1. Introduction
Symbiotic dinoflagellates of the genus
Symbiodinium provide their coral hosts with photosynthetic products that sustain polyp metabolism and growth in oligotrophic waters [
1,
2]. Environmental stresses, such as elevated seawater temperatures and high irradiances, may disrupt the symbiosis, leading to losses of either
Symbiodinium cells or the pigmentation of those within the host tissues; this is the phenomenon of coral bleaching [
3,
4]. The mechanism of coral bleaching is not fully understood. However, it is clear that damage to the algal photosynthetic apparatus may lead to the production of reactive oxygen species (ROS) that cause apoptosis, autophagy, and necrosis of host and algal cells [
4,
5,
6]. Details of the pathway to cell death depend on the intensity and duration of the environmental stress [
4].
Thermal stress has different impacts on the photobiology and growth of diverse phylotypes of cultured
Symbiodinium cells [
7,
8,
9,
10,
11]. The production of reactive oxygen species (ROS) in response to elevated temperature differs among
Symbiodinium types [
12,
13]. Constitutive activity and up-regulation of components of the antioxidant network under thermal stress are different among
Symbiodinium types [
14]. Thus, variations in the physiological characteristics of
Symbiodinium types may be crucial in determining the bleaching susceptibility of coral-algal symbiotic complex, though properties of the host may also influence the bleaching susceptibility of the symbiotic complex [
15,
16].
Impairment of the
Symbiodinium photosynthesis mechanism is most likely the main factor that initiates the coral bleaching. Several hypotheses have been proposed to explain how elevated temperatures affect the photochemical pathway: excessive photon absorption by light harvesting antennae damages the PSII reaction center, particularly the D1 protein [
17,
18,
19]; elevated temperatures may limit photosynthesis by destabilizing the thylakoid membranes [
7]; the activity of Rubisco may be a primary site of damage by elevated temperature [
20], though a recent study suggests that inhibition of the Calvin-Benson cycle alone does not induce coral bleaching [
21]; damaged PSII reaction centers may be replaced by re-synthesized D1 protein, but this mechanism is inhibited when temperature is elevated [
10]; elevated temperatures may also suppress the synthesis of light harvesting antennae protein at the translational step, thereby promoting losses of major light harvesting proteins [
9]. Several of these postulates have been supported by recent studies indicating that the thermal stress susceptibility of
Symbiodinium is determined by the repair capacity of the photosynthetic mechanism [
10,
22,
23].
Robison and Warner [
8] found that growth of A1 phylotype algal symbionts declined significantly with a slight reduction in PSII activity under combined high temperature and high light stress. Growth of B1 algae declined similarly when exposed to high light alone, but PSII efficiency was much reduced. Krämer
et al. [
24] observed similar differences in growth and PSII responses to combined thermal and light stresses between the clade A subtypes A1 and Ax. They suggested that photosynthesis and growth become uncoupled in A1 cells and that the decline in growth might have resulted from a high energy demand for protein repair and increased respiratory losses. Thus, environmental stress may act on different target processes among the types of
Symbiodinium. Growth inhibition may well occur in some of the
Symbiodinium types without significant impairment of their PSII efficiencies. Improved understanding requires investigation of (i) photosynthesis and growth responses of diverse
Symbiodinium types under stress, and (ii) the PSII repair capacity and other metabolic responses of
Symbiodinium cells exposed to stresses that cause bleaching.
In the current study, we examined the relationships between growth rate and photosynthetic activity (Fv/Fm) in six cultured Symbiodinium spp. strains (clade A, B, C, D, and F) exposed to elevated temperatures. We also investigated the recovery of photodamaged-PSII at high temperature in the presence or absence of a chloroplast protein synthesis inhibitor (lincomycin).
2. Materials and Methods
2.1. Cultures and Growth Conditions
Six cultures of Symbiodinium spp., belonging to five clades, were used: KB8 (clade A, derived from the jellyfish Cassiopea sp.), Y106 (clade A, from the bivalve Tridacna crocea), K100 (clade B, from the sea anemone Aiptasia pulchella), Y103 (clade C, from the bivalve Fragum sp.), K111 (clade D, from the soft coral Sarcophyton glaucum) and K102 (clade F, originally from the scleractinian coral Montipora verrucosa). The Symbiodinium clades were identified based on 28S rDNA RFLP analysis using the restriction enzyme TaqI. The six cultures will be referred to as A-KB8, A-Y106, B-K100, C-Y103, D-K111, and F-K102 (clade type followed by culture name). Some strains were identified at subclade level based on nucleotide sequence of ITS2 region. These stock cultures had been maintained in f/2 medium (Sigma-Aldrich, St Louis, MO, USA) at 25 °C under a photon irradiance of 5–9 μmol photons m−2∙s−1(12 h light:12 h dark photoperiod cycle) provided by a white fluorescent lamp (Toshiba FL2022-BRN/18, Fuzhou, China). Cultures were transplanted to a new medium and put into another incubator (LH-70CCFL-CT, NK System, Tokyo, Japan) at the control condition (25 °C, 22–49 μmol photons m−2∙s−1) for two weeks before the stress exposure experiment started.
2.2. Standard Curves of Symbiodinium Cell Densities and in Vivo Chlorophyll Fluorescence
The growth rate of Symbiodinium cells at different temperatures was measured by changes in in vivo fluorescence. We first established a standard curve for the relationship between Symbiodinium cell density and chlorophyll fluorescence intensity for each strain one day before the stress exposure experiment. The cell densities of stock cultures were measured using a Neubauer improved hemocytometer (Hirschmann, Eberstadt, Germany) under a light microscope (Nikon, Optiphot-2, Tokyo, Japan). A suspension of Symbiodinium cells at known density (2 × 106 cells mL−1) was made up using the hemocytometer, and a series of dilutions (between 1 × 104 and 1 × 106 cells mL−1) were prepared. We then measured the chlorophyll a fluorescence intensities of tubes containing various concentrations of Symbiodinium cells using a fluorometer (TD700, Turner Design, Sunnyvale, CA, USA) equipped with excitation (436 nm) and emission (680 nm) filters, and a daylight white lamp. The fluorescence intensities were plotted against actual densities of Symbiodinium cells to obtain standard relationships between fluorescence intensity and Symbiodinium cell density. The standard curves were linear in the following range: 1 × 104 to 1 × 106 cells mL−1, which was used in the following experiment.
2.3. Growth Rate of Symbiodinium Cells at Different Temperatures
To compare the physiological responses of the six strains of Symbiodinium, we measured the growth rates of cultured cells of each strain at temperatures of 25 °C, 30 °C, and 33 °C and the photochemical efficiency (Fv/Fm) of the algal cells after 14 days of temperature treatment. For each strain, we prepared 12 borosilicate glass tubes (10 mL, Fisherbrand, Pittsburgh, PA, USA) filled with 6 mL of f/2 medium containing Symbiodinium cells at an initial density of 5 × 104 cells mL−1. The 12 tubes were separated into three temperature-treatment groups, which were incubated at 25 °C, 30 °C or 33 °C in separate incubators (LH-70CCFL-CT, NK System, Tokyo, Japan). The cultures were illuminated by fluorescent lamps (NK System LH-55/70CCFL-CT), which provided a photon irradiance of 22–49 μmol photons m−2∙s−1 (12 h light:12 h dark photoperiod). Each temperature treatment was replicated four-fold. The control tubes contained cell-free filtered seawater subjected to the same treatments. To avoid the effect of different light levels on the growth rate, we moved the position of tubes randomly, everyday during the experimental period.
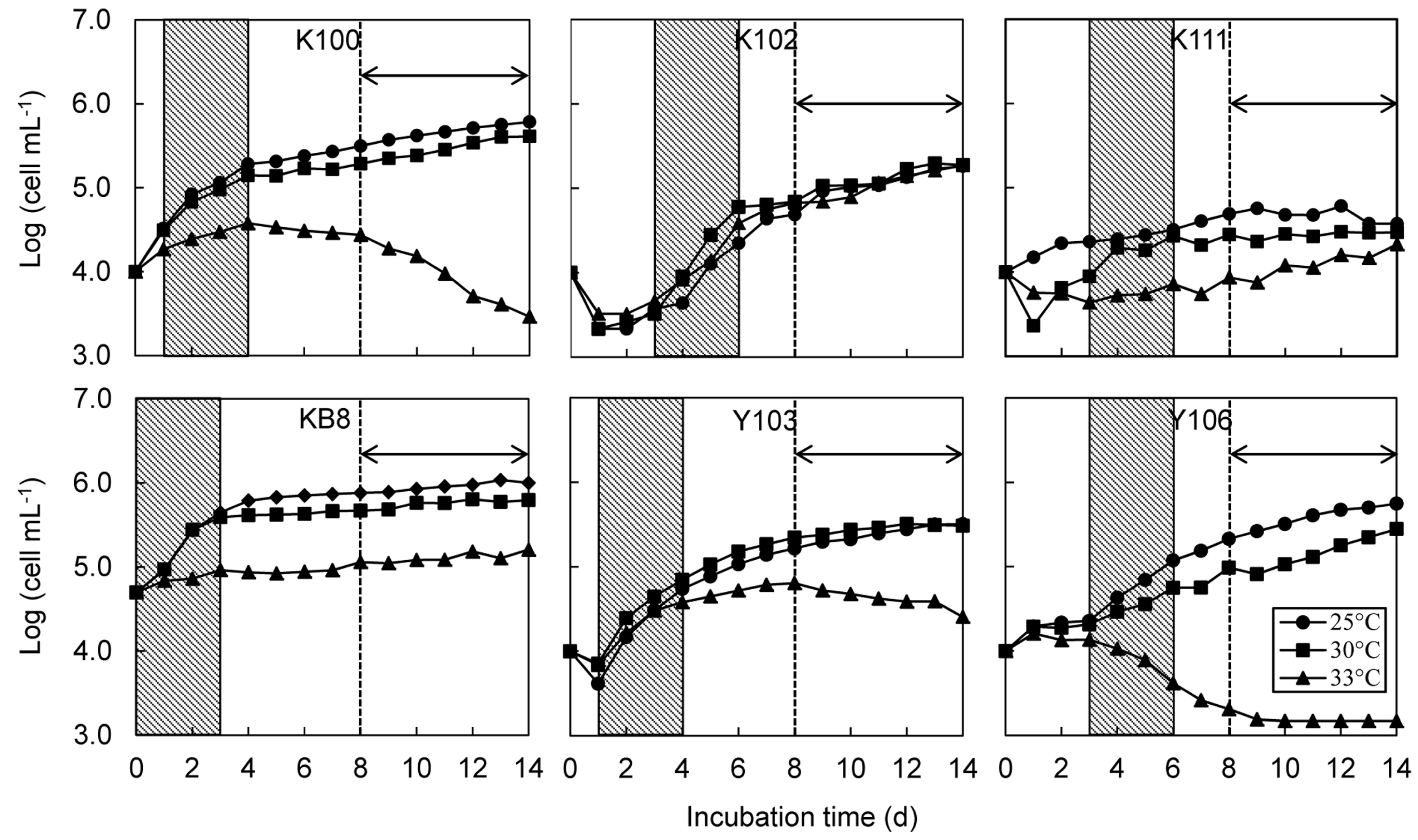
Figure 1.
Growth curves of six strains of Symbiodinium at different temperatures. Changes in mean cell density (n = 4 for each strain) are plotted over a 14-day period. Shaded areas indicates initial logarithmic phases, which were used for calculations of initial growth rate (μ). The arrow and dotted line indicate the growth period used for calculation of secondary growth rate (μs).
Figure 1.
Growth curves of six strains of Symbiodinium at different temperatures. Changes in mean cell density (n = 4 for each strain) are plotted over a 14-day period. Shaded areas indicates initial logarithmic phases, which were used for calculations of initial growth rate (μ). The arrow and dotted line indicate the growth period used for calculation of secondary growth rate (μs).
The tubes containing
Symbiodinium were mixed once daily using a vortex to keep the cells in suspension. We measured fluorescence immediately after vortexing each day for 14 days during the late afternoon (16.00–18.00) using a TD700 fluorometer, thereby circumventing potential effects of a diurnal cell cycle. Cell density was calculated from fluorescence intensity by using the standard curve for each strain. The log (cell density) values were plotted against time (days after inoculation) and the growth rate was estimated from the slope of the regression fit in the linear portion of the semi-logarithmic growth curve [
25]. The linear portion of the growth curve was selected visually [
25]. Log phase growth was linear for 3–5 days; thereafter, growth slowed or plateaued during the stationary phase. High temperature-treated suspensions often had biphasic growth curves; cultures grew linearly at first, but growth became negative after day 8 of the treatment. Thus, we measured an initial growth rate (μ) based on the initial linear portion of the growth curve (days 1–4 for B-K100 and C-Y103, days 3–6 for F-K102, D-K111 and A-Y106 and days 0–3 for A-KB8), and a secondary growth rate (μ
s) based on the later linear portion for days 8 to 14 (
Figure 1). Cell density often decreased on the day after inoculation into the experimental tubes. Therefore, cell density data on day 0 were not used for strains other than A-KB8. The initial density drop may have occurred because cells adhered to the bottoms of the tubes after initial inoculation.
2.4. Measurement of the Photochemical Efficiency of PSII
The photochemical efficiency of PSII (
Fv/Fm) was determined at the end of the growth inhibition experiment (day 14). After vortexing, 3 mL of
Symbiodinium cell suspension were removed from each tube using a syringe. The cells were captured on a Millipore filter (13-mm diameter, 0.45-μm pore size; Millipore, Cork, Ireland). Each filter bearing captured cells was sandwiched between two glass slides held in place with transparent adhesive tape. Cells mounted in this manner were immersed in filtered seawater and dark-adapted for 20 min in a custom-made acrylic black box at room temperature. We measured
Fv/Fm using a Mini-PAM (pulse-amplitude modulated) fluorometer (Walz, Effeltrich, Germany) (measuring light 8, saturation pulse intensity 8, width 0.8; gain = 2) [
26].
Since cell densities in strains of B-K100 and A-Y106 kept at 33 °C were too low for measurements of Fv/Fm, we performed an additional experiments to monitor changes in the Fv/Fm values of strains B-K100 and A-Y106, and C-Y103. We measured cell densities and Fv/Fm values of these strains at 33 °C every second day for 12 days. In the additional experiment, we were able to measure Fv/Fm values without a low signal warning on the Mini-PAM apparatus.
2.5. Photoinhibition Experiments
We compared the sensitivity of PSII to light stress and recovery from photodamage among the six Symbiodinium strains at 25 °C and 33 °C. Cultures were incubated with or without lincomycin, an inhibitor of thylakoid membrane protein synthesis (Wako, Osaka, Japan), to determine the extent to which photodamage and its recovery process depend on re-synthesis of thylakoid membrane proteins, including the D1 protein.
Sixteen 25-mL Falcon cell culture flasks filled with 15-mL f/2 medium containing Symbiodinium cells at a density of 1 × 106 mL−1 were prepared for each strain. The 16 culture flasks were divided to four groups and exposed to high light (470 μmol photons m−2∙s−1) at 25 °C or 33 °C in the presence or absence of 200 μg mL−1 lincomycin. Each treatment was replicated fourfold. Light was provided by two metal halide lamps (“Funnel 2”, Kamihata, Tokyo, Japan), and the temperature was regulated by using waterbaths (WTB, Taitec, Saitama-ken, Japan).
We added 100-μL lincomycin to each culture flask in the lincomycin-treatment group (in darkness) 20 min before exposure to light, thereby making the final concentration 200 μg mL
−1 lincomycin. Cultures with or without the inhibitor were subsequently exposed to strong light at 25 °C or 33 °C for 2 h. After the light exposure treatment,
Symbiodinium cells were transferred to darkness for 1 h at each temperature, and then allowed to recover under a photon irradiance of 20 μmol photons m
−2∙s
−1 (provided by a ColdSpot, PCS-NHF150, Japan) for 2 h at each temperature. A new lincomycin dose was added 20 min before the recovery period since the antibiotic may have degraded during the 2-h light exposure treatment [
27].
Fv/Fm values were measured in 1.5-mL samples of
Symbiodinium cell suspensions taken from the flasks before and after strong light treatment, and following the recovery period.
2.6. Statistical Analysis
Mean growth rates and mean Fv/Fm values of Symbiodinium cells were compared among treatments using Analysis of Variance (ANOVA) and Tukey’s honestly significant difference (HSD) test when the data met requirements of inferential parametric tests. Normality and homogeneity of variances were tested using Kolmogorov-Smirnov and Lilliefors and Levene’s tests, respectively. The data were square root and arcsine transformed when necessary. When transformed data did not meet requirements of parametric testing, we used the non-parametric Kruskal-Wallis ANOVA followed by multiple comparisons tests. All statistical analyses were performed using Statistica 6.0 software (StatSoft Inc., Tulsa, OK, USA).
4. Discussion
4.1. Growth rate of Symbiodinium Cells at Different Temperatures
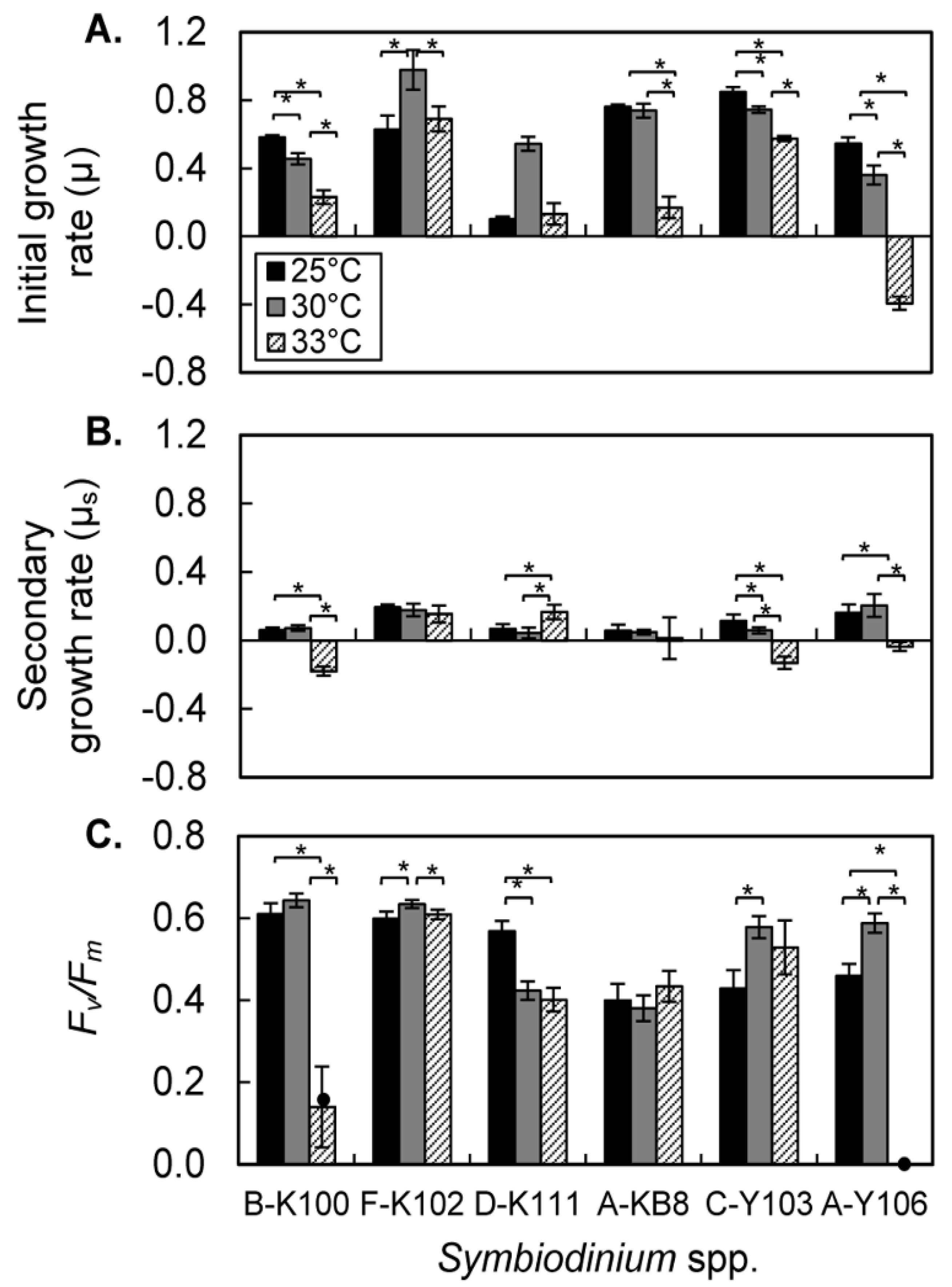
The growth rates of all six strains in the logarithmic phase were in the range 0.6–0.8 at 25 °C. High temperature had different effects among strains. Growth rates were significantly depressed in B-K100, A-KB8, C-Y103, and A-Y106 at 33 °C. A-Y106 had a negative growth rate at 33 °C. Only F-K102 had similar growth rates at 25 °C and 33 °C.
The secondary, or stationary, growth rates were lower than rates during the initial logarithmic phase. High temperature significantly reduced the secondary growth rate of B-K100, C-Y103, and A-Y106; all of their secondary growth rates were negative at 33 °C. A-KB8 had a reduced secondary growth rate at 33 °C, but the effect was not significant due to large variance in the data. Only F-K102 had similar secondary growth rates at 25 °C, 30 °C, and 33 °C. Thus, B-K100, C-Y103, and A-Y106 were highly sensitive to high temperatures, and unable to proliferate at 33 °C. A population collapse would be expected at this temperature. A-KB8 was also sensitive to high temperature, but nevertheless able to maintain cell densities at 33 °C. F-K102 was highly tolerant of elevated temperature and continued growing at 33 °C. The effect of high temperature on the growth rate of D-K111 differed among growth phases; however artifacts caused by cell adherence to culture tube walls introduced a considerable degree of uncertainty in ascertaining temperature responses of this strain. In our preliminary study, we counted the number of adhered cells and those in suspension after 21 days exposure to different temperatures. We found that the percentage of adhered cells was highest in D-K111. Furthermore, high temperature increased the percentage of adhered cells in high temperature-sensitive strains (B-K100 and A-Y106). Improved method should be applied to estimate growth response of strains with tendency to adhere to culture tube walls [
24]
The four-day duration of the logarithmic phase in this study was relatively short. This might be partly due to a relatively high initial cell density used in this study. An initial reduction in cell densities after cell inoculation into the culture vessels occurred in all strains other than A-KB8. This reduction may be explained in part by the adhesion of some cells to the culture tube walls and in part by the initial lag phase before cells entered the logarithmic phase, as reported previously for green algae [
28].
4.2. Photochemical Efficiency of PSII after High Temperature Treatment
Photochemical efficiencies (Fv/Fm) of B-K100 and A-Y106 were significantly depressed at high temperature (33 °C) in comparison with efficiencies at 25 °C. Fv/Fm values of D-K111 were also significantly reduced at 33 °C, although the ratio (ca. 0.4) was, nevertheless, relatively high. Fv/Fm values of F-K102, A-KB8, and C-Y103 were similar at 25 °C and 33 °C.
Table 1 shows the effects of temperature on the initial growth rates, secondary growth rates, and
Fv/Fm ratios for each of the strains. A-Y106 was the most sensitive to high temperature stress: both initial and secondary growth rates were negative, and
Fv/Fm decreased to zero after 10-days of exposure to 33 °C. B-K100 was the next most sensitive to thermal stress; it had a negative secondary growth rate and a marked reduction in
Fv/Fm at 33 °C. A-KB8 and C-Y103 were notably different from other strains; their secondary growth rates became negative even though
Fv/Fm values remained high. Strain F-K102 was the most tolerant of thermal stress; it had the highest
Fv/Fm ratio and initial growth rate at 30 °C. The
Fv/Fm values and secondary growth rates of this strain were closely similar at 33 °C and 25 °C. D-K111 was tolerant of thermal stress: it retained a high
Fv/Fm values at 33 °C and it secondary growth rate at this temperature was significantly higher than those at 25 °C or 30 °C. However, the tendency of D-K111 cells to adhere to the wall of the tubes confounded measurements of cell density by the
in vivo fluorescence method used in this study. Thus, the six strains of
Symbiodinium fell into three categories: (1) growth responses and photochemical efficiencies were highly sensitive to high temperature (B-K100 and A-Y106), (2) the growth responses were highly sensitive to high temperature but photochemical efficiencies were not (C-Y103 and A-KB8), (3) growth responses and photochemical efficiencies were both highly tolerant of elevated temperature (F-K102 and D-K111).
Table 1.
Effects of temperature on the initial growth rate, secondary growth rate, and Fv/Fm of the Symbiodinium strains. Conditions under which significant recovery from photodamage was observed are also shown.
Table 1.
Effects of temperature on the initial growth rate, secondary growth rate, and Fv/Fm of the Symbiodinium strains. Conditions under which significant recovery from photodamage was observed are also shown.
| Symbiodinium strain | Initial growth rate (μ) | Secondary growth rate (μs) | Fv/Fm | Recovery | Category |
|---|
| B-K100 (clade B) | 25 °C > 30 °C > 33 °C | 25 °C = 30 °C > 33 °C | 25 °C = 30 °C > 33 °C | * (25 °C) | heat-sensitive |
| A-Y106 (clade A) | 25 °C > 30 °C > 33 °C | 25 °C = 30 °C > 33 °C | 25 °C < 30 °C > 33 °C | * (25 °C), 25 °C (+Lin)) |
| A-KB8 (clade A) | 25 °C = 30 °C > 33 °C | 25 °C > 30 °C > 33 °C | 25 °C = 30 °C = 33 °C | * (33 °C) | uncoupling of growth and Fv/Fm |
| C-Y103 (clade C) | 25 °C > 30 °C > 33 °C | 25 °C > 30 °C > 33 °C | 25 °C < 30 °C = 33 °C | * (25 °C) |
| F-K102 (clade F) | 25 °C < 30 °C > 33 °C | 25 °C = 30 °C = 33 °C | 25 °C < 30 °C > 33 °C | * (25 °C, 33 °C) | heat-tolerant |
| D-K111 (clade D) | 25 °C = 30 °C = 33 °C | 25 °C = 30 °C < 33 °C | 25 °C > 30 °C = 33 °C | * (33 °C) | heat-tolerant |
Category 2 is unique in that growth responses and PSII activities were uncoupled. Robison and Warner [
8] and Krämer
et al. [
24] also reported such uncoupling in
Symbiodinium strain A1. It is possible that these strains may allocate more energy to repairing damaged PSII mechanisms (resynthesizing protein D1 and other thylakoid proteins) and antioxidant defense mechanisms than to cell proliferation. The repair of damaged PSII is a costly process compared to other photo-protective mechanisms, such as thermal dissipation through NPQ. Such allocations to repair processes may therefore reduce growth rates. It is also possible that metabolic pathways other than the photosynthetic mechanism are affected by elevated temperatures in these strains such as fatty acid composition [
29] and reduction of motility [
30]. These possibilities await further investigation. We should also note that the photochemical efficiency of PSII can remain high even when there is a significant decline in PSII protein content under thermal stress [
31]. Although category 2
Symbiodinium cells in this study were derived from hosts other than corals, A1
Symbiodinium isolated from the coral
Stylophora pistillata also shows similar uncoupling [
24]. It is important to investigate how this uncoupling between algal growth and PSII activity affects susceptibility of corals to thermal stress.
4.3. Recovery of Photodamage of PSII
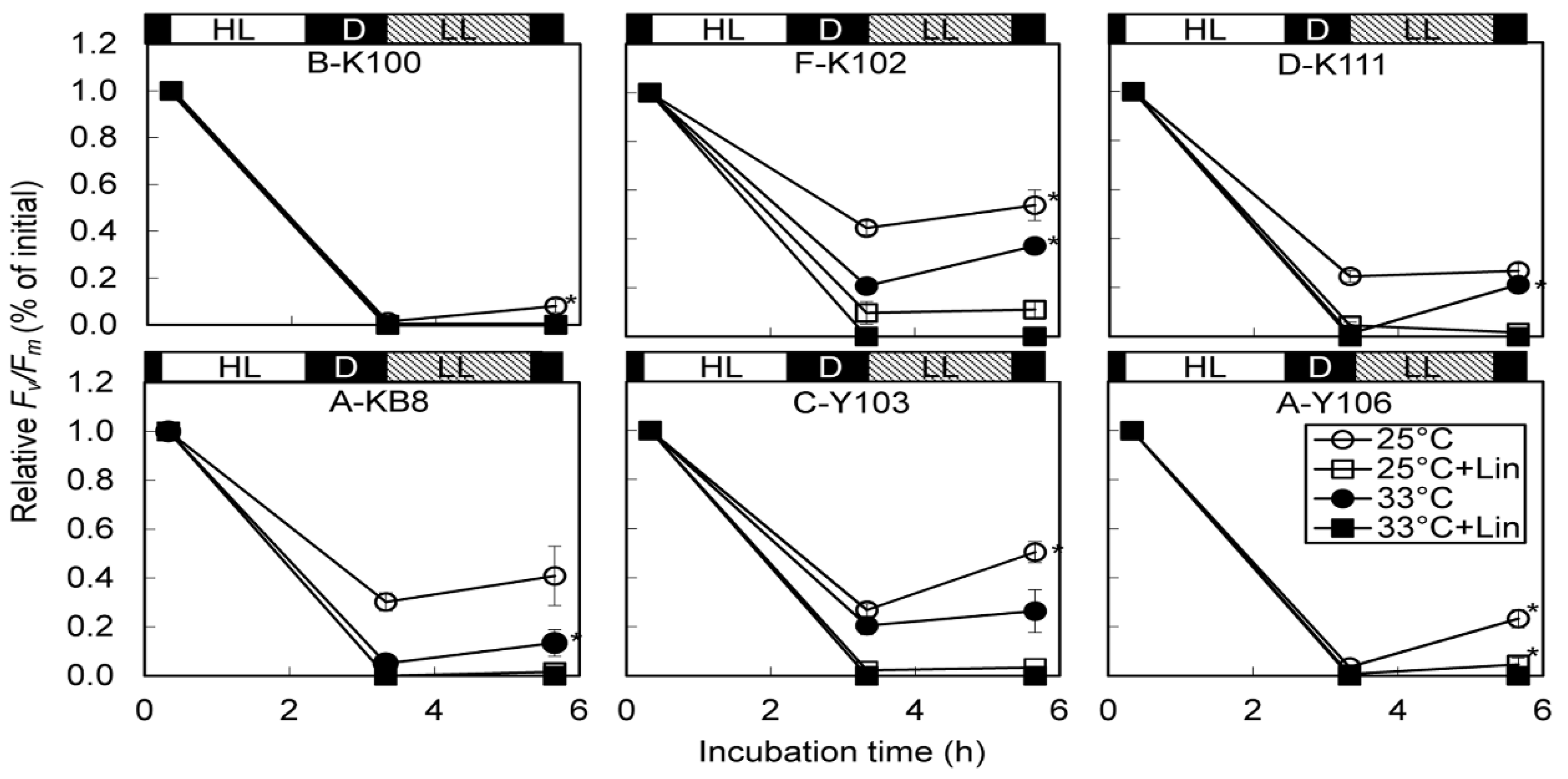
A significant decline of relative Fv/Fm was observed after 2-h exposure to high light. However, the extent of photodamage differed among Symbiodinium types. Declines at 33 °C were more marked than those at 25 °C, indicating that high temperature exacerbated photodamage. Lincomycin also exacerbated photodamage and reduced recovery of PSII function.
Fv/Fm values of B-K100 and A-Y106 decreased to almost zero after 2 h of high light treatment; no recovery occurred at 33 °C. The extremely low growth rates of B-K100 and A-Y106 at elevated temperatures might have been due to severe thermal damage to PSII. An inability to resynthesize D1 protein during stress treatment or recovery at 33 °C is the most probable explanation. F-K102 had relatively high Fv/Fm values after strong light treatment and significant lincomycin-sensitive recovery at both 25 °C and 33 °C. An elevated capacity for repairing thylakoid proteins is likely related to the high thermal tolerance of this strain. The high growth rate in F-K102 at elevated temperature may be indicative of an enhanced capacity to replace damaged-D1 protein during thermal and light stresses.
Growth rate and photochemical efficiency were uncoupled in A-KB8 and C-Y103, and their high photochemical efficiencies at elevated temperatures may be attributable to highly efficient re-synthesis of D1 and other thylakoid proteins at high temperature. If this were the case, these strains may reallocate energy from growth to the repair of damaged photosynthetic machineries or protection pathways, as suggested by Robison and Warner [
8] and Krämer
et al. [
24]. In contrast, recoveries of photodamaged PSII at 33 °C were modest in strains A-KB8 and C-Y103. Thus, thermal stressing of other metabolic pathways may have reduced growth rates in these two strains. This possibility should be explored in future research efforts.






{kind=link}
{kind=link}
{kind=link}