Novel Polymeric Nanocarriers Reduced Zinc and Doxycycline Toxicity in the Nematode Caenorhabditis elegans


 ,
,  ,
, 

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Nanoparticles Production
2.2. Nanoparticle Characterization
2.3. Loading Efficacy of Zinc and Doxycycline
2.4. Test Organism
2.5. Lethal Toxicity Test
2.6. Metabolism: Pharyngeal Pumping Assay
2.7. Body Length: Growth Test
2.8. Reproduction and Fertility Asays
2.9. ROS Production
2.10. Statistical Analysis
3. Results
3.1. Nanoparticle Characterization
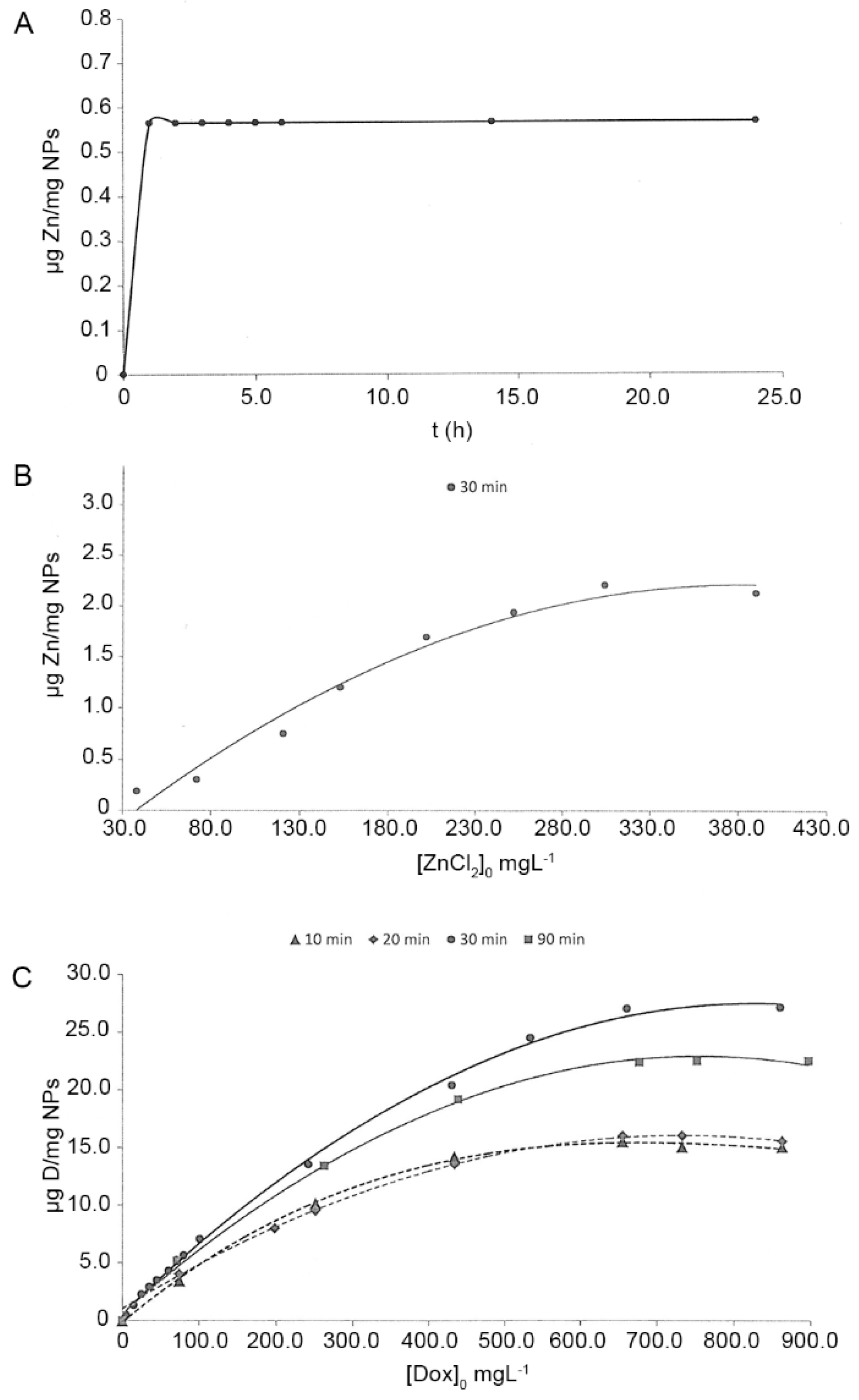
3.2. Loading Efficacy of Zinc and Doxycycline
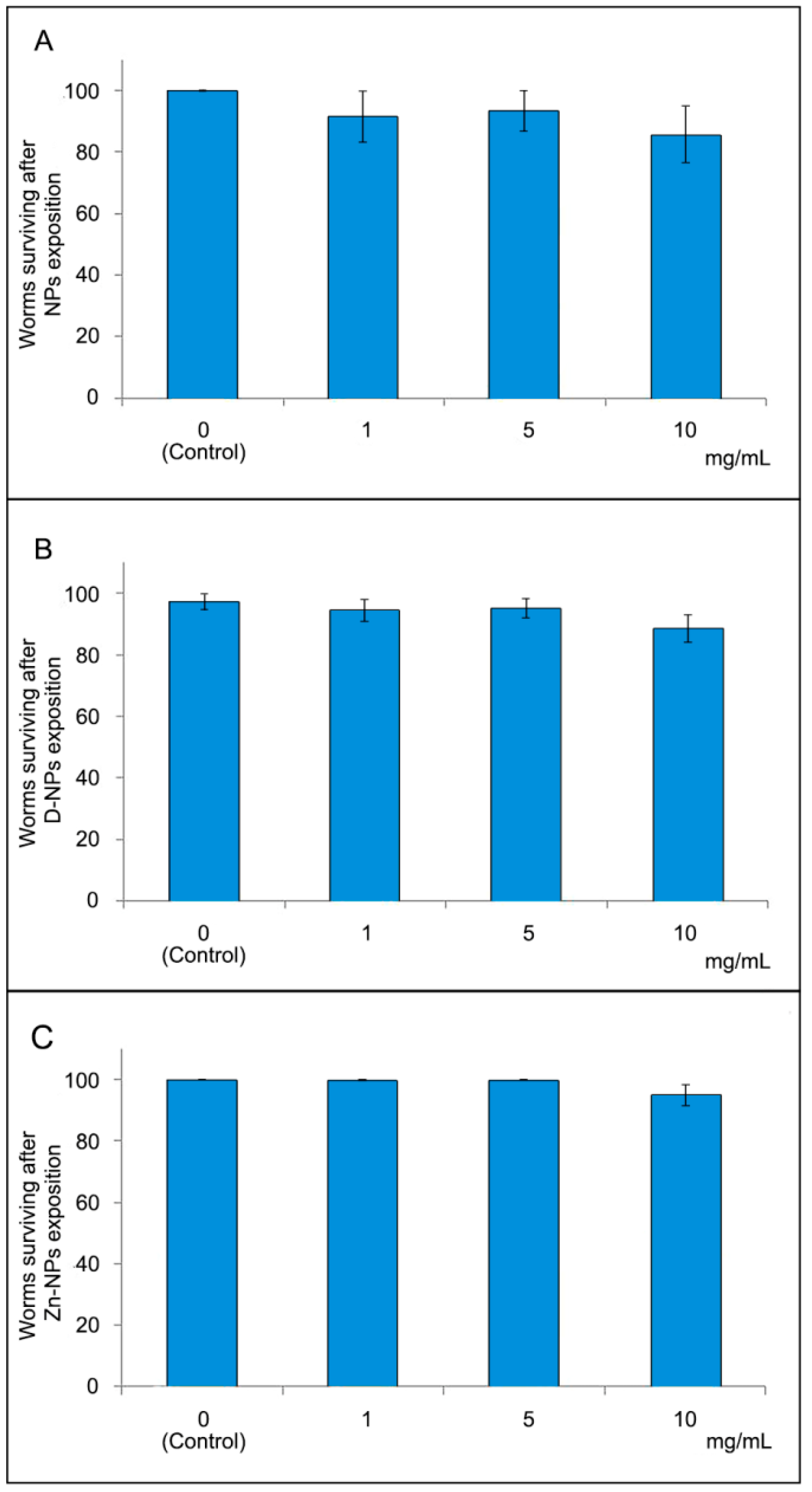
3.3. Lethal Toxicity Assay
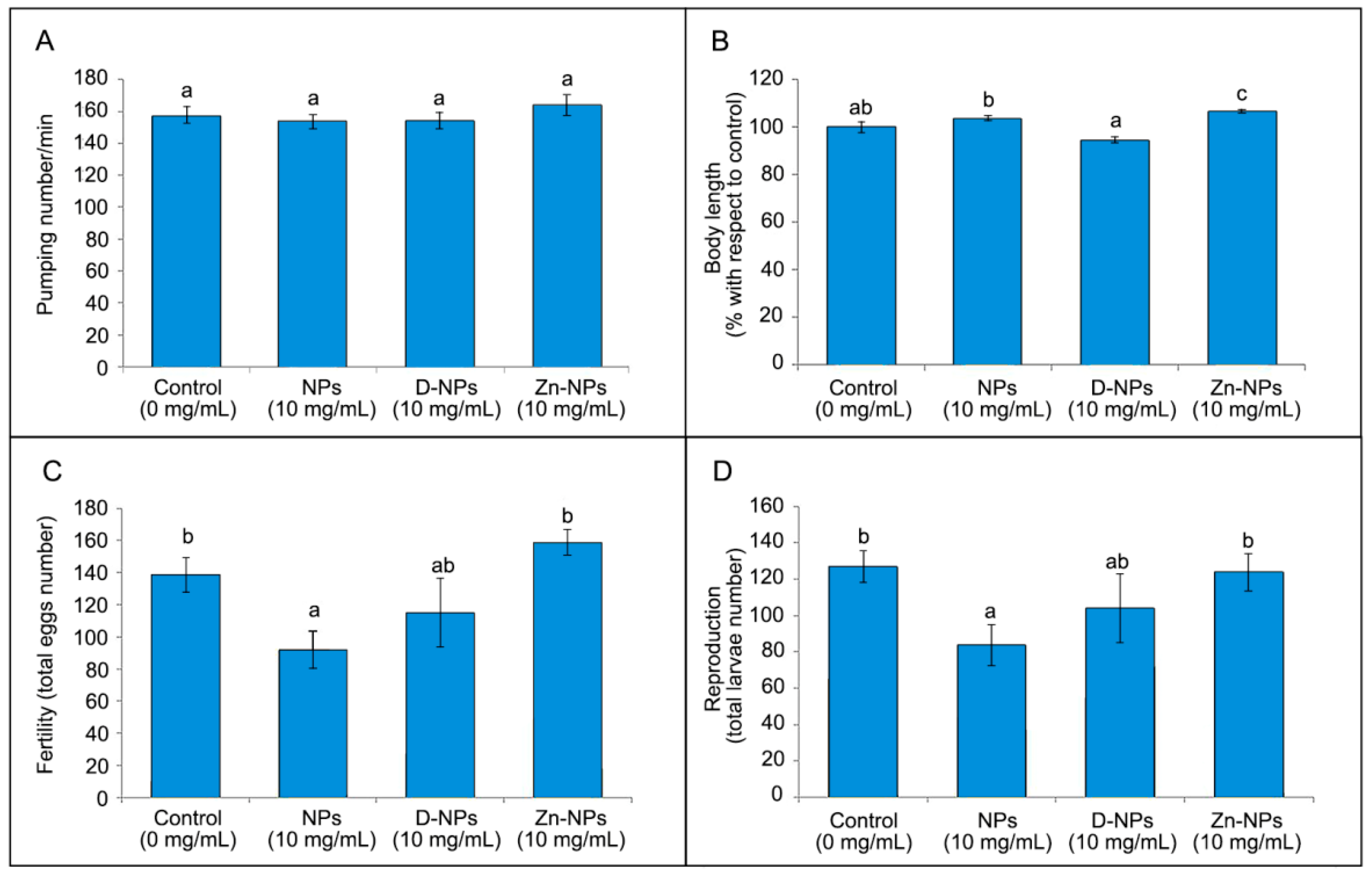
3.4. Pharyngeal Pumping Assay
3.5. Growth Assay
3.6. Reproduction and Fertility Assays
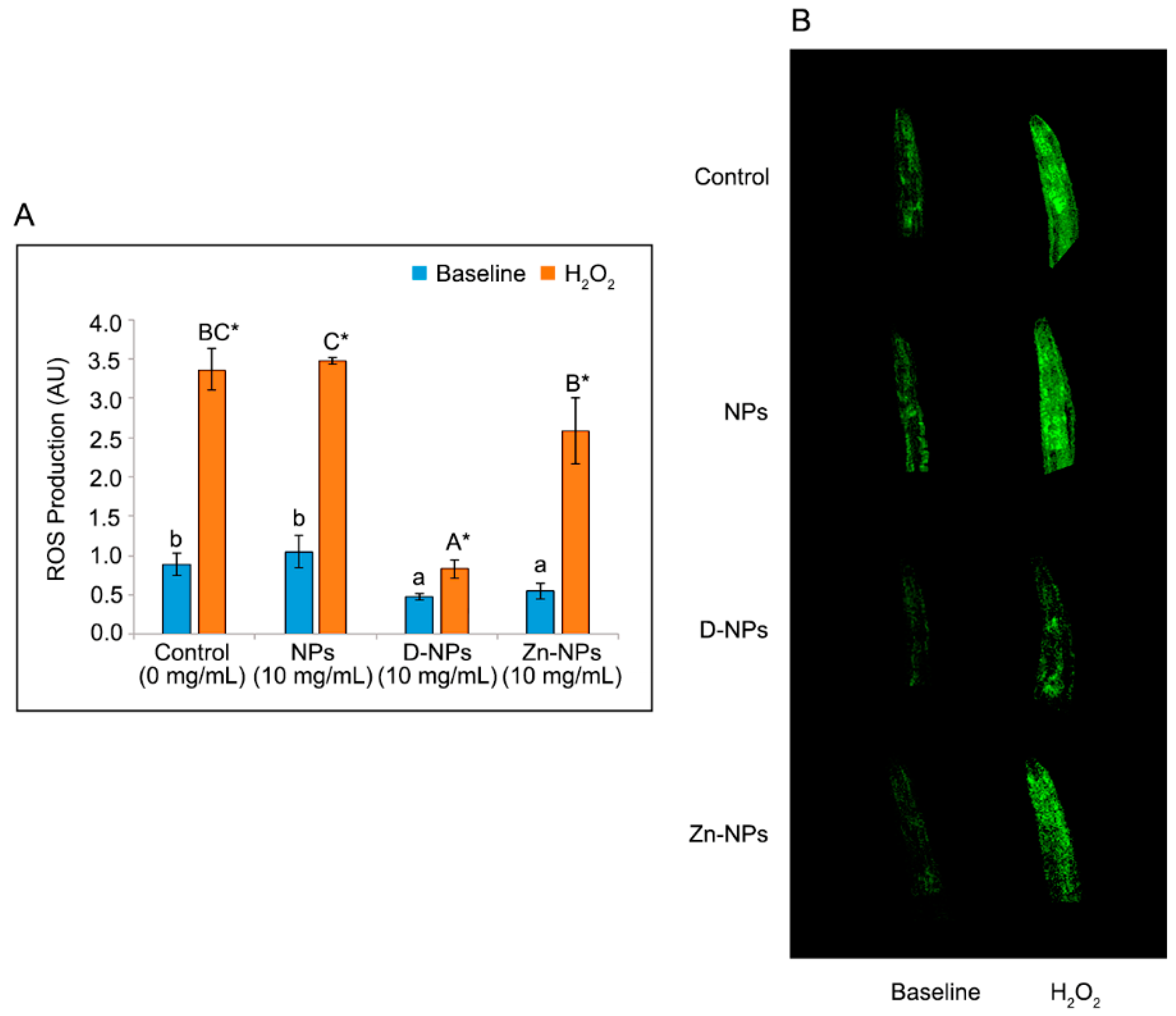
3.7. ROS Production
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Cho, K.; Wang, X.; Nie, S.; Chen, Z.G.; Shin, D.M. Therapeutic Nanoparticles for Drug Delivery in Cancer. Clin. Cancer Res. 2008, 14, 1310–1316. [Google Scholar] [CrossRef] [PubMed]
- Negi, H.; Saikia, S.K.; Kanaujia, R.; Jaiswal, S.; Pandey, R. 3β-Hydroxy-urs-12-en-28-oic acid confers protection against ZnONPs induced adversity in Caenorhabditis elegans. Environ. Toxicol. Pharmacol. 2017, 53, 105–110. [Google Scholar] [CrossRef] [PubMed]
- Ma, H.; Bertsch, P.M.; Glenn, T.C.; Kabengi, N.J.; Williams, P.L. Toxicity of manufactured zinc oxide nanoparticles in the nematode Caenorhabditis elegans. Environ. Toxicol. Chem. 2009, 28, 1324–1330. [Google Scholar] [CrossRef] [PubMed]
- Kamaly, N.; Yameen, B.; Wu, J.; Farokhzad, O.C. Degradable Controlled-Release Polymers and Polymeric Nanoparticles: Mechanisms of Controlling Drug Release. Chem. Rev. 2016, 116, 2602–2663. [Google Scholar] [CrossRef] [PubMed]
- Uhrich, K.E.; Cannizzaro, S.M.; Langer, R.S.; Shakesheff, K.M. Polymeric Systems for Controlled Drug Release. Chem. Rev. 1999, 99, 3181–3198. [Google Scholar] [CrossRef]
- Sonane, M.; Moin, N.; Satish, A. The role of antioxidants in attenuation of Caenorhabditis elegans lethality on exposure to TiO2 and ZnO nanoparticles. Chemosphere 2017, 187, 240–247. [Google Scholar] [CrossRef]
- Feuser, P.E.; Gaspar, P.C.; Ricci-Júnior, E.; da Silva, M.C.S.; Nele, M.; Sayer, C.; Araújo, P.H.H. de Synthesis and Characterization of Poly(Methyl Methacrylate) PMMA and Evaluation of Cytotoxicity for Biomedical Application. Macromol. Symp. 2014, 343, 65–69. [Google Scholar] [CrossRef]
- Medina-Castillo, A.L.; Fernandez-Sanchez, J.F.; Segura-Carretero, A.; Fernandez-Gutierrez, A. Micrometer and Submicrometer Particles Prepared by Precipitation Polymerization: Thermodynamic Model and Experimental Evidence of the Relation between Flory’s Parameter and Particle Size. Macromolecules 2010, 43, 5804–5813. [Google Scholar] [CrossRef]
- Kim, T.-S.; Bürklin, T.; Schacher, B.; Ratka-Krüger, P.; Schaecken, M.T.; Renggli, H.H.; Fiehn, W.; Eickholz, P. Pharmacokinetic Profile of a Locally Administered Doxycycline Gel in Crevicular Fluid, Blood, and Saliva. J. Periodontol. 2002, 73, 1285–1291. [Google Scholar] [CrossRef]
- Riba, A.; Deres, L.; Eros, K.; Szabo, A.; Magyar, K.; Sumegi, B.; Toth, K.; Halmosi, R.; Szabados, E. Doxycycline protects against ROS-induced mitochondrial fragmentation and ISO-induced heart failure. PLoS ONE 2017, 12. [Google Scholar] [CrossRef]
- Osorio, R.; Yamauti, M.; Osorio, E.; Ruiz-Requena, M.E.; Pashley, D.H.; Tay, F.R.; Toledano, M. Zinc reduces collagen degradation in demineralized human dentin explants. J. Dent. 2011, 39, 148–153. [Google Scholar] [CrossRef] [PubMed]
- Prasad, A.S. Clinical, immunological, anti-inflammatory and antioxidant roles of zinc. Exp. Gerontol. 2008, 43, 370–377. [Google Scholar] [CrossRef] [PubMed]
- Clemens, D.L.; Duryee, M.J.; Sarmiento, C.; Chiou, A.; McGowan, J.D.; Hunter, C.D.; Schlichte, S.L.; Tian, J.; Klassen, L.W.; O’Dell, J.R.; et al. Novel Antioxidant Properties of Doxycycline. Int. J. Mol. Sci. 2018, 19, 4078. [Google Scholar] [CrossRef]
- Marreiro, D.D.N.; Cruz, K.J.C.; Morais, J.B.S.; Beserra, J.B.; Severo, J.S.; de Oliveira, A.R.S. Zinc and Oxidative Stress: Current Mechanisms. Antioxidants 2017, 6, 24. [Google Scholar] [CrossRef]
- Dietrich, N.; Tan, C.-H.; Cubillas, C.; Earley, B.J.; Kornfeld, K. Insights into zinc and cadmium biology in the nematode Caenorhabditis elegans. Arch. Biochem. Biophys. 2016, 611, 120–133. [Google Scholar] [CrossRef] [PubMed]
- Donkin, S.G.; Ohlson, D.L.; Teaf, C.M. Properties and Effects of Metals. In Principles of Toxicology; Williams, P., James, R., Roberts, S., Eds.; John Wiley & Sons: New York, NY, USA, 2003; pp. 325–344. [Google Scholar]
- Misra, R.; Sahoo, S.K. Antibacterial activity of doxycycline-loaded nanoparticles. Methods Enzymol. 2012, 509, 61–85. [Google Scholar] [CrossRef] [PubMed]
- Ranjan, A.; Pothayee, N.; Seleem, M.N.; Tyler, R.D.; Brenseke, B.; Sriranganathan, N.; Riffle, J.S.; Kasimanickam, R. Antibacterial efficacy of core-shell nanostructures encapsulating gentamicin against an in vivo intracellular Salmonella model. Int. J. Nanomed. 2009, 4, 289–297. [Google Scholar] [CrossRef]
- Osorio, R.; Alfonso-Rodríguez, C.A.; Medina-Castillo, A.L.; Alaminos, M.; Toledano, M. Bioactive Polymeric Nanoparticles for Periodontal Therapy. PLoS ONE 2016, 11, e0166217. [Google Scholar] [CrossRef]
- Toledano-Osorio, M.; Babu, J.P.; Osorio, R.; Medina-Castillo, A.L.; García-Godoy, F.; Toledano, M. Modified Polymeric Nanoparticles Exert In Vitro Antimicrobial Activity Against Oral Bacteria. Materials 2018, 11, 1013. [Google Scholar] [CrossRef]
- Sánchez, M.C.; Toledano-Osorio, M.; Bueno, J.; Figuero, E.; Toledano, M.; Medina-Castillo, A.L.; Osorio, R.; Herrera, D.; Sanz, M. Antibacterial effects of polymeric PolymP-N Active nanoparticles. An in vitro biofilm study. Dent. Mater. 2019, 35, 156–168. [Google Scholar] [CrossRef]
- Toledano, M.; Cabello, I.; Osorio, E.; Aguilera, F.S.; Medina-Castillo, A.L.; Toledano-Osorio, M.; Osorio, R. Zn-containing polymer nanogels promote cervical dentin remineralization. Clin. Oral Investig. 2019, 23, 1197–1208. [Google Scholar] [CrossRef] [PubMed]
- Agarwal, A.; Bhattacharya, H.S.; Srikanth, G.; Singh, A. Comparative evaluation of decalcified freeze dried bone allograft with and without local doxycycline in non-contained human periodontal infrabony defects. J. Indian Soc. Periodontol. 2013, 17, 490–494. [Google Scholar] [CrossRef] [PubMed]
- Liss, K.-D.; Bartels, A.; Schreyer, A.; Clemens, H. High-energy X-rays: A tool for advanced bulk investigations in materials science and physics. Textures Microstruct. 2003, 35, 219–252. [Google Scholar] [CrossRef]
- Toledano, M.; Osorio, R.; Osorio, E.; Medina-Castillo, A.L.; Toledano-Osorio, M.; Aguilera, F.S. Ions-modified nanoparticles affect functional remineralization and energy dissipation through the resin-dentin interface. J. Mech. Behav. Biomed. Mater. 2017, 68, 62–79. [Google Scholar] [CrossRef] [PubMed]
- Chaturvedi, R.; Gill, A.S.; Sikri, P. Evaluation of the regenerative potential of 25% doxycycline-loaded biodegradable membrane vs biodegradable membrane alone in the treatment of human periodontal infrabony defects: a clinical and radiological study. Indian J. Dent. Res. 2008, 19, 116–123. [Google Scholar] [CrossRef] [PubMed]
- Smith, G.N.; Mickler, E.A.; Hasty, K.A.; Brandt, K.D. Specificity of inhibition of matrix metalloproteinase activity by doxycycline: Relationship to structure of the enzyme. Arthritis Rheum. 1999, 42, 1140–1146. [Google Scholar] [CrossRef]
- Beekman, B.; Verzijl, N.; de Roos, J.A.; Koopman, J.L.; TeKoppele, J.M. Doxycycline inhibits collagen synthesis by bovine chondrocytes cultured in alginate. Biochem. Biophys. Res. Commun. 1997, 237, 107–110. [Google Scholar] [CrossRef]
- Zhou, Y.; Wang, Q.; Song, B.; Wu, S.; Su, Y.; Zhang, H.; He, Y. A real-time documentation and mechanistic investigation of quantum dots-induced autophagy in live Caenorhabditis elegans. Biomaterials 2015, 72, 38–48. [Google Scholar] [CrossRef]
- Jiang, Y.; Chen, J.; Wu, Y.; Wang, Q.; Li, H. Sublethal Toxicity Endpoints of Heavy Metals to the Nematode Caenorhabditis elegans. PLoS ONE 2016, 11, e0148014. [Google Scholar] [CrossRef]
- Wu, T.; Xu, H.; Liang, X.; Tang, M. Caenorhabditis elegans as a complete model organism for biosafety assessments of nanoparticles. Chemosphere 2019, 221, 708–726. [Google Scholar] [CrossRef]
- Wang, Q.; Zhou, Y.; Song, B.; Zhong, Y.; Wu, S.; Cui, R.; Cong, H.; Su, Y.; Zhang, H.; He, Y. Linking Subcellular Disturbance to Physiological Behavior and Toxicity Induced by Quantum Dots in Caenorhabditis elegans. Small 2016, 12, 3143–3154. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Jiang, Y.; Xu, C.; Yu, L.; Sun, D.; Xu, L.; Hu, F.; Li, H. Comparison of two mathematical prediction models in assessing the toxicity of heavy metal mixtures to the feeding of the nematode Caenorhabditis elegans. Ecotoxicol. Environ. Saf. 2013, 94, 73–79. [Google Scholar] [CrossRef] [PubMed]
- Wu, Q.; He, K.; Liu, P.; Li, Y.; Wang, D. Association of oxidative stress with the formation of reproductive toxicity from mercury exposure on hermaphrodite nematode Caenorhabditis elegans. Environ. Toxicol. Pharmacol. 2011, 32, 175–184. [Google Scholar] [CrossRef] [PubMed]
- Novo, E.; Parola, M. Redox mechanisms in hepatic chronic wound healing and fibrogenesis. Fibrogenesis Tissue Repair 2008, 1, 5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, G.; Roy, I.; Yang, C.; Prasad, P.N. Nanochemistry and Nanomedicine for Nanoparticle-based Diagnostics and Therapy. Chem. Rev. 2016, 116, 2826–2885. [Google Scholar] [CrossRef] [PubMed]
- Bhattarai, G.; Poudel, S.B.; Kook, S.-H.; Lee, J.-C. Anti-inflammatory, anti-osteoclastic, and antioxidant activities of genistein protect against alveolar bone loss and periodontal tissue degradation in a mouse model of periodontitis. J. Biomed. Mater. Res. A 2017, 105, 2510–2521. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.H.; Kim, H.S.; Kim, J.S.; Yu, M.K.; Cho, S.D.; Jeon, J.G.; Yi, H.K. C-myb Regulates Autophagy for Pulp Vitality in Glucose Oxidative Stress. J. Dent. Res. 2016, 95, 430–438. [Google Scholar] [CrossRef]
- Toledano-Osorio, M.; Osorio, E.; Aguilera, F.S.; Medina-Castillo, A.L.; Toledano, M.; Osorio, R. Improved reactive nanoparticles to treat dentin hypersensitivity. Acta Biomater. 2018, 72, 371–380. [Google Scholar] [CrossRef]
- Corsi, A.K.; Wightman, B.; Chalfie, M. A Transparent Window into Biology: A Primer on Caenorhabditis elegans. Genetics 2015, 200, 387–407. [Google Scholar] [CrossRef] [Green Version]
- Wu, Q.; Nouara, A.; Li, Y.; Zhang, M.; Wang, W.; Tang, M.; Ye, B.; Ding, J.; Wang, D. Comparison of toxicities from three metal oxide nanoparticles at environmental relevant concentrations in nematode Caenorhabditis elegans. Chemosphere 2013, 90, 1123–1131. [Google Scholar] [CrossRef]
- Fang, E.F.; Waltz, T.B.; Kassahun, H.; Lu, Q.; Kerr, J.S.; Morevati, M.; Fivenson, E.M.; Wollman, B.N.; Marosi, K.; Wilson, M.A.; et al. Tomatidine enhances lifespan and healthspan in C. elegans through mitophagy induction via the SKN-1/Nrf2 pathway. Sci. Rep. 2017, 7, 46208. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, F.; Zaman, W.Q.; Peng, H.; Li, C.; Cao, X.; Huang, K.; Cui, C.; Zhang, W.; Lin, K.; Luo, Q. Ecotoxicity of Caenorhabditis elegans following a step and repeated chronic exposure to tetrabromobisphenol A. Ecotoxicol. Environ. Saf. 2019, 169, 273–281. [Google Scholar] [CrossRef] [PubMed]
- Lei, L.; Wu, S.; Lu, S.; Liu, M.; Song, Y.; Fu, Z.; Shi, H.; Raley-Susman, K.M.; He, D. Microplastic particles cause intestinal damage and other adverse effects in zebrafish Danio rerio and nematode Caenorhabditis elegans. Sci. Total Environ. 2018, 619–620, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Back, P.; De Vos, W.H.; Depuydt, G.G.; Matthijssens, F.; Vanfleteren, J.R.; Braeckman, B.P. Exploring real-time in vivo redox biology of developing and aging Caenorhabditis elegans. Free Radic. Biol. Med. 2012, 52, 850–859. [Google Scholar] [CrossRef]
- Belousov, V.V.; Fradkov, A.F.; Lukyanov, K.A.; Staroverov, D.B.; Shakhbazov, K.S.; Terskikh, A.V.; Lukyanov, S. Genetically encoded fluorescent indicator for intracellular hydrogen peroxide. Nat. Methods 2006, 3, 281–286. [Google Scholar] [CrossRef]
- Fang-Yen, C.; Avery, L.; Samuel, A.D.T. Two size-selective mechanisms specifically trap bacteria-sized food particles in Caenorhabditis elegans. Proc. Natl. Acad. Sci. USA 2009, 106, 20093–20096. [Google Scholar] [CrossRef] [Green Version]
- Kong, L.; Gao, X.; Zhu, J.; Zhang, T.; Xue, Y.; Tang, M. Reproductive toxicity induced by nickel nanoparticles in Caenorhabditis elegans. Environ. Toxicol. 2017, 32, 1530–1538. [Google Scholar] [CrossRef]
- Gupta, S.; Kushwah, T.; Vishwakarma, A.; Yadav, S. Optimization of ZnO-NPs to Investigate Their Safe Application by Assessing Their Effect on Soil Nematode Caenorhabditis elegans. Nanoscale Res. Lett. 2015, 10, 1010. [Google Scholar] [CrossRef] [Green Version]
- Ahmed, S.; Annu; Chaudhry, S.A.; Ikram, S. A review on biogenic synthesis of ZnO nanoparticles using plant extracts and microbes: A prospect towards green chemistry. J. Photochem. Photobiol. B 2017, 166, 272–284. [Google Scholar] [CrossRef]
- Moyson, S.; Vissenberg, K.; Fransen, E.; Blust, R.; Husson, S.J. Mixture effects of copper, cadmium, and zinc on mortality and behavior of Caenorhabditis elegans. Environ. Toxicol. Chem. 2018, 37, 145–159. [Google Scholar] [CrossRef]
- Kim, Y.; Jeong, J.; Yang, J.; Joo, S.-W.; Hong, J.; Choi, J. Graphene oxide nano-bio interaction induces inhibition of spermatogenesis and disturbance of fatty acid metabolism in the nematode Caenorhabditis elegans. Toxicology 2018, 410, 83–95. [Google Scholar] [CrossRef] [PubMed]
- Hoppe, A.; Güldal, N.S.; Boccaccini, A.R. A review of the biological response to ionic dissolution products from bioactive glasses and glass-ceramics. Biomaterials 2011, 32, 2757–2774. [Google Scholar] [CrossRef] [PubMed]
- Gao, H.; Dai, W.; Zhao, L.; Min, J.; Wang, F. The Role of Zinc and Zinc Homeostasis in Macrophage Function. J. Immunol. Res. 2018, 2018, 6872621. [Google Scholar] [CrossRef] [PubMed]
- Lopes, M.M.G.D.; de Brito, N.J.N.; de Medeiros Rocha, É.D.; França, M.C.; de Almeida, M.D.G.; Brandão-Neto, J. Nutritional assessment methods for zinc supplementation in prepubertal non-zinc-deficient children. Food Nutr. Res. 2015, 59, 29733. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cornaglia, M.; Krishnamani, G.; Mouchiroud, L.; Sorrentino, V.; Lehnert, T.; Auwerx, J.; Gijs, M.A.M. Automated longitudinal monitoring of in vivo protein aggregation in neurodegenerative disease C. elegans models. Mol. Neurodegener. 2016, 11, 17. [Google Scholar] [CrossRef] [Green Version]
- Letizia, M.C.; Cornaglia, M.; Tranchida, G.; Trouillon, R.; Gijs, M.A.M. A design of experiment approach for efficient multi-parametric drug testing using a Caenorhabditis elegans model. Integr. Biol. 2018, 10, 48–56. [Google Scholar] [CrossRef]
- Wolf, T.; Qi, W.; Schindler, V.; Runkel, E.D.; Baumeister, R. Doxycyclin ameliorates a starvation-induced germline tumor in C. elegans daf-18/PTEN mutant background. Exp. Gerontol. 2014, 56, 114–122. [Google Scholar] [CrossRef]
- Tsang, W.Y.; Lemire, B.D. Mitochondrial Genome Content Is Regulated during Nematode Development. Biochem. Biophys. Res. Commun. 2002, 291, 8–16. [Google Scholar] [CrossRef]
- Suri, S.S.; Fenniri, H.; Singh, B. Nanotechnology-based drug delivery systems. J. Occup. Med. Toxicol. 2007, 2, 16. [Google Scholar] [CrossRef] [Green Version]
- Waddington, R.J.; Moseley, R.; Embery, G. Reactive oxygen species: A potential role in the pathogenesis of periodontal diseases. Oral Dis. 2000, 6, 138–151. [Google Scholar] [CrossRef]
- Xu, Y.; Park, Y. Application of Caenorhabditis elegans for Research on Endoplasmic Reticulum Stress. Prev. Nutr. Food Sci. 2018, 23, 275–281. [Google Scholar] [CrossRef] [PubMed]





© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Toledano, M.; Toledano-Osorio, M.; Navarro-Hortal, M.D.; Varela-López, A.; Osorio, R.; Quiles, J.L. Novel Polymeric Nanocarriers Reduced Zinc and Doxycycline Toxicity in the Nematode Caenorhabditis elegans. Antioxidants 2019, 8, 550. https://doi.org/10.3390/antiox8110550
Toledano M, Toledano-Osorio M, Navarro-Hortal MD, Varela-López A, Osorio R, Quiles JL. Novel Polymeric Nanocarriers Reduced Zinc and Doxycycline Toxicity in the Nematode Caenorhabditis elegans. Antioxidants. 2019; 8(11):550. https://doi.org/10.3390/antiox8110550
Chicago/Turabian StyleToledano, Manuel, Manuel Toledano-Osorio, María D. Navarro-Hortal, Alfonso Varela-López, Raquel Osorio, and José L. Quiles. 2019. "Novel Polymeric Nanocarriers Reduced Zinc and Doxycycline Toxicity in the Nematode Caenorhabditis elegans" Antioxidants 8, no. 11: 550. https://doi.org/10.3390/antiox8110550
APA StyleToledano, M., Toledano-Osorio, M., Navarro-Hortal, M. D., Varela-López, A., Osorio, R., & Quiles, J. L. (2019). Novel Polymeric Nanocarriers Reduced Zinc and Doxycycline Toxicity in the Nematode Caenorhabditis elegans. Antioxidants, 8(11), 550. https://doi.org/10.3390/antiox8110550