1. Introduction
Biodiversity loss is one of the problems the world is facing nowadays. It is declining worldwide due to different reasons: global warming, overexploitation, changes in land and sea use, habitat destruction, and bioinvasions [
1,
2,
3]. Bioinvasions are the introduction of non-indigenous species (hereafter NIS) in a different biogeographic region due to human activity, in both direct and indirect ways. In fact, the introduction of NIS and habitat modification are considered the most important drivers of biodiversity loss in coastal areas [
4,
5]. Almost all seas of the world have experienced the introduction of NIS [
6,
7,
8], with the Mediterranean Sea being the most invaded one, with over 700 NIS [
9,
10].
During recent years, studies about NIS have increased, but studies focusing on the variations in temporal recruitment and ecological mechanisms are still scarce [
11,
12]. NIS has several effects on the host habitat, as they compete with native species for resources, such as food and space, and they can change the stablished community structure and alter the ecosystem functions by the substitution of species, up to the point of eliminating the native species [
13,
14]. Hence, they promote the biotic homogenization of biodiversity and functionality [
13,
15,
16].
Biofouling and ballast waters are among the main human-mediated dispersal pathways for aquatic NIS, which make port areas, especially large ports, hotspots for the primary introduction of NIS [
16,
17]. In fact, those areas are the ones with a high number of human-made artificial infrastructures submerged in the marine environment. Those structures are made of both natural and artificial materials, but the artificial ones are more common, such as pontoons, buoys, and dikes. It has been seen that there are differences in biofouling assemblages within different substrates (e.g., [
18,
19,
20]), and one of the reasons is that NIS are more frequent in artificial substrates than in naturally occurring ones [
20,
21,
22]. As such, these artificial structures act as points of entry for NIS in port areas and may support more abundance and richness of them [
23,
24,
25]. This is because artificial materials and port areas are an anthropologically altered environment, in which NIS can compete better than native ones, contrary to what happens in natural habitats [
14,
20,
24].
In this context, there is an urgent need to determine the ecological and evolutionary mechanisms that promote NIS in order to make appropriate management and conservation strategies [
2]. This is especially important when new structures are submerged in the marine environment, as it is aimed to recruit the least possible number of NIS [
24,
26]. Moreover, given the important role of port structures in the introduction and establishment of NIS, these areas are pivotal to assess the early detection of NIS for further management strategies [
16].
Hence, we aimed to observe the colonization of sessile macrofouling from Alicante’s port on two types of substrates (electrolytic carbonated and steel) over 12 months. These materials were used as, contrary to concrete or PVC (the most commonly used materials), they do not have any negative impacts on the marine environment as they do not release any substances from them. In addition, as they are made out of the salts dissolved in the sea water, they are natural substrates and are similar in composition to natural limestone [
27]. The colonization stages of biofouling species on these types of substrates have been analyzed in a previous work [
28]. However, this study focused on the non-indigenous and cryptogenic species in order to observe how their development vary among substrates. It was hypothesized that the NIS would be more abundant on bare steel as it is a more artificial substrate. Moreover, we intended to observe the development and succession patterns of NIS within the different substrates.
4. Discussion
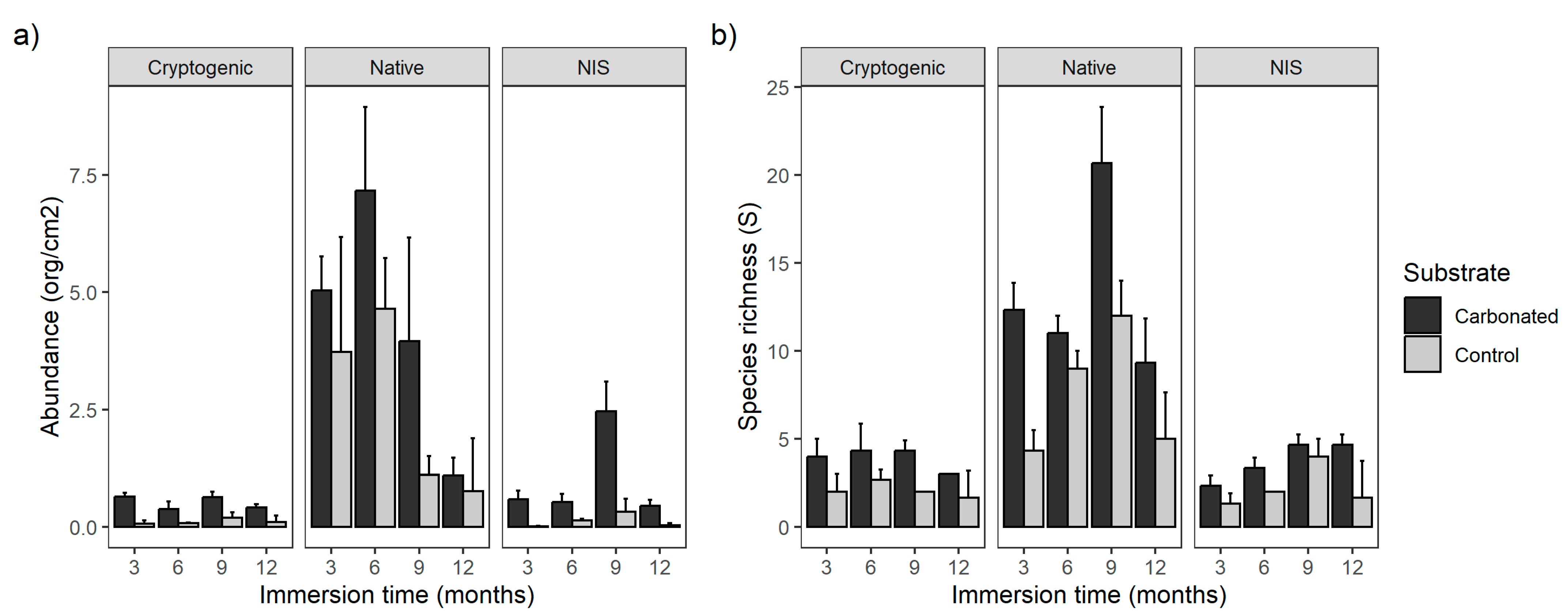
The abundance of NIS, native, and cryptogenic species was higher in the carbonated treatment than in the steel one. Moreover, it was influenced by the immersion time, especially for the native species and NIS. Those two were offset during the succession, as at 6 months the natives showed their maximum abundance and NIS decreased, but after 9 months NIS abundance increased almost up to the native values, and they kept almost the same relative abundance to each other in the last period, although both abundances decreased. Similarly to the abundance, the same occurred for the species richness, except both the native species and NIS had their higher number of species at the third period (9 months). These results show that species were clearly influenced by seasonal larval supply and seasonality [
12,
49,
50]. In fact, according to what was expected, the NIS had their highest abundance and species richness values after 9 months, from later spring to early–mid-summer (May–July), as they have been shown to have higher peaks in warmer months [
49,
51,
52]. Some authors, such as Fargione et al. [
53], have suggested that the success of NIS over natives is due to a non-overlapping recruitment period, but this trend does not have to be like the one experienced by [
12] in the Central Mediterranean. Indeed, in our case, the trend was not completely clear, as abundances did not overlap in time, whereas the peak of species richness for both natives and NIS occurred at the same period, so more studies should be carried out in order to determine the trend in the port of Alicante.
Referring to the effect of the substrate over the species, contrary to expectations, both abundance and species richness of NIS were higher in the carbonated treatment. This is peculiar as NIS have a preference or they can compete better in artificial substrates than native ones, and because the carbonated material is very similar to naturally occurring limestone, so it was expected to have more NIS in the control treatment. Nevertheless, by looking at the control treatment, it is clear that NIS became more abundant than natives in their peak of recruitment (third period), agreeing with other studies [
14,
20,
24]. The electrolytic carbonated substrate is so similar to natural rock that it imitates the home range substrate of a large number of species, independently of their status of introduction. This could explain the reason for having a high number of both natives and NIS, and why it enhanced the recruitment of them over the control treatment, where the expected trend occurred.
In port environments, it is not always clear what is the naturally occurring community as most of the substrates are artificial, it is an altered system [
54], and there are (when there are) few previous studies about the inhabiting species. For the case of Alicante’s port, there are a few studies, of which, we highlight a study from [
34,
44,
47], in which they found 10 NIS by rapid assessment protocol and both mobile and sessile macrofouling species were considered (
Styela plicata,
Celleporaria brunnea,
Watersipora arcuata,
Oculina patagonica,
Caprella scaura,
Mesanthura cf.
romulea,
Paranthura japonica,
Branchiomma bairdi,
Ficopomatus enigmaticus, and
Hydroides elegans). In our study, a similar number of NIS (seven spp.) were found, but with some differences, as we did not find
O. patagonica,
F. enigmaticus,
W. arcuata, and
C. brunnea, while
Spirobranchus cf.
tetraceros and
Botrylloides cf.
niger where not found in [
34,
44]. It has to be highlighted that
S.cf.
tetraceros was only recorded in Spain in the port of Valencia [
55] until this study. Although only one specimen has been found, it could mean that the population from Valencia is spreading, probably thanks to shipping, and it reached Alicante’s port. Moreover, this is the first record for
B. cf.
niger in the Spanish Mediterranean. However, one of the authors (AARE) probably detected this in 1979 and identified it as
Botrylloides leachii in the port of Alicante during summer months [
47]. Furthermore, there are some other new records for Alicante’s harbor that can have been overlooked due to the lack of studies, such as the case of
Hydroides dirampha that has been detected since 1923 in Valencia [
56], which could have arrived some years ago. Serpulids, Bryozoans, and Cirripedia are typical fouling species, and it is possible that the species recorded here have been in the study area for decades. In addition, it should be noted that cryptogenic and non-indigenous species abundance in the port of Alicante and recorded since the 1970s [
45,
47], such as the bryozoan
Amathia verticillata (delle Chiaje, 1822), and the ascidians
Clavelina lepadiformis (Müller, 1776),
Botryllus schlosseri (Pallas, 1766), and
Microcosmus squamiger Michaelsen, 1927 (as
M. exasperatus), have not been collected on the meshes (carbonated and steel). So, it shows the need for a greater effort in temporal monitoring of present NIS in order to prevent further bioinvasions and local spreading. In fact, this monitoring should be a standardized method to remove the bias.
Referring to each NIS, it can be seen that there are three groups: (i) the ones not influenced by any of the studied factors (
S. cf.
tetraceros), (ii) ones influenced by both factors and interaction (
B. bairdi,
B. trigonus,
S. plicata), and (iii) ones affected by immersion time and/or interaction (
H. dirampha and
H. elegans). As
Hydroides spp. are the ones that dominate worldwide fouling communities [
8,
57], it makes sense for these to be time-dependent more than substrate-material-dependent, as they are gregarious species and grow over other serpulid tubes. In fact,
H. elegans was only more abundant in the carbonated treatment in the first period, although it was abundant during the study, which means that it is a pioneer species, but it does not disappear. In the case of other species, this preference for the electrolytic carbonated substrate could be because of its similarity to limestone, as it has been said. Nevertheless, in all cases, the seasonality of species was demonstrated as most of them had their highest abundances after 9 months of immersion (third period), corresponding to warmer months. There was a case where high abundances were found, especially in the carbonated treatment, in the first months (
Balanus trignous and
H. elegans). Moreover, there were non-indigenous species that appeared only in the warmer periods, as in the case of
B. cf.
niger, and others appeared during the cold months and were maintained along the study (
B. bairdi and
S. plicata). This could be due to the propagule supply and because they need a specific development stage of the community.
Among cryptogenic species, Savignyella lafontii presented a specific development stage of the community as it only appeared in the last period, and Amphibalanus amphitrite, S. plicata, S. errata, and Watersipora subtorquata were influenced by the substrate and immersion length. A. amphitrite was more abundant in the carbonated treatment after 3 and 9 months, and after 12 months it did not appear alive, only the shell. This could mean that A. amphitrite acts as a pioneer species and, as the community develops it leaves the space to other species, such as S. lafontii, as it cannot compete against them. One of those species could be S. errata, which has a laminar growth and can grow over the shell, limiting or blocking its feeding capacity. In fact, this could be crucial in the development of the community, as it generates more available hard substrate for the colonization of other species, as happens with the shells of dead cirripedes. Moreover, S. errata was more abundant in the carbonated treatment for almost every sample, which goes along with the fact that this substrate has more space (carbonated layer plus other hard bodied organisms) to be colonized. Furthermore, W. subtorquata could have a similar behavior to S. errata as it also has this laminar growth, but this species seems to be influenced by the immersion period. Nevertheless, there are some species, mainly ascidians, that have not been recorded during the study and they are known for the port. This implies that these substrates are not suitable for their settlement, or they need a more developed community to correctly settle and develop.
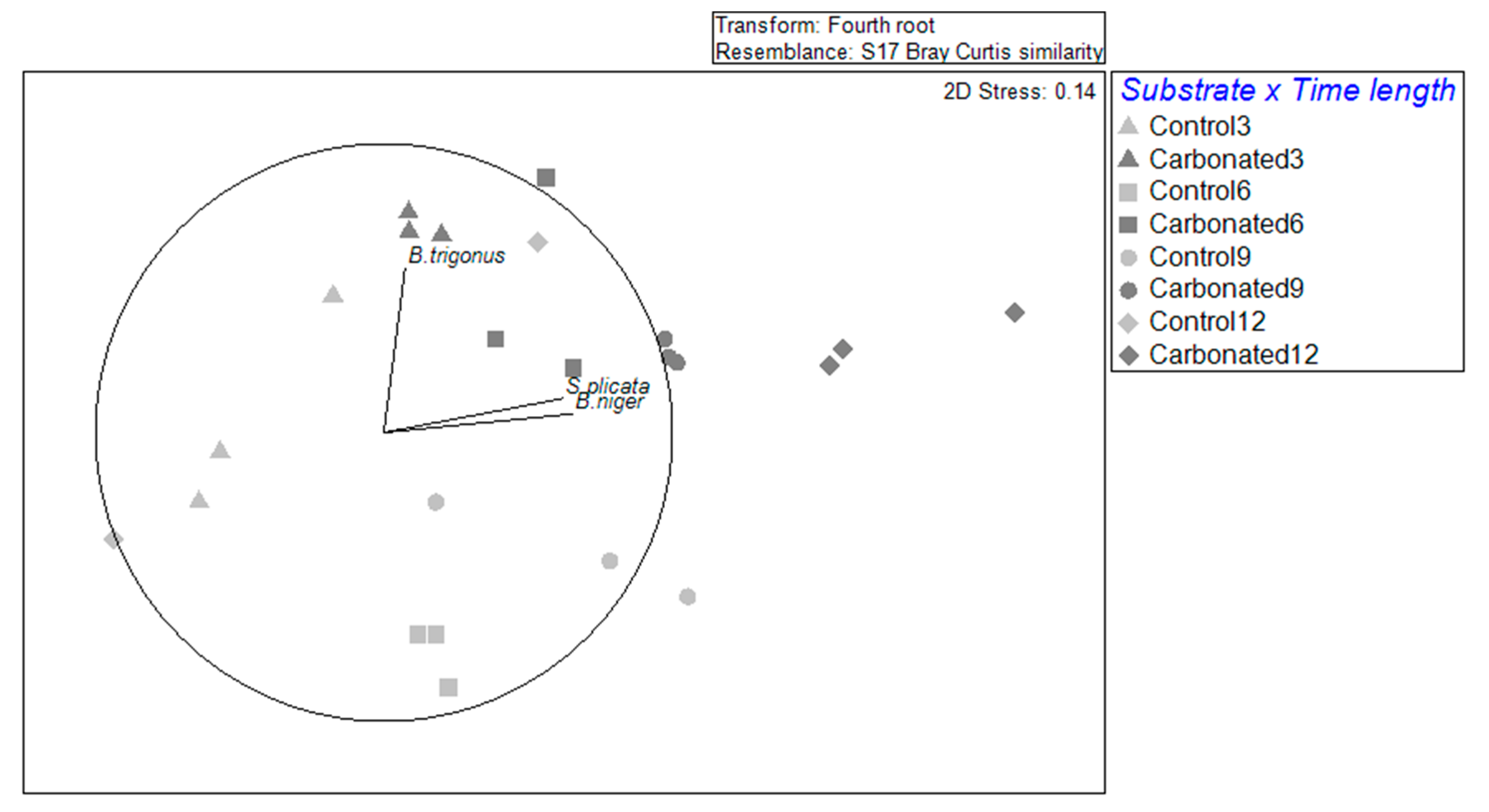
Regarding NIS assemblages, it was observed that they were differentiated by substrate and by immersion time, as there are two groups: 3 to 6 months and 9 to 12 months. This trend was especially marked in the control treatment, and it matched the separation of months by their temperature: the colder months and the warmer ones. In the carbonated treatment, this separation could be seen between the first three months and the last 9 and 12, being the stage of 6 months a transitory and dispersed time. These results go along with what was observed, studying the whole fouling community in these typed of substrates [
28]. In addition, it also suggests a development of the assemblages, which could be seen in the carbonated treatment, for which
B. trigonus will be a pioneer species and, as far as the immersion duration goes ahead, ascidians will appear, and they will be more important in that step of succession.
All non-indigenous species were positively correlated between them in the steel treatment. This result, and the fact that steel is an artificial substrate, will support the idea that NIS can enhance the recruitment of each other, being the reason for appearing as better colonizers than natives in artificial materials. On the other hand, in the carbonated treatment, there were species with negative correlations. These were B. cf. niger with B. trigonus. These negative correlations could be caused by the encrusting growth of B. cf. niger as it could grow over the barnacle, hindering its capacity to feed. In fact, B. cf. niger negatively correlated with the rest of the species, except for S. plicata, supporting the idea that its growth type prevents the growth, feeding, and/or settlement of other species.
In addition, all tube-forming species (order Sabellida) correlated together, which would indicate that the species belonging to the taxa tend to recruit together. These results could mean that they not only do not compete with each other for the space, but also, they can co-occur within the same habitat and enhance the recruitment of others. In fact, this was seen with
W. subtorquata in the carbonated treatment, which used the formed new substrate to grow, and with
S. plicata in the control treatment. In this last case, both taxa benefitted from each other, as the ascidian could use the new substrata and generate more substrate with its tunic to the growth of other species [
58]. Moreover, these species are the typical ones with high abundance and rapid growth (the case of serpulids, principally) in port areas, as well as other artificial substrates and perturbed areas [
57,
59], which could be a good explanation.
Taking all of this into account, most of the NIS have demonstrated a preference for the electrolytic carbonated substrate and the NIS that appeared are the typical ones in the fouling communities worldwide. It has been shown that the trend in artificial substrates in the recruitment of NIS is not always happening, so there is a need to determine which types of substrates recruit more NIS. In addition, the number of NIS present in the port of Alicante is increasing over the years and there is no control over it, so, there is a need for monitoring port environments in order to assess and to control new bioinvasions and the spreading of occurring NIS. Thus, as NIS preferred the electrolytic carbonated substrate, it would be a suitable material as an early detection tool of NIS in port areas. Actually, even though NIS were more abundant on it, native ones were too and the relative abundance between them was higher than in steel, so it can be considered a better construction material for those areas.


 ,
, 


{kind=link}
{kind=link}
{kind=link}