
Mitigating Fish-Killing Prymnesium parvum Algal Blooms in Aquaculture Ponds with Clay: The Importance of pH and Clay Type

 ,
, 

Abstract
:1. Introduction
2. Methods
2.1 Clay Materials

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Clay group | Clay code | Major clay type | Product name/Company | Cation exchange capacity (CEC) | Suppliers description |
|---|---|---|---|---|---|
| Bentonite | B1 | Ca-bentonite | B1-AM/Bentonite Products WA, Watheroo, Australia | N.A. | 50% A, 50% gypsum; low swelling |
| B2 | Ca-bentonite | Es/Bentonite Products WA, Watheroo, Australia | 95.70 | 39.02% clay, 9.44% coarse sand, 37.39% fine sand, <14.15% silt | |
| B3 | Modified Na-bentonite | Phoslock™/Phoslock Water Solutions Ltd, Sydney, Australia | N.A. | Lanthanum-modified bentonite (5% La3+) | |
| B4 | Ca-bentonite | A/Bentonite Products WA, Watheroo, Australia | 64.54 | 81.9% clay, 2.2% coarse sand, 16% fine sand, <0.01% silt | |
| B5 | Ca-bentonite | Ed/Bentonite Products WA, Watheroo, Australia | 85.80 | 83.4% clay, 4.4% coarse sand, 9.8% fine sand, <2.5% silt | |
| B6 | Na-bentonite | Trubond/Sibelco, Sydney, Australia | 82 | Montmorillonite as a major component | |
| Kaolin | K1 | Kaolin | Fine ceramic clay obtained from potter, Hobart, Australia | N.A. | N.A. |
| K2 | Kaolin | CA-1/Claypro, Junortoun, Australia | N.A. | Dry milled ball clay (kaolinite as a major component) | |
| K3 | Kaolin | Kaolinite/Zeolite Australia, Werris Creek, Australia | 29–39 | 60%–75% opaline silica, 20%–30% kaolinite, 5% other clays and mica | |
| K4 | Kaolin | Snobrite C/Sibelco, Sydney, Australia | N.A. | 70%–95% kaolinite, 5%–15% quartz, <15% other minerals | |
| Zeolites | Z1 | Zeolite | Zeolite/Zeolite Australia, Werris Creek, Australia | >120 | Clinoptotilite with minor amounts of quartz, smectite |
| Z2 | Zeolite | ANZ38/Castle Mountain Zeolites, Quirindi, Australia | 147 | 85% clinoptotilite, 15% mordenite), minor amounts of quartz, feldspar, montmorillonite | |
| Korean loess | Kor 1 | Korea A | Sourced from Tongyeong, South Korea | N.A. | Korean loess |
| Kor 2 | Korea B | Sourced from Geoje, South Korea | N.A. | Korean loess |
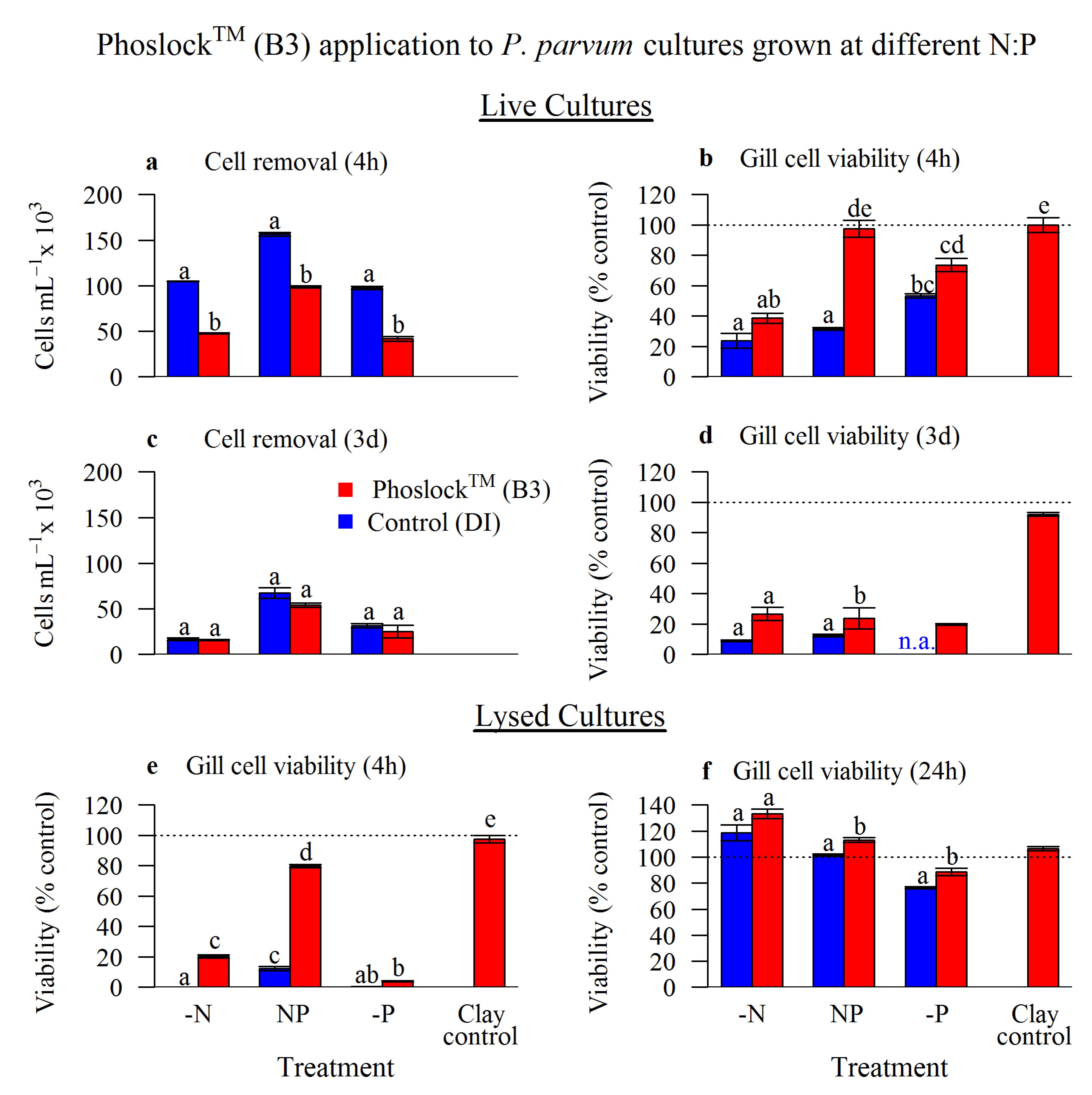
2.2. Phoslock™ (B3) Application
2.2.1. Live Cultures
2.2.2. Lysed Cultures
2.2.3. High pH Culture
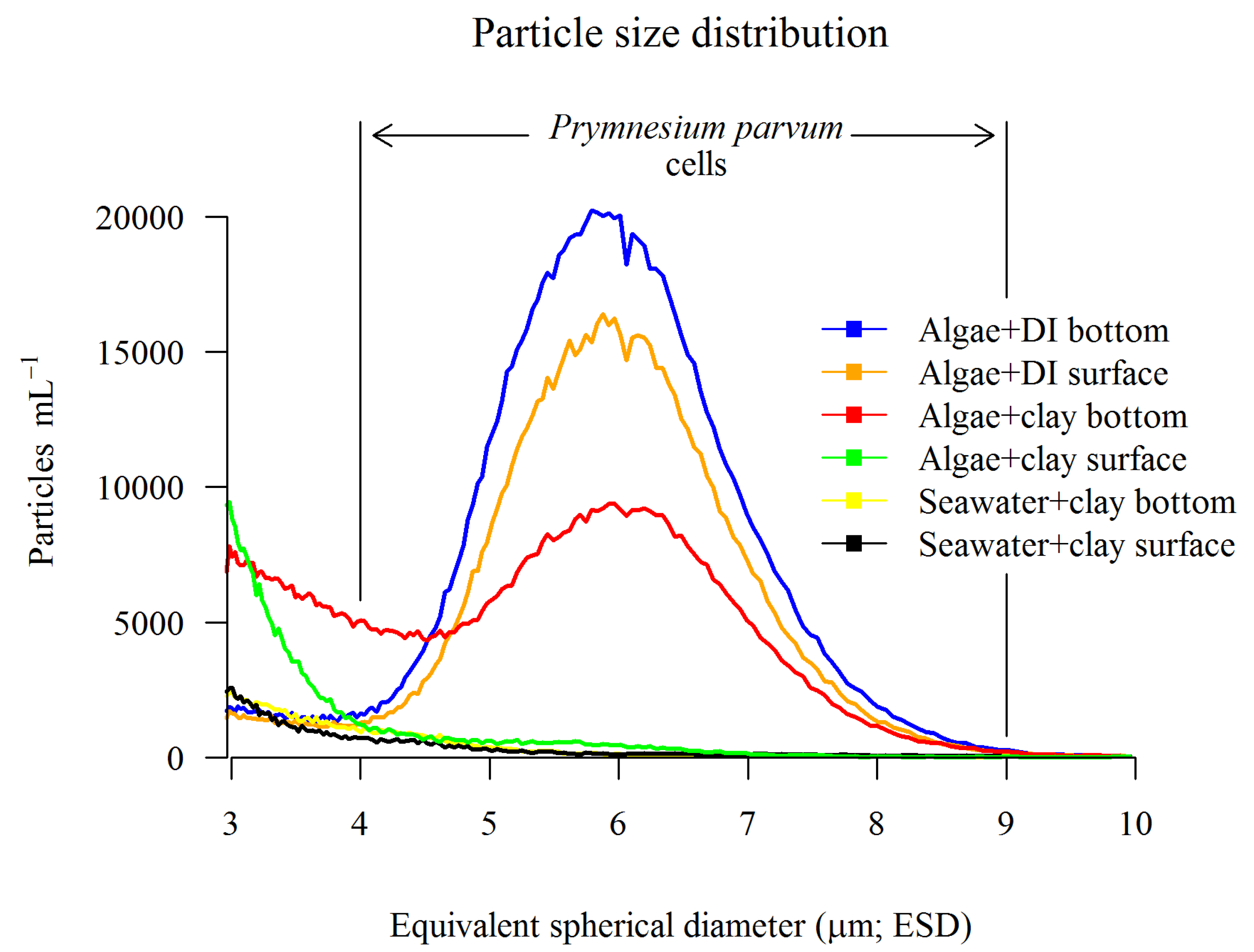
2.3. Clay Screening
2.3.1. Cell Removal
2.3.2. Ichthyotoxin Removal
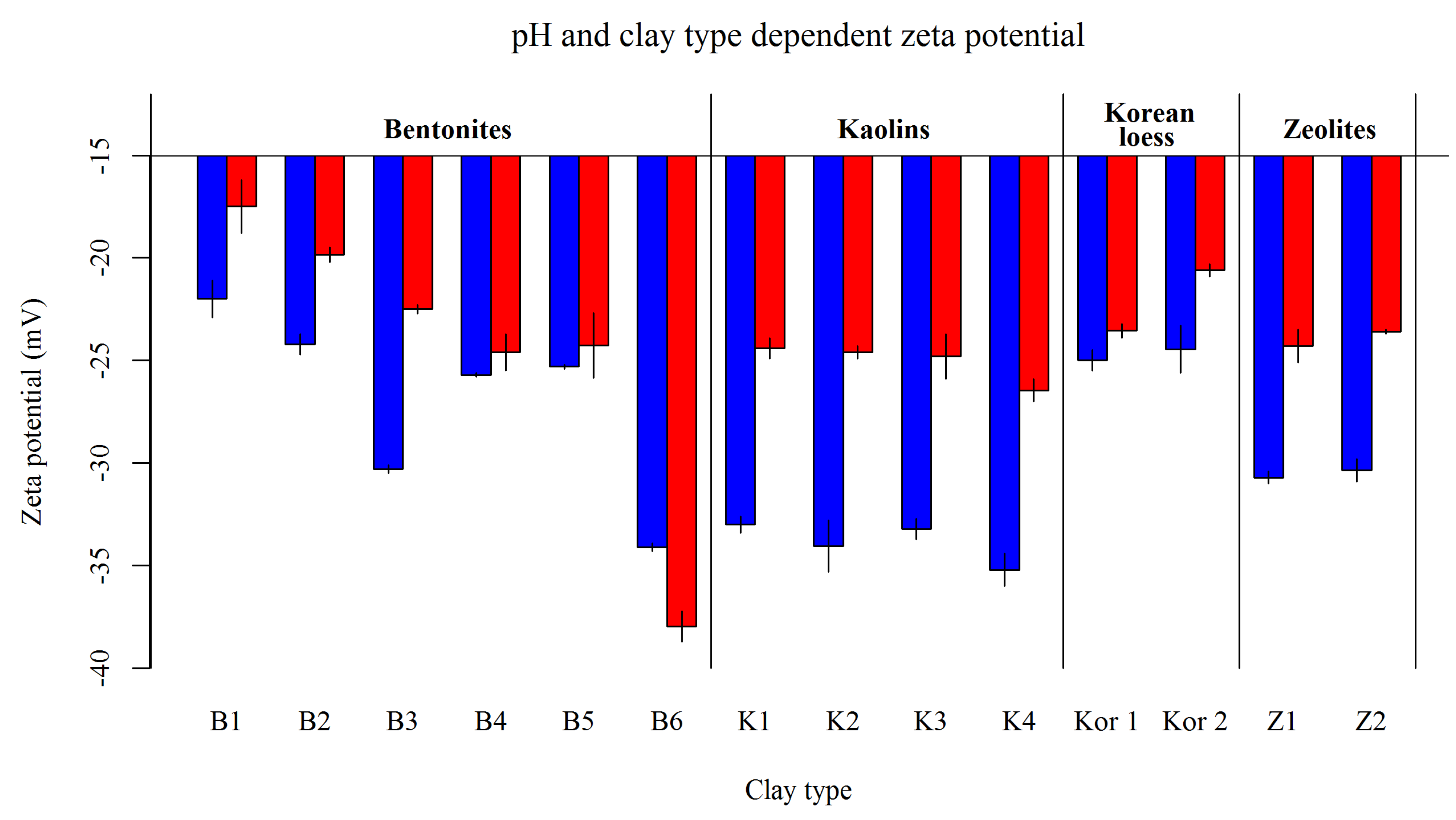
2.3.3. Zeta Potential
2.3.4. Minimum Effective Dose
2.4. Gill Cell Line Assay
2.5. Statistics
3. Results
3.1. Phoslock™ (B3) Application
3.1.1. Live Cultures

3.1.2. Lysed Cultures

3.1.3. High pH Culture

3.2. Clay Screening for Cell and Ichthyotoxin Removal
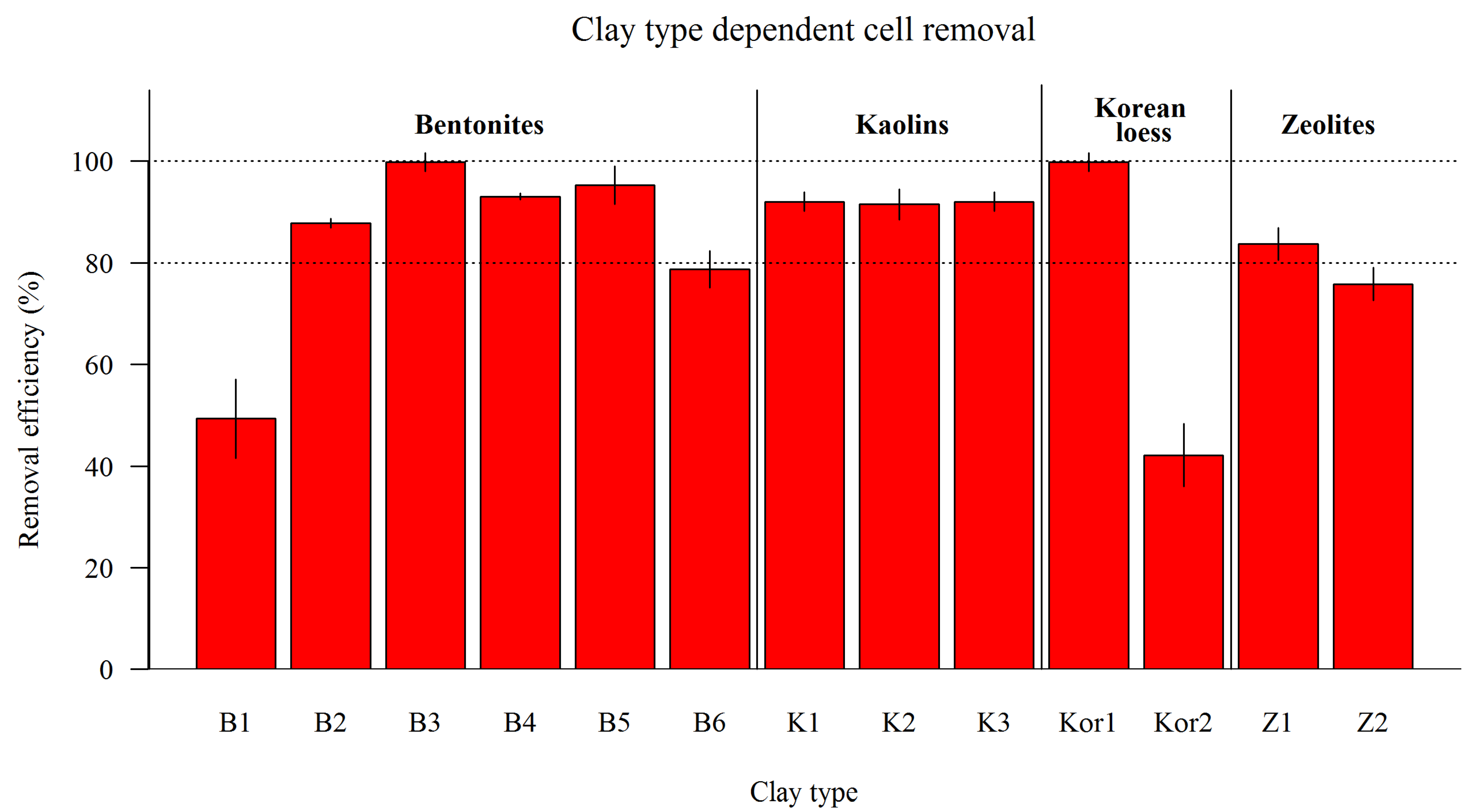
3.2.1. Cell Removal

3.2.2. Ichthyotoxin Removal
| Source | Df | SS | MS | F | p |
|---|---|---|---|---|---|
| Clay type | 13 | 452.40 | 34.80 | 227.22 | <0.001 |
| pH | 1 | 90.60 | 90.60 | 591.51 | <0.001 |
| Clay type * pH | 13 | 141.00 | 10.85 | 70.83 | <0.001 |
| Residuals | 56 | 8.60 | 0.15 |

3.2.3. Zeta Potential


3.2.4. Minimum Effective Dose
4. Discussion
4.1. Phoslock™ (B3) Application
4.2. Clay Screening
5. Conclusion
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Shilo, M.; Shilo, M. Conditions which determine the efficiency of ammonium sulphate in the control of Prymnesium parvum in fish breeding ponds. Appli. Microbiol. 1953, 1, 330–333. [Google Scholar]
- Body, A. Stable pond blooms through phosphate and pH control. Infofish Int. 2011, 1, 22–25. [Google Scholar]
- Johnsen, T.M.; Eikrem, W.; Olseng, C.D.; Tollefsen, K.E.; Bjerknes, V. Prymnesium parvum: The norwegian experience. J. Am. Water Resour. Assoc. 2010, 46, 6–13. [Google Scholar] [CrossRef]
- Roelke, D.L.; Grover, J.P.; Brooks, B.W.; Glass, J.; Buzan, D.; Southard, G.M.; Fries, L.; Gable, G.M.; Schwierzke-Wade, L.; Byrd, M.; et al. A decade of fish-killing Prymnesium parvum blooms in Texas: Roles of inflow and salinity. J. Plankton Res. 2011, 33, 243–253. [Google Scholar] [CrossRef]
- Tomas, C.; Glass, J.; Ralph, J.; Lewitus, A. Blooms of the ichthyotoxic flagellate Prymnesium parvum in US waters: An emerging or a perennial problem? Harmful Algae 2002, 1, 369–370. [Google Scholar]
- Manning, S.R.; La Claire, J.W. Prymnesins: Toxic metabolites of the golden alga, Prymnesium parvum Carter (Haptophyta). Mar. Drugs 2010, 8, 678–704. [Google Scholar] [CrossRef] [PubMed]
- Henrikson, J.C.; Gharfeh, M.S.; Easton, A.C.; Easton, J.D.; Glenn, K.L.; Shadfan, M.; Mooberry, S.L.; Hambright, K.D.; Cichewicz, R.H. Reassessing the ichthyotoxin profile of cultured Prymnesium parvum (golden algae) and comparing it to samples collected from recent freshwater bloom and fish kill events in North America. Toxicon 2010, 55, 1396–1404. [Google Scholar] [CrossRef] [PubMed]
- Shilo, M.; Aschner, M. Factors governing the toxicity of cultures containing the phytoflagellate Prymnesium parvum Carter. J. Gen. Microbiol. 1953, 8, 333–343. [Google Scholar] [CrossRef] [PubMed]
- Ulitzur, S.; Shilo, M. A sensitive assay system for determination of the ichthyotoxicity of Prymnesium parvum. Microbiology 1964, 36, 161–169. [Google Scholar]
- Johansson, N.; Granéli, E. Influence of different nutrient conditions on cell density, chemical composition and toxicity of Prymnesium parvum (Haptophyta) in semi-continuous cultures. J. Exp. Mar. Biol. Ecol. 1999, 239, 243–258. [Google Scholar] [CrossRef]
- Seger, A.; Body, A.; Hallegraeff, G.M. Exploration of Phoslock™ clay in mitigating Prymnesium parvum fish-killing algal blooms in aquaculture ponds. In Proceedings of the 15th International Conference on Harmful Algae, Changwon, South Korea, October 29–November 2 2015; Kim, H.G., Reguera, B., Hallegraeff, G., Lee, C.K., Han, M.S., Choi, J.K., Eds.; International Society for the Study of Harmful Algae: Busan, South Korea, 2014; pp. 231–234. [Google Scholar]
- Hagström, J.A.; Granéli, E. Removal of Prymnesium parvum (Haptophyceae) cells under different nutrient conditions by clay. Harmful Algae 2005, 4, 249–260. [Google Scholar] [CrossRef]
- Kurten, G.L.; Barkoh, A.; Begley, D.C.; Fries, L.T. Refining nitrogen and phosphorus fertilization strategies for controlling the toxigenic alga Prymnesium parvum. J. Am. Water Resour. Assoc. 2010, 46, 170–186. [Google Scholar] [CrossRef]
- Kurten, G.L.; Barkoh, A.; Fries, L.T.; Begley, D.C. Combined nitrogen and phosphorus fertilization for controlling the toxigenic alga Prymnesium parvum. N. Am. J. Aquac. 2007, 69, 214–222. [Google Scholar] [CrossRef]
- Ross, G.; Haghseresht, F.; Cloete, T.E. The effect of pH and anoxia on the performance of Phoslock™, a phosphorus binding clay. Harmful Algae 2008, 7, 545–550. [Google Scholar] [CrossRef]
- Sengco, M.R.; Hagström, J.A.; Granéli, E.; Anderson, D.M. Removal of Prymnesium parvum (Haptophyceae) and its toxins using clay minerals. Harmful Algae 2005, 4, 261–274. [Google Scholar] [CrossRef]
- Pierce, R.H.; Henry, M.S.; Higham, C.J.; Blum, P.; Sengco, M.R.; Anderson, D.M. Removal of harmful algal cells (Karenia brevis) and toxins from seawater culture by clay flocculation. Harmful Algae 2004, 3, 141–148. [Google Scholar] [CrossRef]
- Prochazka, E.; Hwang, G.S.; Shaw, G.; Hawker, D.W.; Stewart, I.; Wickramasinghe, W. The Removal of Microcystins in Drinking Water by Clay Minerals, Proceedings of 14th International Conference on Harmful Algae (ICHA), Crete, Greece, 1–5 November 2013; pp. 230–232.
- Lee, Y.-C.; Jin, E.; Jung, S.W.; Kim, Y.-M.; Chang, K.S.; Yang, J.-W.; Kim, S.-W.; Kim, Y.-O.; Shin, H.-J. Utilizing the algicidal activity of aminoclay as a practical treatment for toxic red tides. Sci. Rep. 2013, 3, 1–8. [Google Scholar]
- Sengco, M.R. The Aggregation of Clay Minerals and Marine Microalgal Cells: Physicochemical Theory and Implications for Controlling Harmful Algal Blooms; MIT/WHOI: Woods Hole, MA, USA, 2001. [Google Scholar]
- Rivera, P.P.L.; Azanza, R.V.; Diego-McGlone, M.L.S.; Sellner, K. Notable physiological and morphological effects of ball clay addition on bloom forming organisms Pyrodinium bahamense, Gymnodinium catenatum and Alexandrium catenella. In Proceedings of 16th International Conference on Harmful Algae, Wellington, New Zealand, 27–31 October 2014.
- Sengco, M.R.; Li, A.; Tugend, K.; Kulis, D.; Anderson, D.M. Removal of red- and brown-tide cells using clay flocculation. I. Laboratory culture experiments with Gymnodinium breve and Aureococcus anophagefferens. Mar. Ecol. Prog. Ser. 2001, 210, 41–53. [Google Scholar] [CrossRef]
- Dorantes-Aranda, J.J.; Waite, T.D.; Godrant, A.; Rose, A.L.; Tovar, C.D.; Woods, G.M.; Hallegraeff, G.M. Novel application of a fish gill cell line assay to assess ichthyotoxicity of harmful marine microalgae. Harmful Algae 2011, 366–373. [Google Scholar] [CrossRef]
- Deeds, J.R.; Terlizzi, D.E.; Adolf, J.E.; Stoecker, D.K.; Place, A.R. Toxic activity from cultures of Karlodinium micrum (=Gyrodinium galatheanum) (Dinophyceae)—A dinoflagellate associated with fish mortalities in an estuarine aquaculture facility. Harmful Algae 2002, 1, 169–189. [Google Scholar] [CrossRef]
- Figueroa, R.A.; Leonard, A.; Mackay, A.A. Modeling Tetracycline Antibiotic Sorption to Clays. Environ. Sci. Technol. 2004, 38, 476–483. [Google Scholar] [CrossRef]
- Polati, S.; Angioi, S.; Gianotti, V.; Gosetti, F.; Gennaro, M. Sorption of pesticides on kaolinite and montmorillonite as a function of hydrophilicity. J. Environ. Sci. Health 2006, 41, 333–344. [Google Scholar] [CrossRef]
- Fukahori, S.; Fujiwara, T.; Ito, R.; Funamizu, N. pH-Dependent adsorption of sulfa drugs on high silica zeolite: Modeling and kinetic study. Desalination 2011, 275, 237–242. [Google Scholar] [CrossRef]
- Kester, D.R.; Duedall, I.W.; Connors, D.N.; Pytkowicz, R.M. Preparation of artificial seawater. Limnol. Oceanogr. 1967, 12, 176–179. [Google Scholar] [CrossRef]
- Schirmer, K.; Chan, A.G.J.; Greenberg, B.M.; Dixon, D.G.; Bols, N.C. Methodology for demonstrating and measuring the photocytotoxicity of fluoranthene to fish cells in culture. Toxicol. in Vitro 1997, 11, 107–119. [Google Scholar] [CrossRef] [PubMed]
- Hunter, R.J. Zeta Potential in Colloid Science, Principles and Applications; Academic Press: New York, NY, USA, 1981; p. 31. [Google Scholar]
- Baker, J.W.; Grover, J.P.; Brooks, B.W.; Ureña-Boeck, F.; Roelke, D.L.; Errera, R.; Kiesling, R.L. Growth and toxicity of Prymnesium parvum (Haptophyta) as a function of salinity, light, and temperature. J. Phycol. 2007, 43, 219–227. [Google Scholar] [CrossRef]
- Yu, Z.; Sengco, M.R.; Anderson, D.M. Flocculation and removal of the brown tide organism, Aureococcus anophagefferens (Chrysophyceae), using clays. J. Appli. Phycol. 2004, 16, 101–110. [Google Scholar] [CrossRef]
- Stückenschneider, K.; Merz, J.; Hanke, F.; Rozyczko, P.; Milman, V.; Schembecker, G. Amino-acid adsorption in MFI-type zeolites enabled by the pH-dependent ability to displace water. J. Phys. Chem. C 2013, 117, 18927–18935. [Google Scholar] [CrossRef]
- Chorom, M.; Rengasamy, P. Dispersion and zeta potential of pure clays as related to net particle charge under varying pH, electrolyte concentration and cation type. Eur. J. Soil Sci. 1995, 46, 657–665. [Google Scholar] [CrossRef]
- Saka, E.; Güler, C. The effects of electrolyte concentration, ion species and pH on the zeta potential and electrokinetic charge density of montmorillonite. Clay Miner. 2006, 41, 853–861. [Google Scholar] [CrossRef]
- Moayedi, H.; Huat, B.B.; Kazemian, S.; Daneshmand, S.; Moazami, D.; Niroumand, H. Electrophoresis of suspended kaolinite in multivalent electrolyte solution. Int. J. Electrochem. Sci. 2011, 6, 6514–6524. [Google Scholar]
- Bilgiç, C. Investigation of the factors affecting organic cation adsorption on some silicate minerals. J. Colloid Interface Sci. 2005, 281, 33–38. [Google Scholar] [CrossRef] [PubMed]
- Niriella, D.; Carnahan, R. Comparison study of zeta potential values of bentonite in salt solutions. J. Dispers. Sci. Technol. 2006, 27, 123–131. [Google Scholar] [CrossRef]
- Yukselen, Y.; Kaya, A. Zeta potential of kaolinite in the presence of alkali, alkaline earth and hydrolyzable metal ions. Water Air Soil Pollut. 2003, 145, 155–168. [Google Scholar] [CrossRef]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license ( http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Seger, A.; Dorantes-Aranda, J.J.; Müller, M.N.; Body, A.; Peristyy, A.; Place, A.R.; Park, T.G.; Hallegraeff, G. Mitigating Fish-Killing Prymnesium parvum Algal Blooms in Aquaculture Ponds with Clay: The Importance of pH and Clay Type. J. Mar. Sci. Eng. 2015, 3, 154-174. https://doi.org/10.3390/jmse3020154
Seger A, Dorantes-Aranda JJ, Müller MN, Body A, Peristyy A, Place AR, Park TG, Hallegraeff G. Mitigating Fish-Killing Prymnesium parvum Algal Blooms in Aquaculture Ponds with Clay: The Importance of pH and Clay Type. Journal of Marine Science and Engineering. 2015; 3(2):154-174. https://doi.org/10.3390/jmse3020154
Chicago/Turabian StyleSeger, Andreas, Juan José Dorantes-Aranda, Marius N. Müller, Adam Body, Anton Peristyy, Allen R. Place, Tae Gyu Park, and Gustaaf Hallegraeff. 2015. "Mitigating Fish-Killing Prymnesium parvum Algal Blooms in Aquaculture Ponds with Clay: The Importance of pH and Clay Type" Journal of Marine Science and Engineering 3, no. 2: 154-174. https://doi.org/10.3390/jmse3020154