in the hippocampus. Trends Neurosci. 2011, 34, 515–525. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stark, S.M.; Kirwan, C.B.; Stark, C.E.L. Mnemonic Similarity Task: A Tool for Assessing Hippocampal Integrity. Trends Cogn. Sci. 2019, 23, 938–951. [Google Scholar] [CrossRef] [PubMed]
- Leal, S.L.; Noche, J.A.; Murray, E.A.; Yassa, M.A. Age-related individual variability in memory performance is associated with amygdala-hippocampal circuit function and emotional pattern separation. Neurobiol. Aging 2017, 49, 9–19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grella, S.L.; Neil, J.M.; Edison, H.T.; Strong, V.D.; Odintsova, I.V.; Walling, S.G.; Martin, G.M.; Marrone, D.F.; Harley, C.W. Locus Coeruleus Phasic, But Not Tonic, Activation Initiates Global Remapping in a Familiar Environment. J. Neurosci. 2019, 39, 445–455. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dahl, M.J.; Mather, M.; Düzel, S.; Bodammer, N.C.; Lindenberger, U.; Kühn, S.; Werkle-Bergner, M. Rostral locus coeruleus integrity is associated with better memory performance in older adults. Nat. Hum. Behav. 2019, 3, 1203–1214. [Google Scholar] [CrossRef] [PubMed]
- Hämmerer, D.; Callaghan, M.F.; Hopkins, A.; Kosciessa, J.; Betts, M.; Cardenas-Blanco, A.; Kanowski, M.; Weiskopf, N.; Dayan, P.; Dolan, R.J.; et al. Locus coeruleus integrity in old age is selectively related to memories linked with salient negative events. Proc. Natl. Acad. Sci. USA 2018, 115, 2228–2233. [Google Scholar] [CrossRef] [Green Version]
- Braak, H.; Del Tredici, K. Alzheimer’s disease: Pathogenesis and prevention. Alzheimers Dement. 2012, 8, 227–233. [Google Scholar] [CrossRef] [PubMed]
- Braak, H.; Thal, D.R.; Ghebremedhin, E.; Del Tredici, K. Stages of the pathologic process in Alzheimer disease: Age categories from 1 to 100 years. J. Neuropathol. Exp. Neurol. 2011, 70, 960–969. [Google Scholar] [CrossRef] [PubMed]
- Ehrenberg, A.J.; Nguy, A.K.; Theofilas, P.; Dunlop, S.; Suemoto, C.K.; Di Lorenzo Alho, A.T.; Leite, R.P.; Diehl Rodriguez, R.; Mejia, M.B.; Rüb, U.; et al. Quantifying the accretion of hyperphosphorylated tau in the locus coeruleus and dorsal raphe nucleus: The pathological building blocks of early Alzheimer’s disease. Neuropathol. Appl. Neurobiol. 2017, 43, 393–408. [Google Scholar] [CrossRef] [PubMed]
- McAlister, C.; Schmitter-Edgecombe, M. Everyday functioning and cognitive correlates in healthy older adults with subjective cognitive concerns. Clin. Neuropsychol. 2016, 30, 1087–1103. [Google Scholar] [CrossRef] [PubMed]
- Jonker, C.; Geerlings, M.I.; Schmand, B. Are memory complaints predictive for dementia? A review of clinical and population-based studies. Int. J. Geriatr. Psychiatry 2000, 15, 983–991. [Google Scholar] [CrossRef]
- Commissaris, C.; Ponds, R.; Jolles, J. Subjective forgetfulness in a normal Dutch population: Possibilities for health education and other interventions. Patient Educ. Couns. 1998, 34, 25–32. [Google Scholar] [CrossRef]
- Verhaeghen, P.; Geraerts, N.; Marcoen, A. Memory complaints, coping, and well-being in old age: A systemic approach. Gerontologist 2000, 40, 540–548. [Google Scholar] [CrossRef] [PubMed]
- Podewils, L.J.; McLay, R.N.; Rebok, G.W.; Lyketsos, C.G. Relationship of Self-Perceptions of Memory and Worry to Objective Measures of Memory and Cognition in the General Population. Psychosomatics 2003, 44, 461–470. [Google Scholar] [CrossRef] [PubMed]
- Wolfsgruber, S.; Wagner, M.; Schmidtke, K.; Froelich, L.; Kurz, A.; Schulz, S.R.; Hampel, H.; Heuser, I.; Peters, O.; Reischies, F.M.; et al. Memory concerns, memory performance and risk of dementia in patients with mild cognitive impairment. PLoS ONE 2014, 9, e100812. [Google Scholar] [CrossRef] [PubMed]
- Jessen, F.; Kleineidam, L.; Wolfsgruber, S.; Bickel, H.; Brettschneider, C.; Fuchs, A.; Kaduszkiewicz, H.; König, H.; Mallon, T.; Mamone, S.; et al. Prediction of dementia of Alzheimer type by different types of subjective cognitive decline. Alzheimers Dement. 2020, 16, 1745–1749. [Google Scholar] [CrossRef] [PubMed]
- Heffner, K.L.; Quinones, M.M.; Gallegos, A.M.; Crean, H.F.; Lin, F.; Suhr, J.A. Subjective memory in adults over 50 years of age: Associations with affective and physiological markers of emotion regulation. Aging Ment. Health. 2021, 1–9. [Google Scholar] [CrossRef]
- Lehmann, D.J.; Refsum, H.; Nurk, E.; Warden, D.R.; Tell, G.S.; Vollset, S.E.; Engedal, K.; Nygaard, H.A.; Smith, A.D. Apolipoprotein E epsilon4 and impaired episodic memory in community-dwelling elderly people: A marked sex difference. The Hordaland Health Study. J. Neurol. Neurosurg. Psychiatry 2006, 77, 902–908. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jolles, J.; Van Boxtel, M.P.; Ponds, R.W.; Metsemakers, J.F.; Houx, P.J. The Maastricht aging study (MAAS). The longitudinal perspective of cognitive aging. Tijdschrift Voor Gerontologie En Geriatrie 1998, 29, 120–129. [Google Scholar]
- Metsemakers, J.F.; Höppener, P.; Knottnerus, J.A.; Kocken, R.J.; Limonard, C.B. Computerized health information in The Netherlands: A registration network of family practices. Br. J. Gen. Pract. 1992, 42, 102–106. [Google Scholar] [PubMed]
- World Medical Association. World Medical Association Declaration of Helsinki: Ethical principles for medical research involving human subjects. JAMA 2013, 310, 2191–2194. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Folstein, M.F.; Folstein, S.E.; McHugh, P.R. “Mini-mental state”: A practical method for grading the cognitive state of patients for the clinician. J. Psychiatr. Res. 1975, 12, 189–198. [Google Scholar] [CrossRef]
- Van der Elst, W.; van Boxtel, M.P.; van Breukelen, G.J.; Jolles, J. Rey’s verbal learning test: Normative data for 1855 healthy participants aged 24–81 years and the influence of age, sex, education, and mode of presentation. J. Int. Neuropsychol. Soc. 2005, 11, 290–302. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van der Elst, W.; van Boxtel, M.P.J.; van Breukelen, G.J.P.; Jolles, J. The Letter Digit Substitution Test: Normative data for 1858 healthy participants aged 24–81 from the Maastricht Aging Study (MAAS): Influence of age, education, and sex. J. Clin. Exp. Neuropsychol. 2006, 28, 998–1009. [Google Scholar] [CrossRef] [PubMed]
- Van der Elst, W.; Van Boxtel, M.P.J.; Van Breukelen, G.J.P.; Jolles, J. The Stroop color-word test: Influence of age, sex, and education; and normative data for a large sample across the adult age range. Assessment 2006, 13, 62–79. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van der Elst, W.; Van Boxtel, M.P.J.; Van Breukelen, G.J.P.; Jolles, J. The concept shifting test: Adult normative data. Psychol. Assess. 2006, 18, 424–1939. [Google Scholar] [CrossRef] [PubMed]
- Hamilton, M. A Rating Scale for Depression. J. Neurol. Neurosurg. Psychiatry 1960, 23, 56. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ponds, R.W.H.M.; Jolles, J. The Abridged Dutch Metamemory in Adulthood (MIA) Questionnaire: Structure and effects of age, sex, and education. Psychol. Aging 1996, 11, 324–1939. [Google Scholar] [CrossRef] [PubMed]
- Sterpenich, V.; D’Argembeau, A.; Desseilles, M.; Balteau, E.; Albouy, G.; Vandewalle, G.; Degueldre, C.; Luxen, A.; Collette, F.; Maquet, P. The locus ceruleus is involved in the successful retrieval of emotional memories in humans. J. Neurosci. 2006, 26, 7416–7423. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lang, P.J.; Bradley, M.M.; Cuthbert, B.N. International Affective Picture System (IAPS): Affective Ratings of Pictures and Instruction Manual; Technical Report A-8; University of Florida: Gainesville, FL, USA, 2008. [Google Scholar]
- Kim, J.; Yassa, M.A. Assessing recollection and familiarity of similar lures in a behavioral pattern separation task. Hippocampus 2013, 23, 287–294. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dale, A.M. Optimal experimental design for event-related fMRI. Hum. Brain Mapp. 1999, 8, 109–114. [Google Scholar] [CrossRef]
- Willenbockel, V.; Sadr, J.; Fiset, D.; Horne, G.O.; Gosselin, F.; Tanaka, J.W. Controlling low-level image properties: The SHINE toolbox. Behav. Res. Methods 2010, 42, 671–684. [Google Scholar] [CrossRef] [PubMed]
- Marques, J.P.; Kober, T.; Krueger, G.; van der Zwaag, W.; Van de Moortele, P.F.; Gruetter, R. MP2RAGE, a self bias-field corrected sequence for improved segmentation and T1-mapping at high field. Neuroimage 2010, 49, 1271–1281. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Priovoulos, N.; Jacobs, H.I.L.; Ivanov, D.; Uludag, K.; Verhey, F.R.J.; Poser, B.A. High-resolution in vivo imaging of human locus coeruleus by Magnetization Transfer MRI at 3T and 7T. Neuroimage 2017, 168, 427–436. [Google Scholar] [CrossRef] [PubMed]
- Moeller, S.; Yacoub, E.; Olman, C.A.; Auerbach, E.; Strupp, J.; Harel, N.; Uğurbil, K. Multiband multislice GE-EPI at 7 tesla, with 16-fold acceleration using partial parallel imaging with application to high spatial and temporal whole-brain fMRI. Magn. Reson. Med. 2010, 63, 1144–1153. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Setsompop, K.; Gagoski, B.A.; Polimeni, J.R.; Witzel, T.; Wedeen, V.J.; Wald, L.L. Blipped-controlled aliasing in parallel imaging for simultaneous multislice echo planar imaging with reduced g-factor penalty. Magn. Reson. Med. 2012, 67, 1210–1224. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brooks, J.C.; Faull, O.K.; Pattinson, K.T.; Jenkinson, M. Physiological noise in brainstem FMRI. Front. Hum. Neurosci. 2013, 7, 623. [Google Scholar] [CrossRef] [Green Version]
- Fischl, B.; Sereno, M.I.; Dale, A.M. Cortical surface-based analysis: II: Inflation, flattening, and a surface-based coordinate system. Neuroimage 1999, 9, 195–207. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Iglesias, J.E.; Augustinack, J.C.; Nguyen, K.; Player, C.M.; Player, A.; Wright, M.; Roy, N.; Frosch, M.P.; McKee, A.C.; Wald, L.; et al. A computational atlas of the hippocampal formation using ex vivo, ultra-high resolution MRI: Application to adaptive segmentation of in vivo MRI. Neuroimage 2015, 115, 117–137. [Google Scholar] [CrossRef] [PubMed]
- Avants, B.B.; Tustison, N.; Song, G. Advanced normalization tools (ANTS). Insight J. 2009, 2, 1–35. [Google Scholar]
- Jenkinson, M.; Bannister, P.; Brady, M.; Smith, S. Improved Optimization for the Robust and Accurate Linear Registration and Motion Correction of Brain Images. NeuroImage 2002, 17, 825–841. [Google Scholar] [CrossRef] [PubMed]
- R Core Team. R: A Language and Environment for Statistical Computing [Computer Software Manual]; R Core Team: Vienna, Austria, 2016. [Google Scholar]
- Benjamini, Y.; Hochberg, Y. Controlling the false discovery rate: A practical and powerful approach to multiple testing. J. R. Stat. Soc. Ser. B (Methodol.) 1995, 57, 289–300. [Google Scholar] [CrossRef]
- United Nations Educational Service; Cultural Organization PDoSoE. International Standard Classification of Education (ISCED): ERIC Clearinghouse; 1976; Available online: https://www.ubos.org/wp-content/uploads/publications/03_2018International_Standard_Classification_of_Education__March_1976.pdf (accessed on 11 February 2022).
- Jacobs, H.I.; Priovoulos, N.; Poser, B.A.; Pagen, L.H.; Ivanov, D.; Verhey, F.R.; Uludağ, K. Dynamic behavior of the locus coeruleus during arousal-related memory processing in a multi-modal 7T fMRI paradigm. eLife 2020, 9, e52059. [Google Scholar] [CrossRef] [PubMed]
- Petrovich, G.D.; Canteras, N.S.; Swanson, L.W. Combinatorial amygdalar inputs to hippocampal domains and hypothalamic behavior systems. Brain Res. Rev. 2001, 38, 247–289. [Google Scholar] [CrossRef]
- Yang, Y.; Wang, J.Z. From Structure to Behavior in Basolateral Amygdala-Hippocampus Circuits. Front. Neural Circuits 2017, 11, 86. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hanert, A.; Pedersen, A.; Bartsch, T. Transient hippocampal CA1 lesions in humans impair pattern separation performance. Hippocampus 2019, 29, 736–747. [Google Scholar] [CrossRef] [PubMed]
- Leal, S.L.; Yassa, M.A. Effects of aging on mnemonic discrimination of emotional information. Behav. Neurosci. 2014, 128, 539–547. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kensinger, E.A. Remembering the details: Effects of emotion. Emot. Rev. 2009, 1, 99–113. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leal, S.L.; Ferguson, L.A.; Harrison, T.M.; Jagust, W.J. Development of a mnemonic discrimination task using naturalistic stimuli with applications to aging and preclinical Alzheimer’s disease. Learn. Mem. 2019, 26, 219–228. [Google Scholar] [CrossRef] [PubMed]
- Eysenck, M.W.; Derakshan, N. New perspectives in attentional control theory. Personal. Individ. Differ. 2011, 50, 955–960. [Google Scholar] [CrossRef]
- Priovoulos, N.; van Boxel, S.C.; Jacobs, H.I.; Poser, B.A.; Uludag, K.; Verhey, F.R.; Ivanov, D. Unraveling the contributions to the neuromelanin-MRI contrast. Brain Struct. Funct. 2020, 225, 2757–2774. [Google Scholar] [CrossRef] [PubMed]
- De Cicco, V.; Tramonti Fantozzi, M.P.; Cataldo, E.; Barresi, M.; Bruschini, L.; Faraguna, U.; Manzoni, D. Trigeminal, Visceral and Vestibular Inputs May Improve Cognitive Functions by Acting through the Locus Coeruleus and the Ascending Reticular Activating System: A New Hypothesis. Front. Neuroanat. 2017, 11, 130. [Google Scholar] [CrossRef] [PubMed]
- Tramonti Fantozzi, M.P.; De Cicco, V.; De Cicco, D.; d’Ascanio, P.; Cataldo, E.; Bruschini, L.; Faraguna, U.; Manzoni, D. Chewing and Cognitive Improvement: The Side Matters. Front. Syst. Neurosci. 2021, 15, 749444. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Mean (SD) | |
|---|---|
| N | 23 |
| Age (years) | 67.1 (4.9) |
| Sex n (% female) | 47.8% |
| Education level | 4.3 (1.6) |
| MMSE (score) | 29.3 (0.8) |
| VLT-total (nr words) | 46.7 (8.5) |
| VLT-delayed (nr words) | 9.2 (2.5) |
| HDRS (score) | 3.8 (3.6) |
| MIA anxiety (score) | 2.8 (0.6) |
| High Arousal Hits (score) | 0.6 (0.08) |
| Low Arousal Hits (score) | 0.7 (0.08) |
| High Arousal FP (score) | 0.5 (0.2) |
| Low Arousal FP (score) | 0.4 (0.2) |
| High Arousal FN (score) | 0.2 (0.1) |
| Low Arousal FN (score) | 0.2 (0.1) |
| Est. | t | p | P (FDR) | |
|---|---|---|---|---|
| Arousal Contrast: Hit HA-LA | ||||
| Amygdala | 0.03 | 0.06 | 0.95 | 0.95 |
| CA1 | 0.84 | 2.08 | 0.047 | 0.12 |
| CA3 | 0.31 | 1.06 | 0.30 | 0.50 |
| DG | 0.63 | 2.82 | 0.01 | 0.05 |
| LC | 0.04 | 0.33 | 0.74 | 0.93 |
| Arousal Contrast: Hit LA-HA | ||||
| Amygdala | −0.22 | −0.57 | 0.57 | 0.71 |
| CA1 | −0.85 | −1.43 | 0.18 | 0.71 |
| CA3 | −0.07 | −0.20 | 0.84 | 0.84 |
| DG | −0.24 | −0.66 | 0.51 | 0.71 |
| LC | −0.09 | −0.59 | 0.56 | 0.71 |
| Arousal Contrast: FP HA-LA | ||||
| Amygdala | 0.54 | 1.39 | 0.18 | 0.40 |
| CA1 | −0.12 | −0.22 | 0.83 | 0.83 |
| CA3 | −0.10 | −0.33 | 0.74 | 0.83 |
| DG | −0.41 | −1.41 | 0.17 | 0.40 |
| LC | −0.29 | −1.21 | 0.24 | 0.40 |
| Arousal Contrast: FP LA-HA | ||||
| Amygdala | −0.34 | −0.97 | 0.33 | 0.48 |
| CA1 | −0.27 | −0.53 | 0.59 | 0.59 |
| CA3 | 0.23 | 0.86 | 0.38 | 0.48 |
| DG | 0.28 | 0.91 | 0.37 | 0.48 |
| LC | 0.28 | 1.04 | 0.31 | 0.48 |
| Memory Contrast: HA Hit-FP | ||||
| Amygdala | −0.69 | −2.22 | 0.037 | 0.19 |
| CA1 | −0.06 | −0.17 | 0.87 | 0.87 |
| CA3 | −0.35 | −1.49 | 0.15 | 0.32 |
| DG | −0.32 | −1.35 | 0.19 | 0.32 |
| LC | −0.10 | −0.55 | 0.59 | 0.74 |
| Memory Contrast: LA Hit-FP | ||||
| Amygdala | −0.16 | −0.58 | 0.58 | 0.73 |
| CA1 | −0.36 | −1.03 | 0.30 | 0.73 |
| CA3 | −0.03 | −0.15 | 0.88 | 0.88 |
| DG | 0.13 | 0.56 | 0.58 | 0.73 |
| LC | 0.13 | 0.74 | 0.47 | 0.73 |
| Memory Contrast: HA FP-Hit | ||||
| Amygdala | −0.57 | −0.98 | 0.36 | 0.50 |
| CA1 | −0.60 | −0.88 | 0.40 | 0.50 |
| CA3 | −0.71 | −1.40 | 0.18 | 0.45 |
| DG | −0.82 | −1.65 | 0.12 | 0.45 |
| LC | −0.10 | −0.32 | 0.75 | 0.75 |
| Memory Contrast: LA FP-Hit | ||||
| Amygdala | 0.17 | 0.34 | 0.74 | 0.74 |
| CA1 | −0.21 | −0.35 | 0.73 | 0.74 |
| CA3 | 0.35 | 1.24 | 0.23 | 0.47 |
| DG | 0.52 | 1.57 | 0.13 | 0.47 |
| LC | 0.33 | 1.25 | 0.28 | 0.47 |
| Est. | t | p | P (FDR) | |
|---|---|---|---|---|
| Arousal Contrast: Hit HA-LA | ||||
| Amygdala | 0.32 | 0.46 | 0.65 | 0.81 |
| CA1 | 1.31 | 1.40 | 0.19 | 0.35 |
| CA3 | 0.83 | 1.28 | 0.21 | 0.35 |
| DG | 0.03 | 0.07 | 0.94 | 0.94 |
| LC | 0.52 | 2.1 | 0.045 | 0.23 |
| Arousal Contrast: Hit LA-HA | ||||
| Amygdala | 0.02 | 0.03 | 0.98 | 0.98 |
| CA1 | 1.01 | 0.87 | 0.40 | 0.92 |
| CA3 | −0.57 | −0.89 | 0.38 | 0.92 |
| DG | −0.08 | −0.13 | 0.90 | 0.98 |
| LC | −0.21 | −0.61 | 0.55 | 0.92 |
| Arousal Contrast: FP HA-LA | ||||
| Amygdala | 0.94 | 1.20 | 0.24 | 0.99 |
| CA1 | 0.02 | 0.02 | 0.99 | 0.99 |
| CA3 | −0.11 | −0.21 | 0.84 | 0.99 |
| DG | 0.39 | 0.62 | 0.55 | 0.99 |
| LC | −0.11 | −0.23 | 0.82 | 0.99 |
| Arousal Contrast: FP LA-HA | ||||
| Amygdala | −0.21 | −0.26 | 0.79 | 0.99 |
| CA1 | 0.50 | 0.49 | 0.63 | 0.99 |
| CA3 | −0.05 | −0.11 | 0.91 | 0.99 |
| DG | −0.28 | −0.45 | 0.66 | 0.99 |
| LC | 0.01 | 0.01 | 0.99 | 0.99 |
| Memory Contrast: HA Hit-FP | ||||
| Amygdala | 1.30 | 1.76 | 0.10 | 0.50 |
| CA1 | 1.06 | 1.23 | 0.23 | 0.58 |
| CA3 | 0.20 | 0.48 | 0.64 | 0.69 |
| DG | 0.19 | 0.40 | 0.69 | 0.69 |
| LC | 0.30 | 0.78 | 0.44 | 0.69 |
| Memory Contrast: LA Hit-FP | ||||
| Amygdala | 0.38 | 0.60 | 0.55 | 0.69 |
| CA1 | 0.30 | 0.65 | 0.53 | 0.69 |
| CA3 | 0.15 | 0.50 | 0.62 | 0.69 |
| DG | −0.18 | −0.41 | 0.69 | 0.69 |
| LC | −0.61 | −3.13 | 0.006 | 0.03 |
| Memory Contrast: HA FP-Hit | ||||
| Amygdala | 1.21 | 0.78 | 0.44 | 0.69 |
| CA1 | 0.83 | 0.62 | 0.55 | 0.69 |
| CA3 | −0.04 | −0.04 | 0.97 | 0.97 |
| DG | 0.77 | 0.85 | 0.43 | 0.69 |
| LC | 0.68 | 1.18 | 0.26 | 0.69 |
| Memory Contrast: LA FP-Hit | ||||
| Amygdala | −0.80 | −0.67 | 0.51 | 0.82 |
| CA1 | 0.22 | 0.24 | 0.81 | 0.82 |
| CA3 | −0.11 | −0.23 | 0.82 | 0.82 |
| DG | −0.77 | −1.07 | 0.30 | 0.75 |
| LC | −1.25 | −2.92 | 0.009 | 0.045 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pagen, L.H.G.; Poser, B.A.; van Boxtel, M.P.J.; Priovoulos, N.; van Hooren, R.W.E.; Verhey, F.R.J.; Jacobs, H.I.L. Worry Modifies the Relationship between Locus Coeruleus Activity and Emotional Mnemonic Discrimination. Brain Sci. 2022, 12, 381. https://doi.org/10.3390/brainsci12030381
Pagen LHG, Poser BA, van Boxtel MPJ, Priovoulos N, van Hooren RWE, Verhey FRJ, Jacobs HIL. Worry Modifies the Relationship between Locus Coeruleus Activity and Emotional Mnemonic Discrimination. Brain Sciences. 2022; 12(3):381. https://doi.org/10.3390/brainsci12030381
Chicago/Turabian StylePagen, Linda H. G., Benedikt A. Poser, Martin P. J. van Boxtel, Nikos Priovoulos, Roy W. E. van Hooren, Frans R. J. Verhey, and Heidi I. L. Jacobs. 2022. "Worry Modifies the Relationship between Locus Coeruleus Activity and Emotional Mnemonic Discrimination" Brain Sciences 12, no. 3: 381. https://doi.org/10.3390/brainsci12030381