
Hesperidin Protects Human HaCaT Keratinocytes from Particulate Matter 2.5-Induced Apoptosis via the Inhibition of Oxidative Stress and Autophagy


 and
and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Reagents
2.2. Cell Culture
2.3. Intracellular ROS Detection
2.4. Cell Viability Assessment
2.5. Lipid Peroxidation Assay
2.6. Protein Carbonylation Assay
2.7. Single-Cell Gel Electrophoresis Assay (Comet Assay)
2.8. Detection of 8-Oxoguanine (8-oxoG)
2.9. Quantification of Cellular Ca2+ Level
2.10. Mitochondrial Membrane Potential (Δψm) Analysis
2.11. Acridine Orange Morphology Assessment
2.12. Western Blotting
2.13. Hoechst 33342 Staining
2.14. Statistical Analysis
3. Results
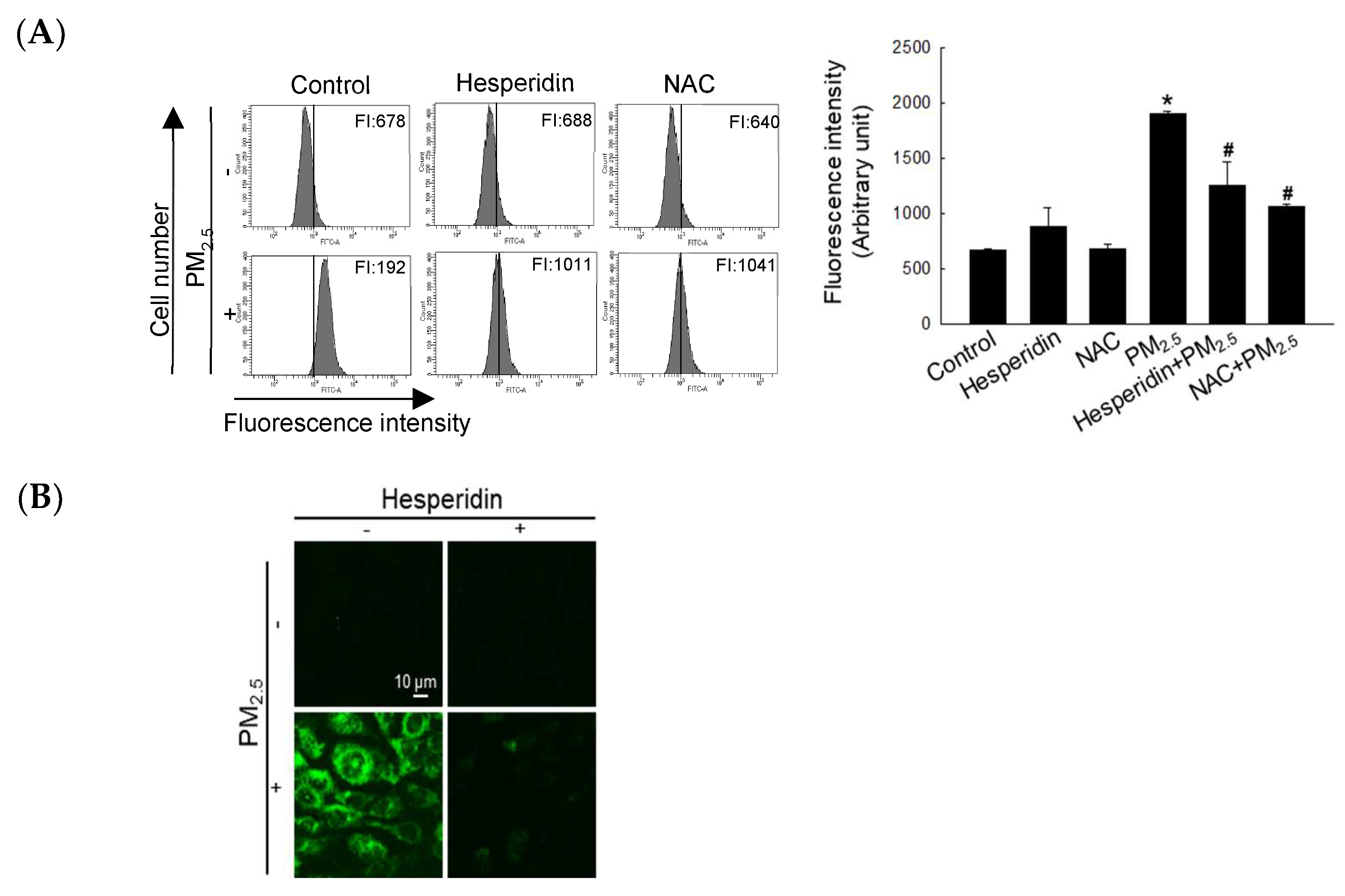
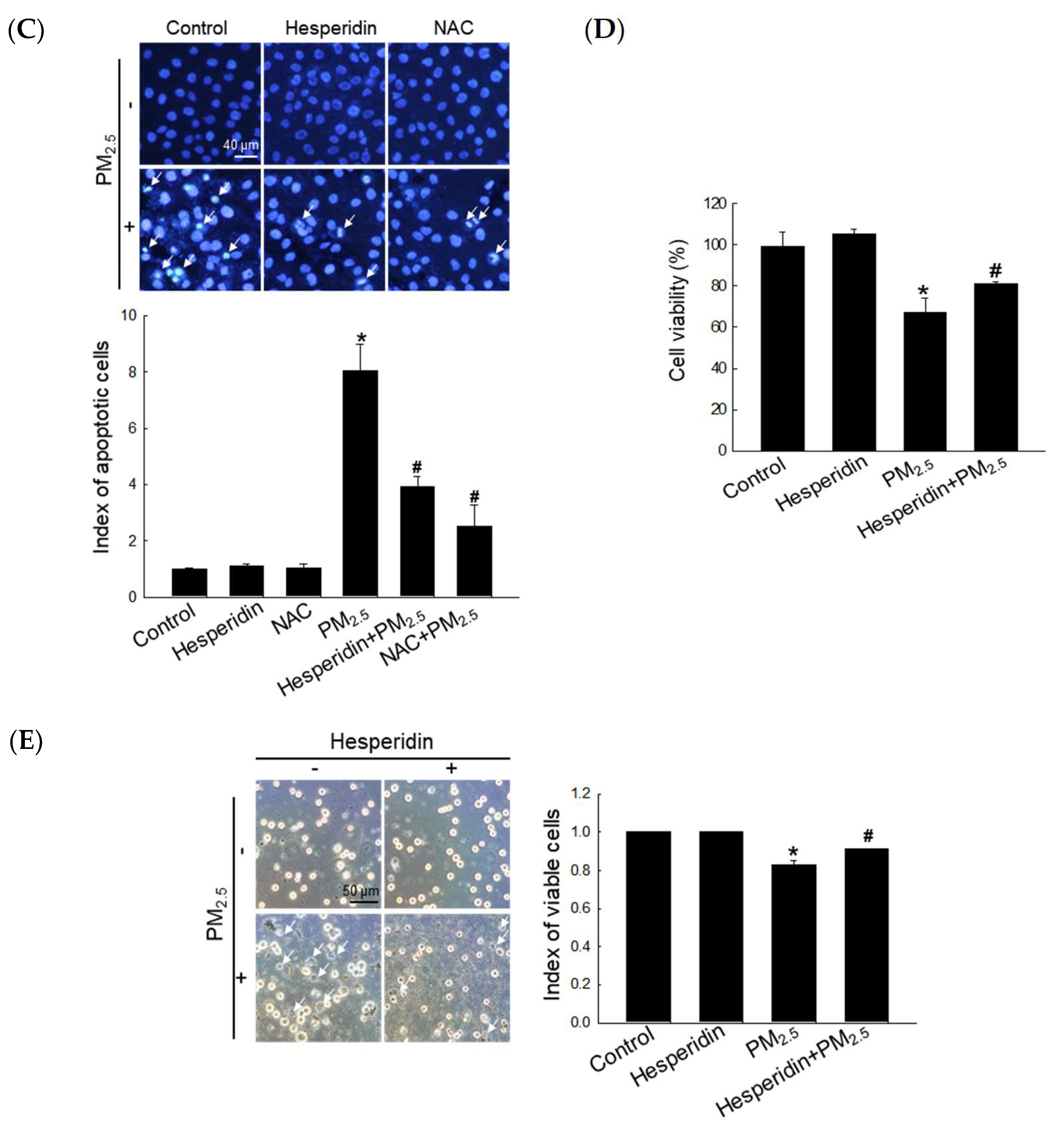
3.1. Hesperidin Restored PM2.5-Mediated Reduced Cell Viability by Mitigating Intracellular ROS Generation
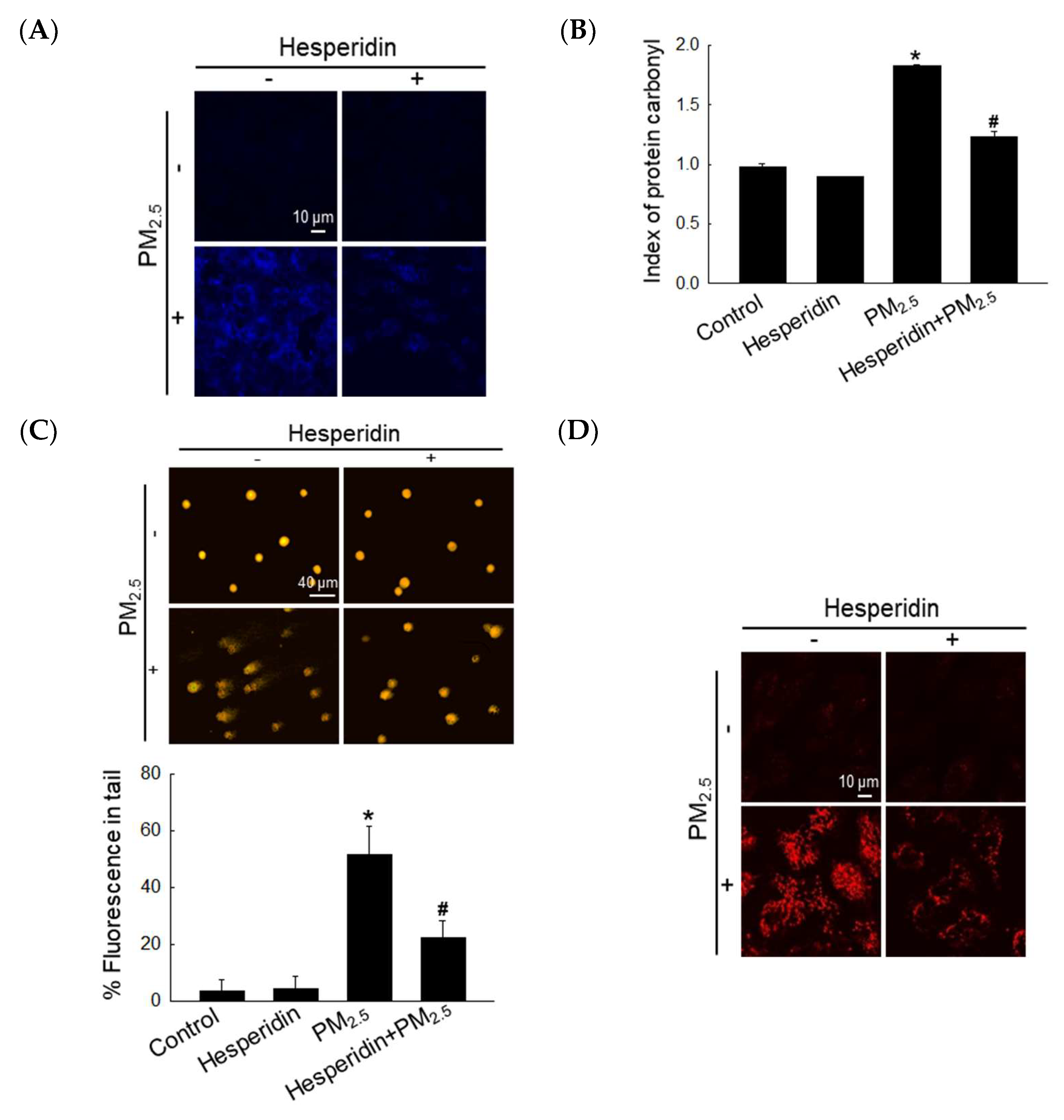
3.2. Hesperidin Inhibited the Damage of Cellular Components by PM2.5-Induced Oxidative Stress
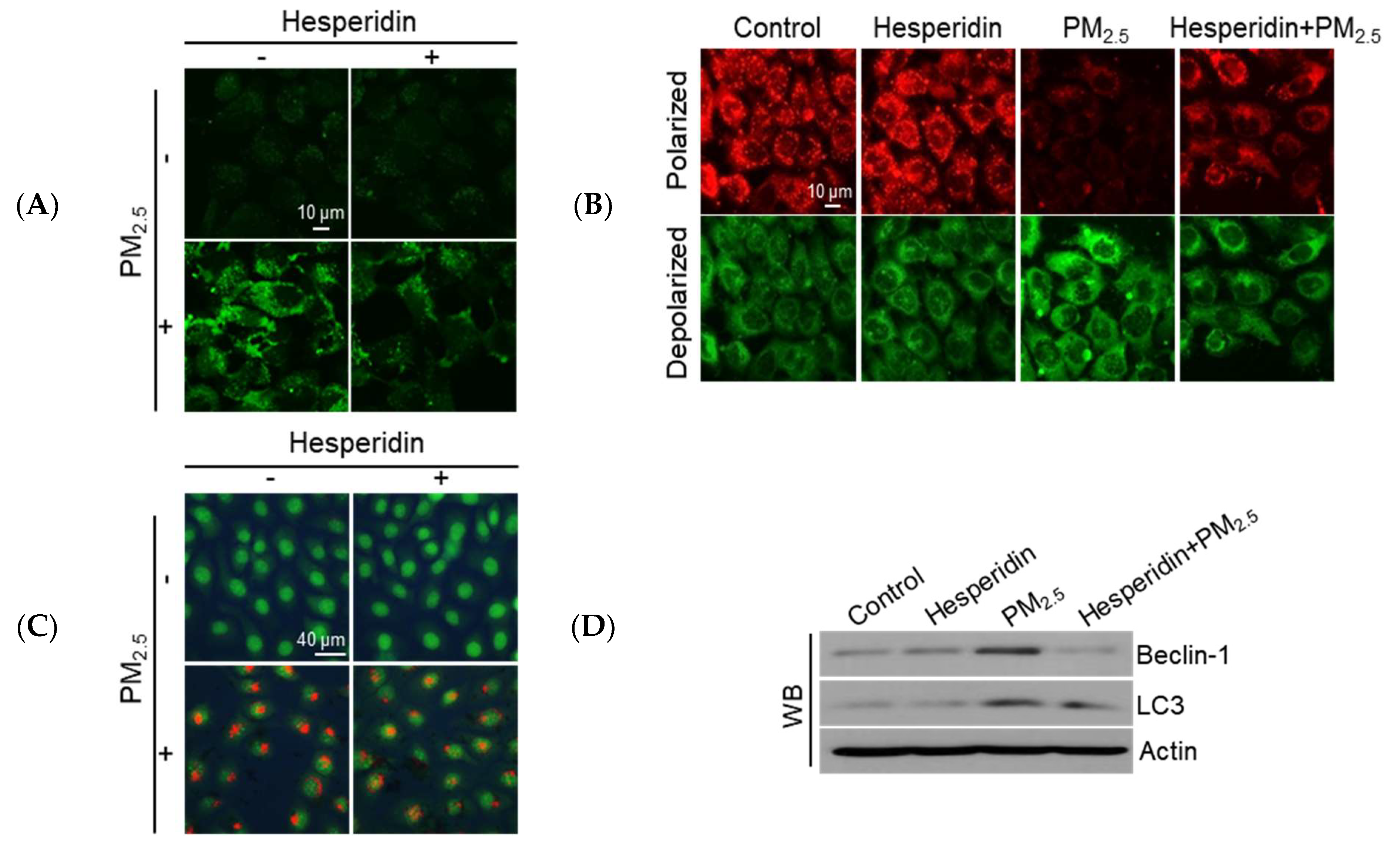
3.3. Hesperidin Mitigated PM2.5-Induced Mitochondrial Dysfunction and Autophagy Activation
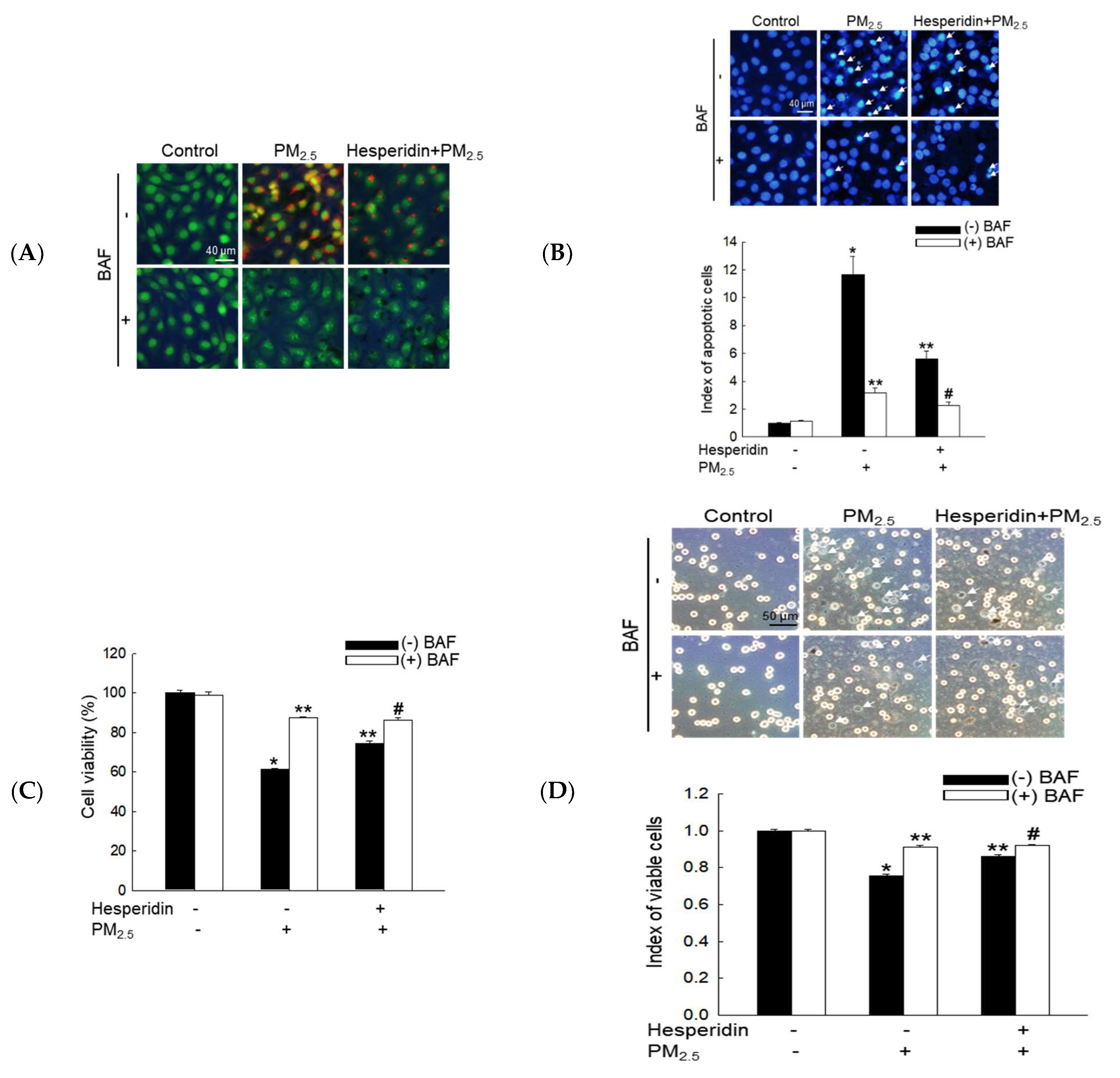
3.4. Hesperidin Ameliorated Cellular Apoptosis via the Inhibition of PM2.5-Induced Autophagy
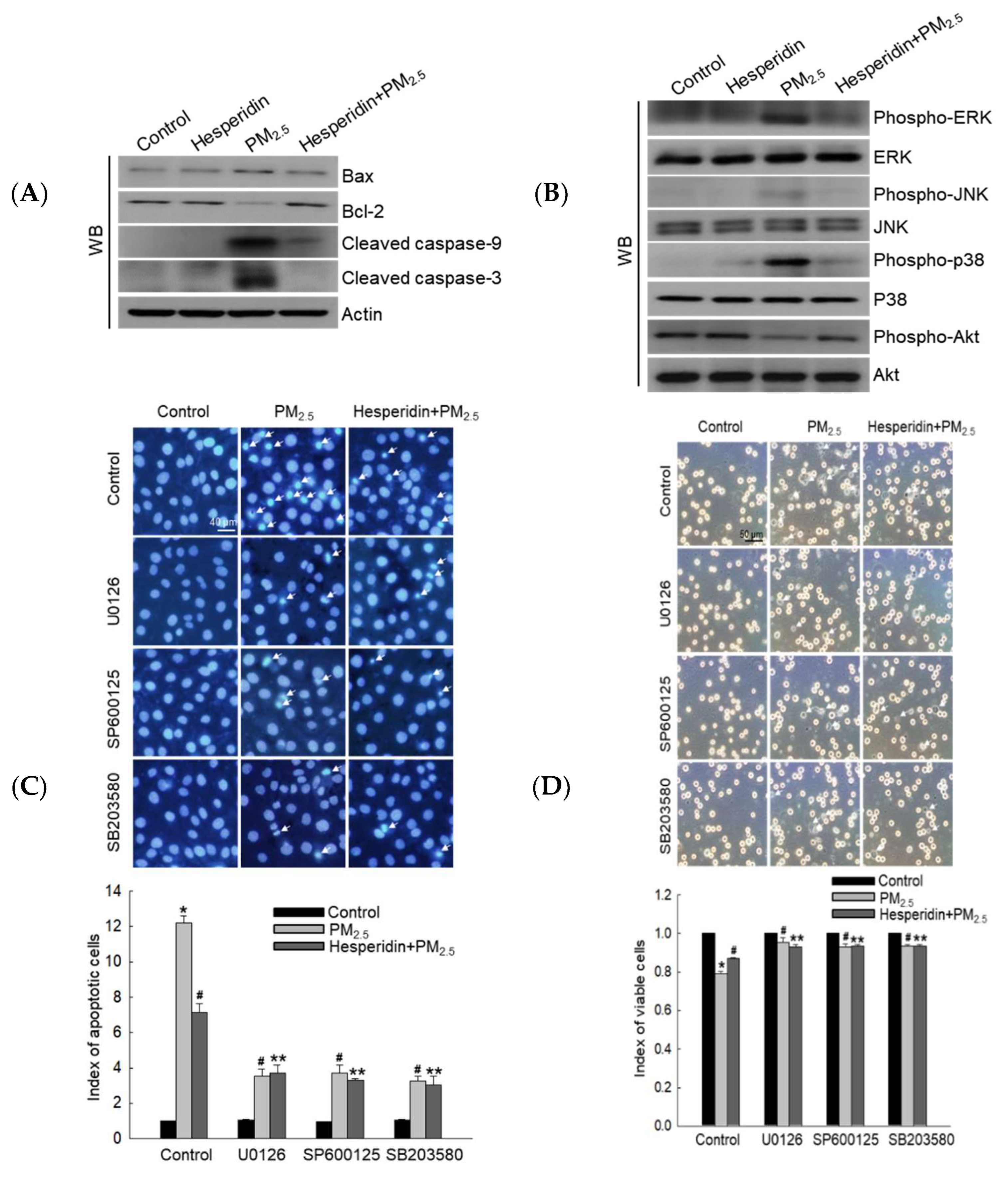
3.5. Hesperidin Inhibited PM2.5-Induced Cellular Apoptosis via MAPK Inactivation
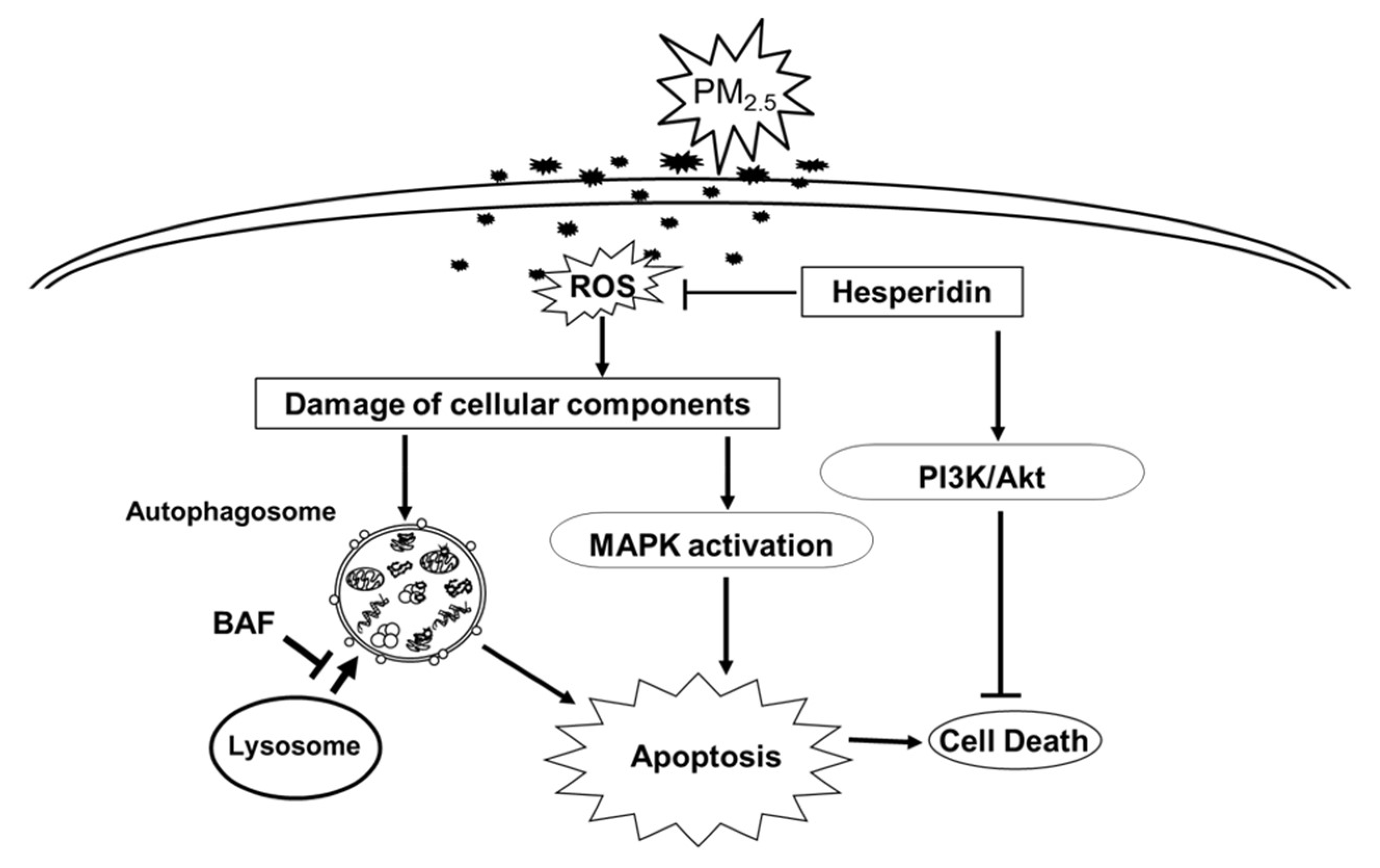
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Lahmer, N.; Belboukhari, N.; Cheriti, A.; Sekkoum, K. Hesperidin and hesperitin preparation and purification from citrus sinensis peels. Der Pharma Chem. 2015, 7, 1–4. [Google Scholar]
- Ahmadi, A.; Shadboorestan, A. Oxidative stress and cancer; the role of hesperidin, a citrus natural bioflavonoid, as a cancer chemoprotective agent. Nutr. Cancer 2015, 68, 29–39. [Google Scholar] [CrossRef] [PubMed]
- Iranshahi, M.; Rezaee, R.; Parhiz, H.; Roohbakhsh, A.; Soltani, F. Protective effects of flavonoids against microbes and toxins: The cases of hesperidin and hesperetin. Life Sci. 2015, 137, 125–132. [Google Scholar] [CrossRef] [PubMed]
- El Barky, A.R.; Mohamed, T.M. Isolation, characterization and the biological activity of some natural components of marine sea cucumber and orange peel. Biomed. J. Sci. Tech. Res. 2020, 27, 20555–20565. [Google Scholar] [CrossRef]
- Singh, V.; Singh, S.; Biswal, A. Exceedances and trends of particulate matter (PM2.5) in five Indian megacities. Sci. Total Environ. 2021, 750, 141461. [Google Scholar] [CrossRef]
- Fiordelisi, A.; Piscitelli, P.; Trimarco, B.; Coscioni, E.; Iaccarino, G.; Sorriento, D. The mechanisms of air pollution and particulate matter in cardiovascular diseases. Heart Fail. Rev. 2017, 22, 337–347. [Google Scholar] [CrossRef]
- Janta, R.; Sekiguchi, K.; Yamaguchi, R.; Sopajaree, K.; Pongpiachan, S.; Chetiyanukornkul, T. Ambient PM2.5, polycyclic aromatic hydrocarbons and biomass burning tracer in Mae Sot district, western Thailand. Atmos. Pollut. Res. 2020, 11, 27–39. [Google Scholar] [CrossRef]
- Lei, W.; Zhang, L.; Xu, J.; Liu, Z.; Xin, J.; Li, X.; Zhao, W. Spatiotemporal variations and source apportionment of metals in atmospheric particulate matter in Beijing and its surrounding areas. Atmos. Pollut. Res. 2021, 12, 101–213. [Google Scholar] [CrossRef]
- Rönkkö, T.J.; Hirvonen, M.R.; Happo, M.S.; Ihantola, T.; Hakkarainen, H.; Martikainen, M.V.; Gu, C.; Wang, Q.G.; Jokiniemi, J.; Komppula, M.; et al. Inflammatory responses of urban air PM modulated by chemical composition and different air quality situations in Nanjing, China. Environ. Res. 2021, 192, 110382. [Google Scholar] [CrossRef]
- Kumar, P.; Kalaiarasan, G.; Porter, A.E.; Pinna, A.; Kłosowski, M.M.; Demokritou, P.; Chung, K.F.; Pain, C.; Arvind, D.K.; Arcucci, R.; et al. An overview of methods of fine and ultrafine particle collection for physicochemical characterisation and toxicity assessments. Sci. Total Environ. 2021, 756, 143553. [Google Scholar] [CrossRef]
- Piao, M.J.; Ahn, M.J.; Kang, K.A.; Ryu, Y.S.; Hyun, Y.J.; Shilnikova, K.; Zhen, A.X.; Jeong, J.W.; Choi, Y.H.; Kang, H.K.; et al. Particulate matter 2.5 damages skin cells by inducing oxidative stress, subcellular organelle dysfunction, and apoptosis. Arch. Toxicol. 2018, 92, 2077–2091. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ryu, Y.S.; Kang, K.A.; Piao, M.J.; Ahn, M.J.; Yi, J.M.; Bossis, G.; Hyun, Y.M.; Park, C.O.; Hyun, J.W. Particulate matter-induced senescence of skin keratinocytes involves oxidative stress-dependent epigenetic modifications. Exp. Mol. Med. 2019, 51, 1–14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ryu, Y.S.; Kang, K.A.; Piao, M.J.; Ahn, M.J.; Yi, J.M.; Hyun, Y.M.; Kim, S.H.; Ko, M.K.; Park, C.O.; Hyun, J.W. Particulate matter induces inflammatory cytokine production via activation of NFκB by TLR5-NOX4-ROS signaling in human skin keratinocyte and mouse skin. Redox Biol. 2019, 21, 101080. [Google Scholar] [CrossRef] [PubMed]
- Dąbrowska, A.K.; Spano, F.; Derler, S.; Adlhart, C.; Spencer, N.D.; Rossi, R.M. The relationship between skin function, barrier properties, and body-dependent factors. Skin Res. Technol. 2018, 24, 165–174. [Google Scholar] [CrossRef] [Green Version]
- Gomes, A.P.; Blenis, J. A nexus for cellular homeostasis: The interplay between metabolic and signal transduction pathways. Curr. Opin. Biotechnol. 2015, 34, 110–117. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.; Chen, Y.Y.; Cao, J.Y.; Tao, F.B.; Zhu, X.X.; Yao, C.J.; Chen, D.J.; Che, Z.; Zhao, Q.H.; Wen, L.P. Oxidative stress, apoptosis, and cell cycle arrest are induced in primary fetal alveolar type II epithelial cells exposed to fine particulate matter from cooking oil fumes. Environ. Sci. Pollut. Res. Int. 2015, 22, 9728–9741. [Google Scholar] [CrossRef]
- Ryu, Y.S.; Fernando, P.D.S.M.; Kang, K.A.; Piao, M.J.; Zhen, A.X.; Kang, H.K.; Koh, Y.S.; Hyun, J.W. Marine compound 3-bromo-4, 5-dihydroxybenzaldehyde protects skin cells against oxidative damage via the Nrf2/HO-1 pathway. Mar. Drugs 2019, 17, 234. [Google Scholar] [CrossRef] [Green Version]
- Kang, K.A.; Wang, Z.H.; Zhang, R.; Piao, M.J.; Kim, K.C.; Kang, S.S.; Kim, Y.W.; Lee, J.; Park, D.; Hyun, J.W. Myricetin protects cells against oxidative stress-induced apoptosis via regulation of PI3K/Akt and MAPK signaling pathways. Int. J. Mol. Sci. 2010, 11, 4348–4360. [Google Scholar] [CrossRef] [Green Version]
- Zhen, A.X.; Hyun, Y.J.; Piao, M.J.; Fernando, P.D.S.M.; Kang, K.A.; Ahn, M.J.; Yi, J.M.; Kang, H.K.; Koh, Y.S.; Lee, N.H.; et al. Eckol inhibits particulate matter 2.5-induced skin keratinocyte damage via MAPK signaling pathway. Mar. Drugs 2019, 17, 444. [Google Scholar] [CrossRef] [Green Version]
- Hyun, Y.J.; Piao, M.J.; Kang, K.A.; Zhen, A.X.; Fernando, P.D.S.M.; Kang, H.K.; Ahn, Y.S.; Hyun, J.W. Effect of fermented fish oil on fine particulate matter-induced skin aging. Mar. Drugs 2019, 17, 61. [Google Scholar] [CrossRef] [Green Version]
- Zhen, A.X.; Piao, M.J.; Hyun, Y.J.; Kang, K.A.; Fernando, P.D.S.M.; Cho, S.J.; Ahn, M.J.; Hyun, J.W. Diphlorethohydroxycarmalol attenuates fine particulate matter-induced subcellular skin dysfunction. Mar. Drugs 2019, 17, 95. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hewage, S.R.K.M.; Piao, M.J.; Kang, K.A.; Ryu, Y.S.; Han, X.; Oh, M.C.; Jung, U.; Kim, I.G.; Hyun, J.W. Hesperidin attenuates ultraviolet B-induced apoptosis by mitigating oxidative stress in human keratinocytes. Biomol. Ther. 2016, 24, 312–319. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Görlach, A.; Bertram, K.; Hudecova, S.; Krizanova, O. Calcium and ROS: A mutual interplay. Redox Biol. 2015, 6, 260–271. [Google Scholar] [CrossRef] [Green Version]
- Yu, K.N.; Chang, S.H.; Park, S.J.; Lim, J.; Lee, J.; Yoon, T.J.; Kim, J.S.; Cho, M.H. Titanium dioxide nanoparticles induce endoplasmic reticulum stress-mediated autophagic cell death via mitochondria-associated endoplasmic reticulum membrane disruption in normal lung cells. PLoS ONE 2015, 10, e0131208. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lin, D.S.; Huang, Y.W.; Ho, C.S.; Hung, P.L.; Hsu, M.H.; Wang, T.J.; Wu, T.Y.; Lee, T.H.; Huang, Z.D.; Chang, P.C.; et al. Oxidative insults and mitochondrial DNA mutation promote enhanced autophagy and mitophagy compromising cell viability in pluripotent cell model of mitochondrial disease. Cells 2019, 8, 65. [Google Scholar] [CrossRef] [Green Version]
- Park, S.H.; Park, H.S.; Lee, J.H.; Chi, G.Y.; Kim, G.Y.; Moon, S.K.; Chang, Y.C.; Hyun, J.W.; Kim, W.J.; Choi, Y.H. Induction of endoplasmic reticulum stress-mediated apoptosis and non-canonical autophagy by luteolin in NCI-H460 lung carcinoma cells. Food Chem. Toxicol. 2013, 56, 100–109. [Google Scholar] [CrossRef]
- Yi, C.; Si, L.; Xu, J.; Yang, J.; Wang, Q.; Wang, X. Effect and mechanism of asiatic acid on autophagy in myocardial ischemia-reperfusion injury in vivo and in vitro. Exp. The. Med. 2020, 20, 54. [Google Scholar] [CrossRef]
- Wang, K. Autophagy and apoptosis in liver injury. Cell Cycle 2015, 14, 1631–1642. [Google Scholar] [CrossRef] [Green Version]
- Carresi, C.; Mollace, R.; Macrì, R.; Scicchitano, M.; Bosco, F.; Scarano, F.; Coppoletta, A.R.; Guarnieri, L.; Ruga, S.; Zito, M.C.; et al. Oxidative stress triggers defective autophagy in endothelial cells: Role in atherothrombosis development. Antioxidants 2021, 10, 387. [Google Scholar] [CrossRef]
- Kang, K.A.; Piao, M.J.; Ryu, Y.S.; Hyun, Y.J.; Park, J.E.; Shilnikova, K.; Zhen, A.X.; Kang, H.K.; Koh, Y.S.; Jeong, Y.J.; et al. Luteolin induces apoptotic cell death via antioxidant activity in human colon cancer cells. Int. J. Oncol. 2017, 51, 1169–1178. [Google Scholar] [CrossRef] [Green Version]
- Hu, R.; Xie, X.Y.; Xu, S.K.; Wang, Y.N.; Jiang, M.; Wen, L.R.; Lai, W.; Guan, L. PM2.5 exposure elicits oxidative stress responses and mitochondrial apoptosis pathway activation in HaCaT keratinocytes. Chin. Med. J. 2017, 130, 2205–2214. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Zhong, Y.; Liao, J.; Wang, G. PM2.5-related cell death patterns. Int. J. Med. Sci. 2021, 18, 1024. [Google Scholar] [CrossRef] [PubMed]

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fernando, P.D.S.M.; Piao, M.J.; Kang, K.A.; Zhen, A.X.; Herath, H.M.U.L.; Kang, H.K.; Choi, Y.H.; Hyun, J.W. Hesperidin Protects Human HaCaT Keratinocytes from Particulate Matter 2.5-Induced Apoptosis via the Inhibition of Oxidative Stress and Autophagy. Antioxidants 2022, 11, 1363. https://doi.org/10.3390/antiox11071363
Fernando PDSM, Piao MJ, Kang KA, Zhen AX, Herath HMUL, Kang HK, Choi YH, Hyun JW. Hesperidin Protects Human HaCaT Keratinocytes from Particulate Matter 2.5-Induced Apoptosis via the Inhibition of Oxidative Stress and Autophagy. Antioxidants. 2022; 11(7):1363. https://doi.org/10.3390/antiox11071363
Chicago/Turabian StyleFernando, Pincha Devage Sameera Madushan, Mei Jing Piao, Kyoung Ah Kang, Ao Xuan Zhen, Herath Mudiyanselage Udari Lakmini Herath, Hee Kyoung Kang, Yung Hyun Choi, and Jin Won Hyun. 2022. "Hesperidin Protects Human HaCaT Keratinocytes from Particulate Matter 2.5-Induced Apoptosis via the Inhibition of Oxidative Stress and Autophagy" Antioxidants 11, no. 7: 1363. https://doi.org/10.3390/antiox11071363
APA StyleFernando, P. D. S. M., Piao, M. J., Kang, K. A., Zhen, A. X., Herath, H. M. U. L., Kang, H. K., Choi, Y. H., & Hyun, J. W. (2022). Hesperidin Protects Human HaCaT Keratinocytes from Particulate Matter 2.5-Induced Apoptosis via the Inhibition of Oxidative Stress and Autophagy. Antioxidants, 11(7), 1363. https://doi.org/10.3390/antiox11071363