Transcriptome Assembly of the Bast Fiber Crop, Ramie, Boehmeria nivea (L.) Gaud. (Urticaceae)



Abstract
:1. Introduction
2. Materials and Methods
3. Results
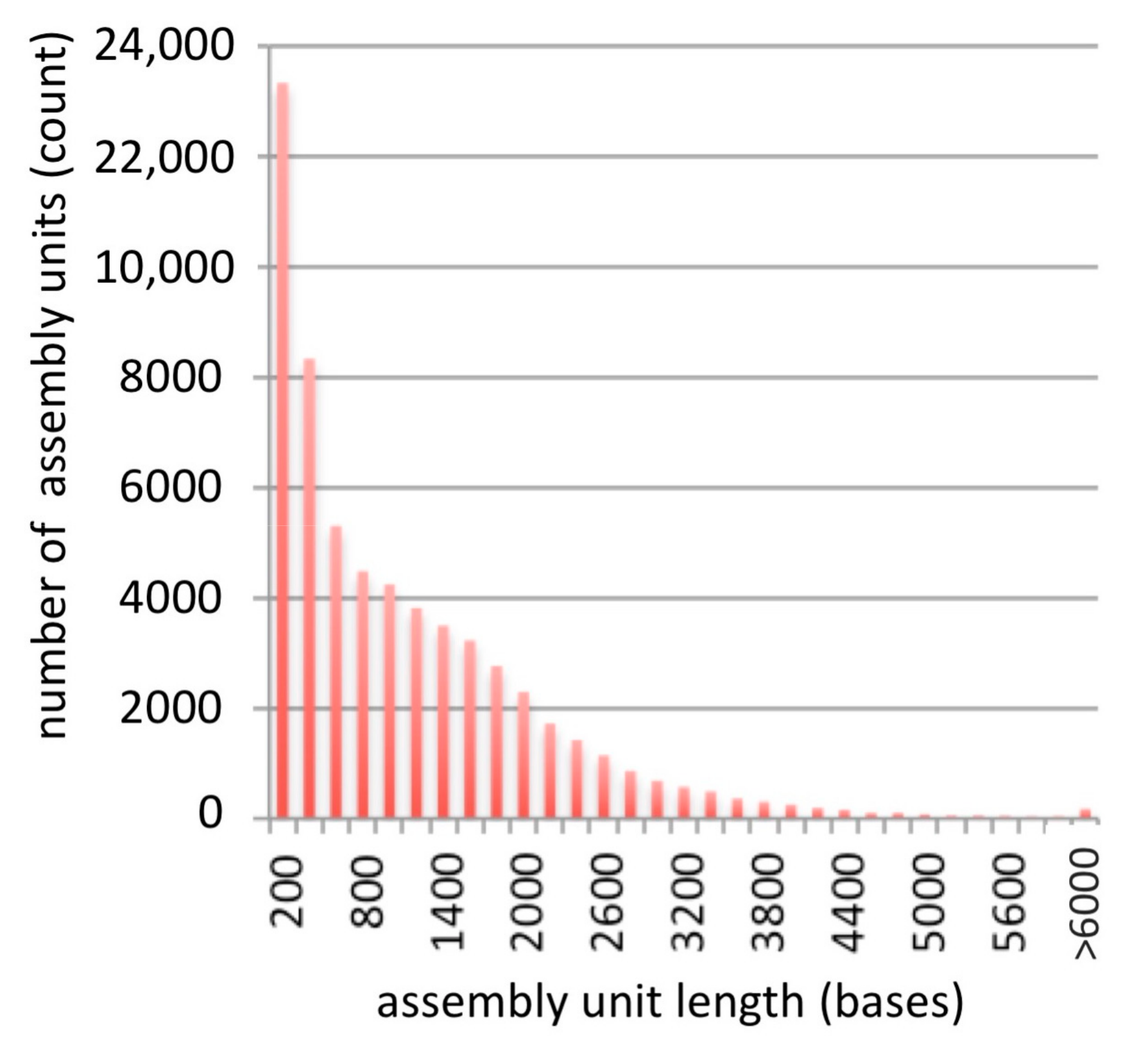
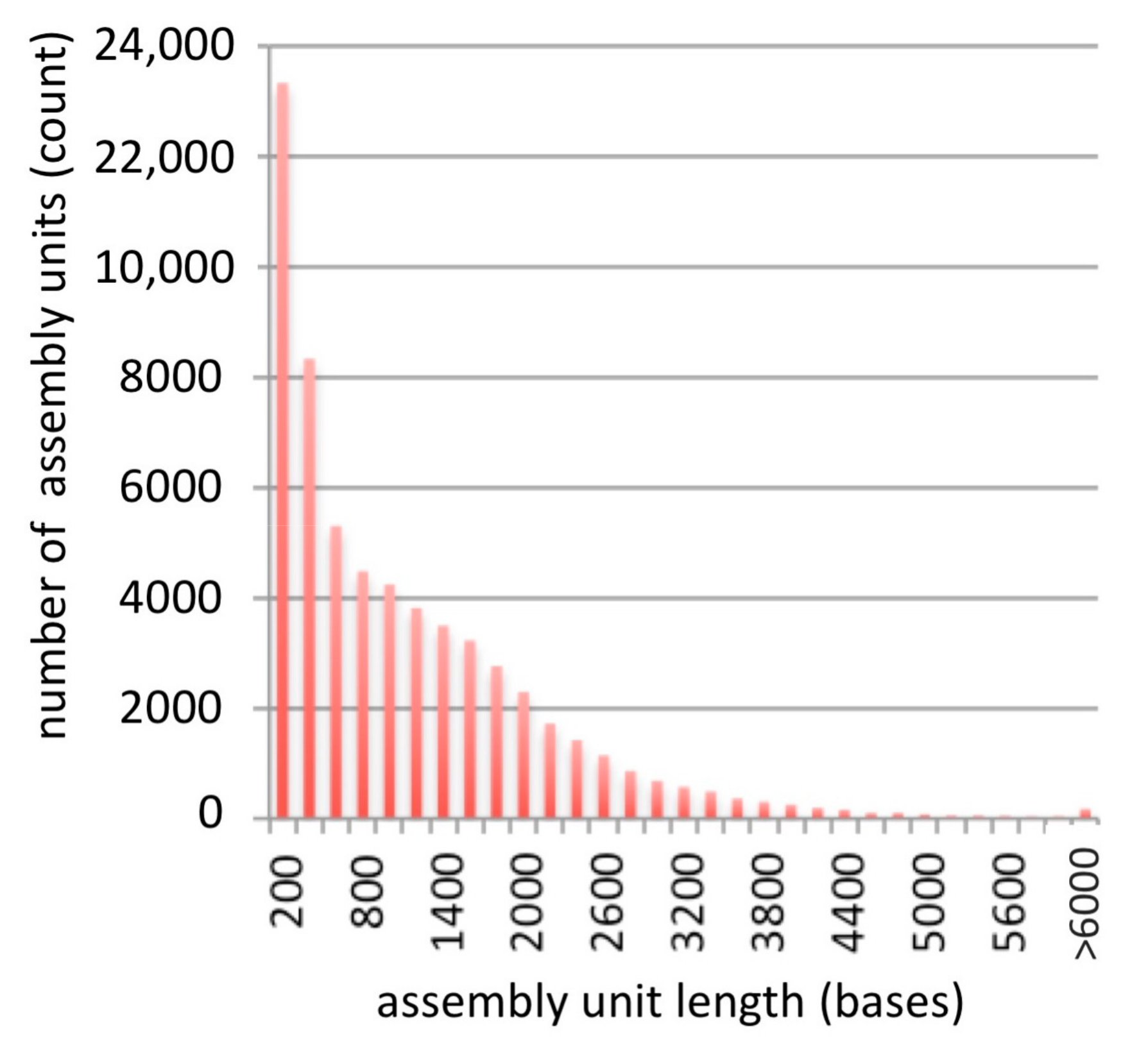
3.1. Assembly
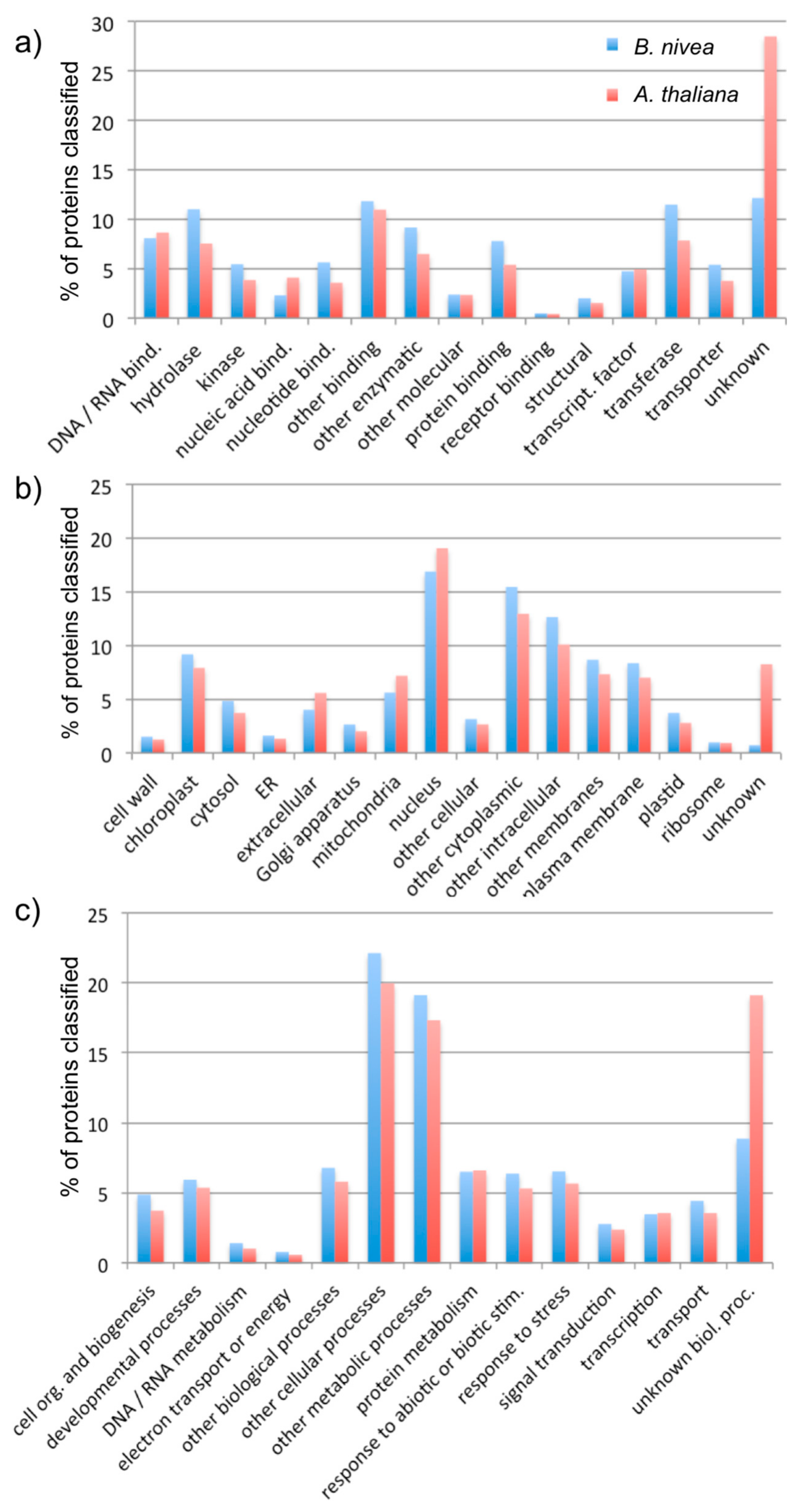
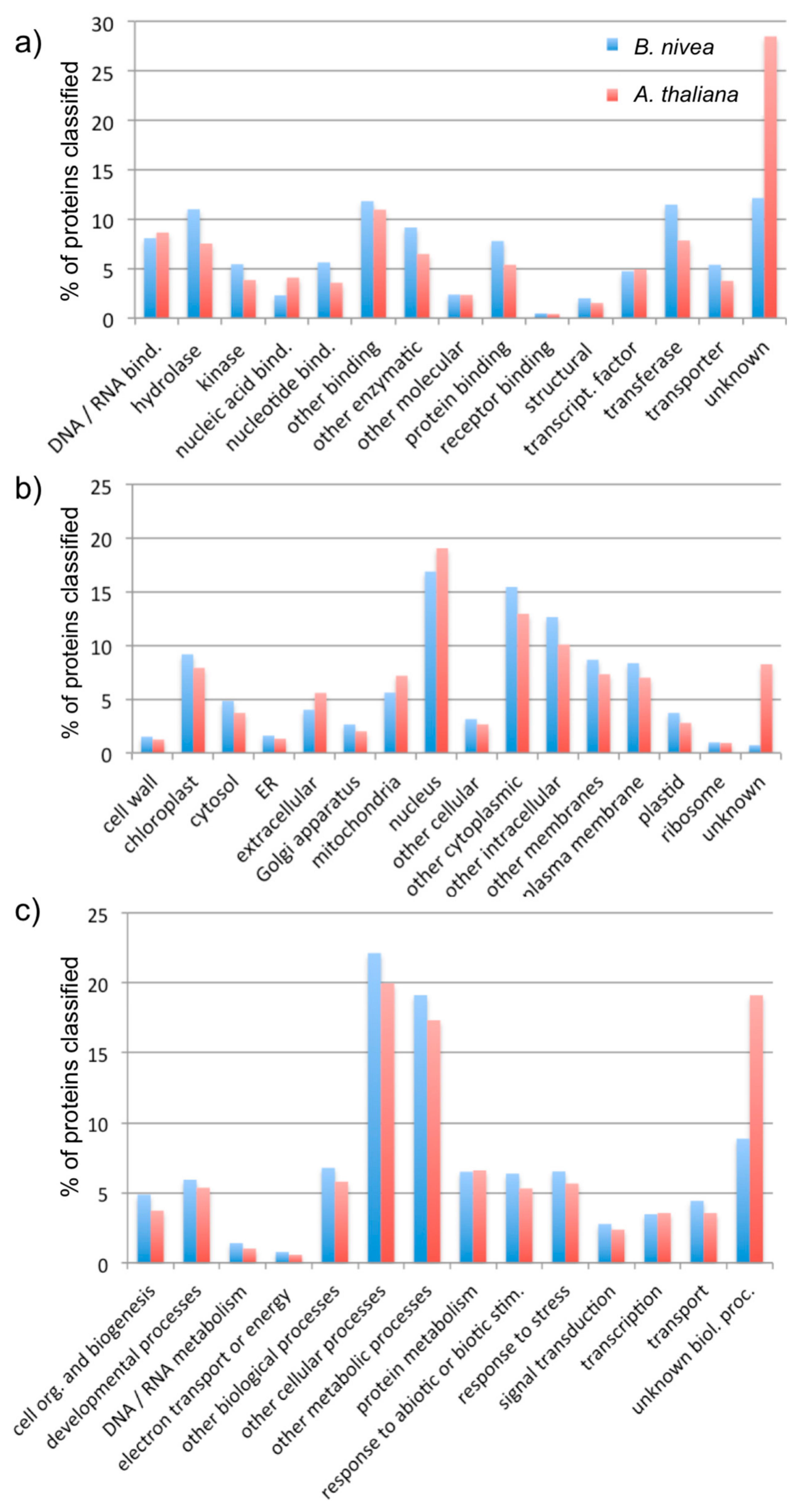
3.2. Analysis of Assembly Quality
4. Discussion
5. Conclusions
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Liao, L.; Li, T.J.; Zhang, J.; Xu, L.L.; Deng, H.S.; Han, X.J. The domestication and dispersal of the cultivated ramie (Boehmeria nivea (L.) Gaud. in Freyc.) determined by nuclear SSR marker analysis. Genet. Resour. Crop Evol. 2014, 61, 55–67. [Google Scholar] [CrossRef]
- Wang, H.; Xian, G.; Li, H.; Sui, L. Durability study of a ramie-fiber reinforced phenolic composite subjected to water immersion. Fibers Polym. 2014, 15, 1029–1034. [Google Scholar] [CrossRef]
- Li, J.; Feng, J.H.; Zhang, H.; Zhang, J.C. Wear properties of hemp, ramie and linen fabrics after liquid ammonia/crosslinking treatment. Fibres Text. East. Eur. 2010, 18, 81–85. [Google Scholar]
- Summerscales, J.; Gwinnett, C. Forensic Identification of Bast Fibres. In Biocomposites for High-Performance Applications; Ray, D., Ed.; Elsevier B.V.: Amsterdam, The Netherlands, 2017; pp. 125–164. [Google Scholar]
- Huang, K.-L.; Lai, Y.-K.; Lin, C.-C.; Chang, J.-M. Inhibition of hepatitis B virus production by Boehmeria nivea root extract in HepG2 2.2.15 cells. World J. Gastroenterol. 2006, 12, 5721–5725. [Google Scholar] [CrossRef] [PubMed]
- Paiva, P.; Nonato, F.; Zafred, R.; Sousa, I.; Ruiz, A.L.; Foglio, M.A.; Carvalho, J.E. Anti-inflammatory activity of Boehmeria caudata sw. (Urticaceae) crude ethanol extract on croton oil mouse ear oedema. Planta Medica 2014, 80, 1528. [Google Scholar] [CrossRef]
- Sung, M.J.; Davaatseren, M.; Kim, S.H.; Kim, M.J.; Hwang, J.-T. Boehmeria nivea attenuates LPS-induced inflammatory markers by inhibiting p38 and jnk phosphorylations in raw264.7 macrophages. Pharm. Biol. 2013, 51, 1131–1136. [Google Scholar] [CrossRef] [PubMed]
- Zhu, S.; Tang, S.; Tang, Q.; Liu, T. Genome-wide transcriptional changes of ramie (Boehmeria nivea L. Gaud) in response to root-lesion nematode infection. Gene 2014, 552, 67–74. [Google Scholar] [CrossRef] [PubMed]
- Liu, T.; Zhu, S.; Tang, Q.; Tang, S. Genome-wide transcriptomic profiling of ramie (Boehmeria nivea L. Gaud) in response to cadmium stress. Gene 2015, 558, 131–137. [Google Scholar] [CrossRef] [PubMed]
- Huang, X.; Chen, J.; Bao, Y.; Liu, L.; Jiang, H.; An, X.; Dai, L.; Wang, B.; Peng, D. Transcript profiling reveals auxin and cytokinin signaling pathways and transcription regulation during in vitro organogenesis of ramie (Boehmeria nivea L. Gaud). PLoS ONE 2014, 9. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Liu, F.; Tang, Y.; Yuan, Y.; Guo, Q. Transcriptome sequencing and profiling of expressed genes in phloem and xylem of ramie (Boehmeria nivea L. Gaud). PLoS ONE 2014, 9. [Google Scholar] [CrossRef] [PubMed]
- Liu, T.; Tang, S.; Zhu, S.; Tang, Q.; Zheng, X. Transcriptome comparison reveals the patterns of selection in domesticated and wild ramie (Boehmeria nivea L. Gaud). Plant Mol. Biol. 2014, 86, 85–92. [Google Scholar] [CrossRef] [PubMed]
- Staff, P.O. Transcriptome sequencing and profiling of expressed genes in phloem and xylem of ramie (Boehmeria nivea L. Gaud). PLoS ONE 2015, 10. [Google Scholar] [CrossRef]
- Chen, J.; Pei, Z.; Dai, L.; Wang, B.; Liu, L.; An, X.; Peng, D. Transcriptome profiling using pyrosequencing shows genes associated with bast fiber development in ramie (Boehmeria nivea L.). BMC Genom. 2014, 15. [Google Scholar] [CrossRef] [PubMed]
- Guerriero, G.; Mangeot-Peter, L.; Hausman, J.-F.; Legay, S. Extraction of High Quality RNA from Cannabis sativa Bast Fibres: A Vademecum for Molecular Biologists. Fibers 2016, 4, 23. [Google Scholar] [CrossRef]
- Zhang, N.; Deyholos, M.K. RNA-Seq analysis of the shoot apex of flax (Linum usitatissimum) to identify phloem fiber specification genes. Front. Plant Sci. 2016, 7. [Google Scholar] [CrossRef]
- Robertson, G.; Schein, J.; Chiu, R.; Corbett, R.; Field, M.; Jackman, S.D.; Mungall, K.; Lee, S.; Okada, H.M.; Qian, J.Q.; et al. De novo assembly and analysis of RNA-seq data. Nat. Methods 2010, 7, 909–912. [Google Scholar] [CrossRef] [PubMed]
- Simpson, J.T.; Wong, K.; Jackman, S.D.; Schein, J.E.; Jones, S.J.M.; Birol, I. ABYSS: A parallel assembler for short read sequence data. Genome Res. 2009, 19, 1117–1123. [Google Scholar] [CrossRef] [PubMed]
- Huang, X.; Madan, A. CAP3: A DNA sequence assembly program. Genome Res. 1999, 9, 868–877. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; Godzik, A. CD-HIT: A fast program for clustering and comparing large sets of protein or nucleotide sequences. Bioinformatics 2006, 22, 1658–1659. [Google Scholar] [CrossRef] [PubMed]
- Langmead, B.; Salzberg, S.L. Fast gapped-read alignment with bowtie 2. Nat. Methods 2012, 9, 357–359. [Google Scholar] [CrossRef] [PubMed]
- Liu, T.; Zhu, S.; Tang, Q.; Chen, P.; Yu, Y.; Tang, S. De novo assembly and characterization of transcriptome using illumina paired-end sequencing and identification of cesa gene in ramie (Boehmeria nivea L. Gaud). BMC Genom. 2013, 14. [Google Scholar] [CrossRef] [PubMed]
- Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. Basic local alignment search tool. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef]
- Lamesch, P.; Berardini, T.Z.; Li, D.H.; Swarbreck, D.; Wilks, C.; Sasidharan, R.; Muller, R.; Dreher, K.; Alexander, D.L.; Garcia-Hernandez, M.; et al. The arabidopsis information resource (tair): Improved gene annotation and new tools. Nucleic Acids Res. 2012, 40, D1202–D1210. [Google Scholar] [CrossRef] [PubMed]
- Ramírez-Sánchez, O.; Pérez-Rodríguez, P.; Delaye, L.; Tiessen, A. Plant Proteins Are Smaller Because They Are Encoded by Fewer Exons than Animal Proteins. Genom. Proteom. Bioinform. 2016, 14, 357–370. [Google Scholar]
- Michael, T.P.; Jackson, S. The First 50 Plant Genomes. Plant Genome 2013, 6. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
| Tissue | SRA 1 Accession | Number of Paired-End Reads | Read Length (bp 2) | Total Reads (Gbp 3) | Reference |
|---|---|---|---|---|---|
| shoot apex with young leaves and woody stem | ERR364387 | 5,571,079 | 73 | 0.813 | this study |
| whole plant excluding root | SRR546782 | 26,457,905 | 90 | 4.76 | [22] |
| phloem | SRR1380980 | 13,513,706 | 90 | 2.43 | [10] |
| xylem | SRR1380981 | 13,865,114 | 90 | 2.50 | [10] |
| whole plant excluding root | SRR1021565 | 28,165,784 | 101 | 5.69 | [7] |
| whole plant excluding root | SRR1021566 | 25,846,434 | 101 | 5.22 | [7] |
| whole plant excluding root; roots were infected by nematodes | SRR1021567 | 21,701,137 | 101 | 4.38 | [7] |
| whole plant excluding root; roots were infected by nematodes | SRR1021568 | 22,499,892 | 101 | 4.54 | [7] |
| Descriptor | Value |
|---|---|
| Total clean paired reads | 157,621,051 |
| Assembled scaffolds 1 | 70,271 |
| Longest scaffold (bp) | 22,363 |
| Smallest scaffold (bp) | 200 |
| N50 length (bp) | 1798 |
| N50 count | 14,016 |
| N90 length (bp) | 459 |
| N90 count | 43,828 |
| Total scaffolds length (bp) 1 | 77,022,208 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Al-Ani, L.; Deyholos, M.K. Transcriptome Assembly of the Bast Fiber Crop, Ramie, Boehmeria nivea (L.) Gaud. (Urticaceae). Fibers 2018, 6, 8. https://doi.org/10.3390/fib6010008
Al-Ani L, Deyholos MK. Transcriptome Assembly of the Bast Fiber Crop, Ramie, Boehmeria nivea (L.) Gaud. (Urticaceae). Fibers. 2018; 6(1):8. https://doi.org/10.3390/fib6010008
Chicago/Turabian StyleAl-Ani, Louay, and Michael K. Deyholos. 2018. "Transcriptome Assembly of the Bast Fiber Crop, Ramie, Boehmeria nivea (L.) Gaud. (Urticaceae)" Fibers 6, no. 1: 8. https://doi.org/10.3390/fib6010008
APA StyleAl-Ani, L., & Deyholos, M. K. (2018). Transcriptome Assembly of the Bast Fiber Crop, Ramie, Boehmeria nivea (L.) Gaud. (Urticaceae). Fibers, 6(1), 8. https://doi.org/10.3390/fib6010008