
Neurotoxicity in Marine Invertebrates: An Update

 ,
,  ,
,  , ,
, ,  and
and 
Abstract
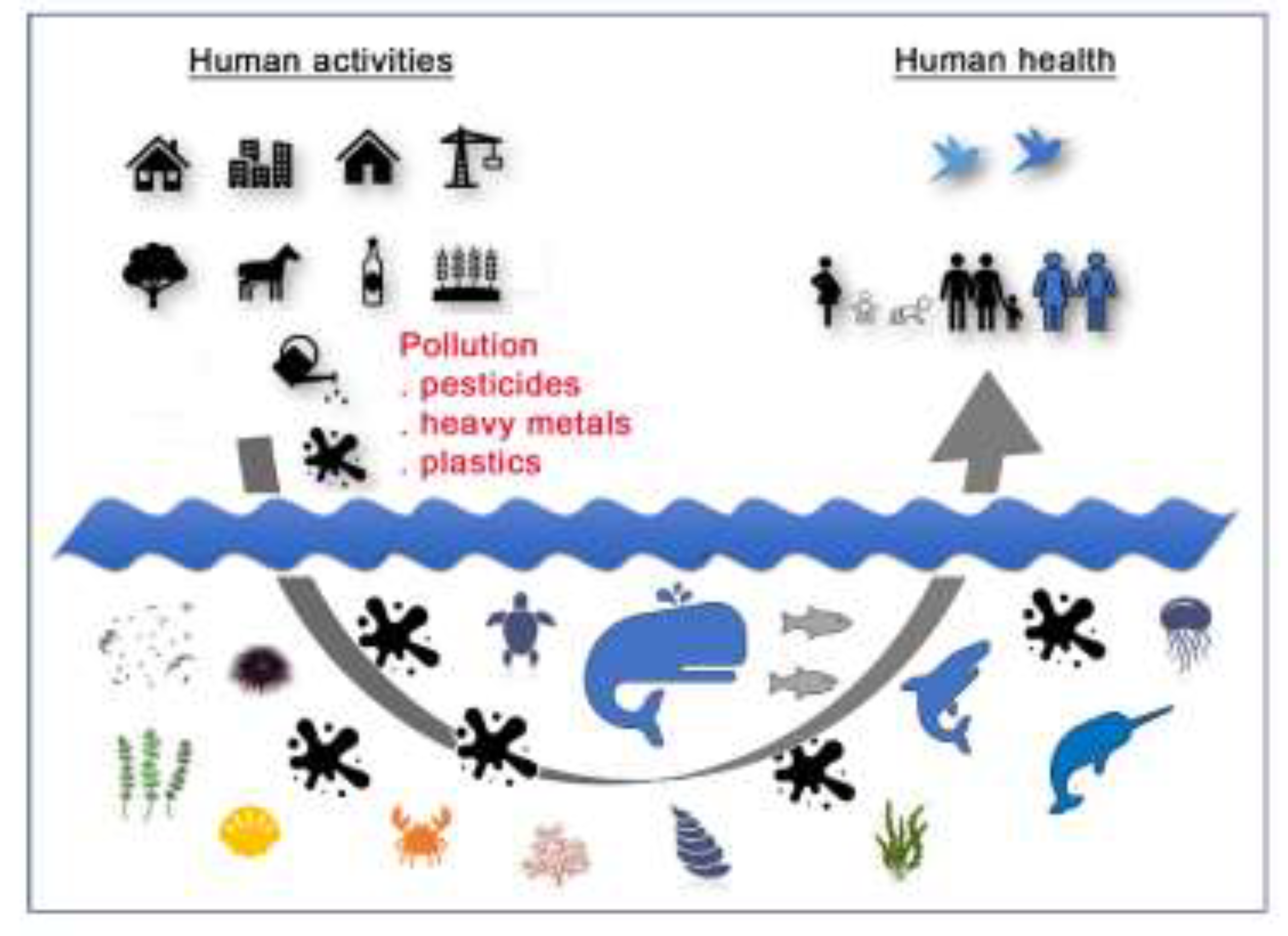
:Simple Summary
Abstract
1. Introduction
2. Acetylcholinesterase as Principal Biomarker of Neurotoxicity
3. Marine Pollutants: Heavy Metals
3.1. Essential Metals
3.2. Non-Essential Metals
4. Marine Pollutants: Pesticides
4.1. Echinoderms
4.2. Molluscs
4.3. Crustaceans
5. Marine Pollutants: Microplastics and Nanoplastics
6. Omics Studies
7. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Alava, J.J.; Cheung, W.W.L.; Ross, P.S.; Sumaila, U.R. Climate change-contaminant interactions in marine food webs: Toward a conceptual framework. Glob. Chang. Biol. 2017, 23, 3984–4001. [Google Scholar] [CrossRef]
- Cabral, H.; Fonseca, V.; Sousa, T.; Leal, M.C. Synergistic effects of climate change and marine pollution: An overlooked in-teraction in coastal and estuarine areas. Int. J. Environ. Res. Public Health 2019, 16, 2737. [Google Scholar] [CrossRef] [Green Version]
- Vonk, J.A.; Kraak, M.H.S. Herbicide Exposure and Toxicity to Aquatic Primary Producers. In Reviews of Environmental Contamination and Toxicology; (Continuation of Residue Reviews); Springer: Cham, Switzerland, 2020. [Google Scholar] [CrossRef]
- Modgil, S.; Lahiri, D.K.; Sharma, V.L.; Anand, A. Role of early life exposure and environment on neurodegeneration: Impli-cations on brain disorders. Transl. Neurodegener. 2014, 3, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Sharma, S.; Chatterjee, S. Microplastic pollution, a threat to marine ecosystem and human health: A short review. Environ. Sci. Pollut. Res. 2017, 24, 21530–21547. [Google Scholar] [CrossRef] [PubMed]
- Borja, A.; White, M.P.; Berdalet, E.; Bock, N.; Eatock, C.; Kristensen, P.; Leonard, A.; Lloret, J.; Pahl, S.; Parga, M.; et al. Moving Toward an Agenda on Ocean Health and Human Health in Europe. Front. Mar. Sci. 2020, 7, 7. [Google Scholar] [CrossRef] [Green Version]
- Iqubal, A.; Ahmed, M.; Ahmad, S.; Sahoo, C.R.; Iqubal, M.K.; Haque, S.E. Environmental neurotoxic pollutants: Review. Environ. Sci. Pollut. Res. 2020, 27, 41175–41198. [Google Scholar] [CrossRef]
- Mehler, M.F. Epigenetic principles and mechanisms underlying nervous system functions in health and disease. Prog. Neurobiol. 2008, 86, 305–341. [Google Scholar] [CrossRef] [Green Version]
- Dondero, F.; Banni, M.; Negri, A.; Boatti, L.; Dagnino, A.; Viarengo, A. Interactions of a pesticide/heavy metal mixture in marine bivalves: A transcriptomic assessment. BMC Genom. 2011, 12, 195. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bundschuh, M.; Zubrod, J.P.; Petschick, L.L.; Schulz, R. Multiple Stressors in Aquatic Ecosystems: Sublethal Effects of Tem-perature, Dissolved Organic Matter, Light and a Neonicotinoid Insecticide on Gammarids. Bull. Environ. Contam. Toxicol. 2020, 105, 345–350. [Google Scholar] [CrossRef]
- Altenburger, R.; Brack, W.; Burgess, R.M.; Busch, W.; Escher, B.I.; Focks, A.; Mark Hewitt, L.; Jacobsen, B.N.; de Alda, M.L.; Ait-Aissa, S.; et al. Future water quality monitoring: Improving the balance between exposure and toxicity assessments of re-al-world pollutant mixtures. Environ. Sci. Eur. 2019, 31, 1–17. [Google Scholar] [CrossRef] [Green Version]
- Schäfer, R.B.; Piggott, J.J. Advancing understanding and prediction in multiple stressor research through a mechanistic basis for null models. Glob. Chang. Biol. 2018, 24, 1817–1826. [Google Scholar] [CrossRef]
- Burke, R.; Angerer, L.; Elphick, M.; Humphrey, G.; Yaguchi, S.; Kiyama, T.; Liang, S.; Mu, X.; Agca, C.; Klein, W.; et al. A genomic view of the sea urchin nervous system. Dev. Biol. 2006, 300, 434–460. [Google Scholar] [CrossRef] [Green Version]
- Ullrich-Lüter, E.M.; Dupont, S.; Arboleda, E.; Hausen, H.; Arnone, M.I. Unique system of photoreceptors in sea urchin tube feet. Proc. Natl. Acad. Sci. USA 2011, 108, 8367–8372. [Google Scholar] [CrossRef] [Green Version]
- Fritsch, M.; Richter, S. The formation of the nervous system during larval development in Triops cancriformis (Bosc) (Crustacea, Branchiopoda): An immunohistochemical survey. J. Morphol. 2010, 271, 1457–1481. [Google Scholar] [CrossRef]
- Yurchenko, O.V.; Savelieva, A.V.; Kolotuchina, N.K.; Voronezhskaya, E.E.; Dyachuk, V.A. Peripheral sensory neurons govern development of the nervous system in bivalve larvae. EvoDevo 2019, 10, 1–9. [Google Scholar] [CrossRef]
- Christie, A.E.; Stanhope, M.E.; Gandler, H.I.; Lameyer, T.J.; Pascual, M.G.; Shea, D.N.; Yu, A.; Dickinson, P.S.; Hull, J.J. Molecular characterization of putative neuropeptide, amine, diffusible gas and small molecule transmitter biosynthetic enzymes in the eyestalk ganglia of the American lobster, Homarus americanus. Invertebr. Neurosci. 2018, 18, 1–29. [Google Scholar] [CrossRef]
- Spurgeon, D.J.; Jones, O.A.; Dorne, J.-L.C.; Svendsen, C.; Swain, S.; Stürzenbaum, S.R. Systems toxicology approaches for understanding the joint effects of environmental chemical mixtures. Sci. Total Environ. 2010, 408, 3725–3734. [Google Scholar] [CrossRef] [PubMed]
- Waters, M.D.; Fostel, J.M. Toxicogenomics and systems toxicology: Aims and prospects. Nat. Rev. Genet. 2004, 5, 936–948. [Google Scholar] [CrossRef]
- Watanabe, H.; Iguchi, T. Using ecotoxicogenomics to evaluate the impact of chemicals on aquatic organisms. Mar. Biol. 2005, 149, 107–115. [Google Scholar] [CrossRef]
- Fulton, M.H.; Key, P.B. Acetylcholinesterase inhibition in estuarine fish and invertebrates as an indicator of organophosphorus insecticide exposure and effects. Environ. Toxicol. Chem. 2001, 20, 37–45. [Google Scholar] [CrossRef]
- Fieber, L.A. Neurotransmitters and Neuropeptides of Invertebrates. Oxford Handb. Invertebr. Neurobiol. 2017, 284–300. [Google Scholar] [CrossRef]
- Díaz-Balzac, C.A.; García-Arrarás, J.E.; Díaz-Balzac, C.A.; García-Arrarás, J.E. Echinoderm Nervous System. In Oxford Research Encyclopedia; Oxford University Press: Oxford, UK, 2018; ISBN 9780190264086. [Google Scholar]
- Pezzementi, L.; Chatonnet, A. Evolution of cholinesterases in the animal kingdom. Chem. Interact. 2010, 187, 27–33. [Google Scholar] [CrossRef]
- Forget, J.; Bocquené, G. Partial purification and enzymatic characterization of acetylcholinesterase from the intertidal marine copepod Tigriopus brevicornis. Comp. Biochem. Physiol. B Biochem. Mol. Biol. 1999, 123, 345–350. [Google Scholar] [CrossRef]
- Jebali, J.; Khedher, S.B.; Sabbagh, M.; Kamel, N.; Banni, M.; Boussetta, H. Cholinesterase activity as biomarker of neuro-toxicity: Utility in the assessment of aquatic environment contamination. Rev. Gestão Costeira Integr. 2013, 13, 525–537. [Google Scholar] [CrossRef] [Green Version]
- Falugi, C.; Aluigi, M.; Chiantore, M.; Privitera, D.; Ramoino, P.; Gatti, M.; Fabrizi, A.; Pinsino, A.; Matranga, V. Toxicity of metal oxide nanoparticles in immune cells of the sea urchin. Mar. Environ. Res. 2012, 76, 114–121. [Google Scholar] [CrossRef] [PubMed]
- Gaitonde, D.; Sarkar, A.; Kaisary, S.; Silva, C.D.; Dias, C.; Rao, D.P.; Ray, D.; Nagarajan, R.; De Sousa, S.N.; Sarker, S.; et al. Acetylcholinesterase activities in marine snail (Cronia contracta) as a biomarker of neurotoxic contaminants along the Goa coast, West coast of India. Ecotoxicology 2006, 15, 353–358. [Google Scholar] [CrossRef] [PubMed]
- Caito, S.; Aschner, M. Neurotoxicity of Metals. In Handbook of Clinical Neurology; Lotti, M., Bleecker, M.L., Eds.; Elsevier BV: Amsterdam, The Netherlands, 2015; Volume 131, pp. 169–189. [Google Scholar]
- Doyle, D. Notoriety to respectability: A short history of arsenic prior to its present day use in haematology. Br. J. Haematol. 2009, 145, 309–317. [Google Scholar] [CrossRef]
- Thompson, E.D.; Hogstrand, C.; Glover, C.N. From sea squirts to squirrelfish: Facultative trace element hyperaccumulation in animals. Metallomics 2018, 10, 777–793. [Google Scholar] [CrossRef] [Green Version]
- Yllmaz, A.B.; Yanar, A.; Alkan, E.N. Review of heavy metal accumulation on aquatic environment in Northern East Medi-terrenean Sea part I: Some essential metals. Rev. Environ. Health 2017, 32, 119–163. [Google Scholar] [CrossRef] [PubMed]
- Ansari, T.M.; Marr, I.L.; Tariq, N. Heavy Metals in Marine Pollution Perspective–A Mini Review. J. Appl. Sci. 2003, 4, 1–20. [Google Scholar] [CrossRef] [Green Version]
- Sonone, S.S.; Jadhav, S.; Sankhla, M.S.; Kumar, R. Water contamination by heavy metals and their toxic effect on aquaculture and human health through food Chain. Lett. Appl. NanoBioScience 2021, 10, 2148–2166. [Google Scholar]
- Green, A.J.; Planchart, A. The Neurodevelopmental Toxicity of Heavy Metals: A Fish Perspective. Comp. Biochem. Physiol. C Toxicol. Pharmacol. 2018, 208, 12–19. [Google Scholar] [CrossRef]
- Li, L.; Yang, X. The Essential Element Manganese, Oxidative Stress, and Metabolic Diseases: Links and Interactions. Oxidative Med. Cell. Longev. 2018, 2018, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Balachandran, R.C.; Mukhopadhyay, S.; McBride, D.; Veevers, J.; Harrison, F.E.; Aschner, M.; Haynes, E.N.; Bowman, A.B. Brain manganese and the balance between essential roles and neurotoxicity. J. Biol. Chem. 2020, 295, 6312–6329. [Google Scholar] [CrossRef] [Green Version]
- Baden, S.P.; Neil, D.M. Accumulation of Manganese in the Haemolymph, Nerve and Muscle Tissue of Nephrops norvegicus (L.) and Its Effect on Neuromuscular Performance. Comp. Biochem. Physiol. Part A Mol. Integr. Physiol. 1998, 119, 351–359. [Google Scholar] [CrossRef]
- Baden, S.; Eriksson, S. Role, Routes and Effects of Manganese in Crustaceans. Oceanogr. Mar. Biol. 2006, 44, 61–84. [Google Scholar] [CrossRef]
- Holmes, J.M.; Gräns, A.-S.; Neil, D.M.; Baden, S.P. The effects of the metal ions Mn 2+ and Co 2+ on muscle contraction in the Norway lobster Nephrops norvegicus (L.). J. Comp. Physiol. B 1999, 169, 402–410. [Google Scholar] [CrossRef]
- Ochs, A.T.; Shields, J.D.; Rice, G.W.; Unger, M.A. Acute and long-term manganese exposure and subsequent accumulation in relation to idiopathic blindness in the American lobster, Homarus americanus. Aquat. Toxicol. 2020, 219, 105379. [Google Scholar] [CrossRef] [PubMed]
- Krång, A.-S.; Rosenqvist, G. Effects of manganese on chemically induced food search behaviour of the Norway lobster, Nephrops norvegicus (L.). Aquat. Toxicol. 2006, 78, 284–291. [Google Scholar] [CrossRef]
- Martin, K.; Huggins, T.; King, C.; Carroll, M.A.; Catapane, E.J. The neurotoxic effects of manganese on the dopaminergic innervation of the gill of the bivalve mollusc, Crassostrea virginica. Comp. Biochem. Physiol. Part C Toxicol. Pharmacol. 2008, 148, 152–159. [Google Scholar] [CrossRef] [Green Version]
- King, C.; Myrthil, M.; Carroll, M.A.; Catapane, E.J. Effects of p-Aminosalicylic acid on the Neurotoxicity of Manganese and Levels of Dopamine and Serotonin in the Nervous System and Innervated Organs of Crassostrea virginica. Vivo 2008, 29, 26–34. [Google Scholar]
- Catapane, E.J.; Nelson, M.; Adams, T.; Carroll, M.A. Innervation of Gill Lateral Cells in the Bivalve Mollusc Crassostrea virginica Affects Cellular Membrane Potential and Cilia Activity. J. Pharmacol. Rep. 2016, 1, 1–16. [Google Scholar]
- Sköld, H.N.; Baden, S.P.; Looström, J.; Eriksson, S.P.; Hernroth, B.E. Motoric impairment following manganese exposure in asteroid echinoderms. Aquat. Toxicol. 2015, 167, 31–37. [Google Scholar] [CrossRef]
- Youdim, M.B.H. Brain iron deficiency and excess; cognitive impairment and neurodegenration with involvement of striatum and hippocampus. Neurotox. Res. 2008, 14, 45–56. [Google Scholar] [CrossRef]
- Levine, S.M.; Macklin, W.B. Iron-enriched oligodendrocytes: A reexamination of their spatial distribution. J. Neurosci. Res. 1990, 26, 508–512. [Google Scholar] [CrossRef] [PubMed]
- Halliwell, B.; Gutteridge, J.M.C. Oxygen toxicity, oxygen radicals, transition metals and disease. Biochem. J. 1984, 219, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Andersen, H.H.; Johnsen, K.B.; Moos, T. Iron deposits in the chronically inflamed central nervous system and contributes to neurodegeneration. Cell. Mol. Life Sci. 2014, 71, 1607–1622. [Google Scholar] [CrossRef] [Green Version]
- Liu, Z.; Li, M.; Yi, Q.; Wang, L.; Song, L. The Neuroendocrine-Immune Regulation in Response to Environmental Stress in Marine Bivalves. Front. Physiol. 2018, 9, 1456. [Google Scholar] [CrossRef] [Green Version]
- Choi, S.; Hong, D.K.; Choi, B.Y.; Suh, S.W. Zinc in the Brain: Friend or Foe? Int. J. Mol. Sci. 2020, 21, 8941. [Google Scholar] [CrossRef] [PubMed]
- Moore, J.K.; Braucher, O. Observations of dissolved iron concentrations in the World Ocean: Implications and constraints for ocean biogeochemical models. Biogeosciences Discuss. 2007, 4, 1241–1277. [Google Scholar]
- Gledhiir, M.; Buck, K.N. The organic complexation of iron in the marine environment: A review. Front. Microbiol. 2012, 3, 1–17. [Google Scholar]
- Najimi, S.; Bouhaimi, A.; Daubèze, M.; Zekhnini, A.; Pellerin, J.; Narbonne, J.F.; Moukrim, A. Use of acetylcholinesterase in Perna perna and Mytilus galloprovincialis as a biomarker of pollution in Agadir Marine Bay (South of Morocco). Bull. Environ. Contam. Toxicol. 1997, 58, 901–908. [Google Scholar] [CrossRef]
- Timourian, H. The effect of zinc on sea urchin morphogenesis. J. Exp. Zool. 1968, 169, 121–131. [Google Scholar] [CrossRef] [PubMed]
- Poustka, A.J.; Kühn, A.; Groth, D.; Weise, V.; Yaguchi, S.; Burke, R.D.; Herwig, R.; Lehrach, H.; Panopoulou, G. A global view of gene expression in lithium and zinc treated sea urchin embryos: New components of gene regulatory networks. Genome Biol. 2007, 8, R85. [Google Scholar] [CrossRef] [Green Version]
- Russo, R.; Bonaventura, R.; Chiaramonte, M.; Costa, C.; Matranga, V.; Zito, F. Response to metals treatment of Fra1, a member of the AP-1 transcription factor family, in P. lividus sea urchin embryos. Mar. Environ. Res. 2018, 139, 99–112. [Google Scholar] [CrossRef] [PubMed]
- Poustka, A.J.; Kuhn, A.; Radosavljevic, V.; Wellenreuther, R.; Lehrach, H.; Panopoulou, G. On the origin of the chordate central nervous system: Expression of onecut in the sea urchin embryo. Evol. Dev. 2004, 6, 227–236. [Google Scholar] [CrossRef]
- Kropp, P.A.; Gannon, M. Onecut transcription factors in development and disease. Trends Dev. Biol. 2016, 9, 43–57. [Google Scholar]
- Bulcke, F.; Dringen, R.; Scheiber, I.F. Neurotoxicity of Copper. In Neurotoxicity of Metals. Advances in Neurobiology; Aschner, M., Costa, L., Eds.; Springer: Cham, Switzerland, 2017; Volume 18. [Google Scholar] [CrossRef]
- Hsu, H.-W.; Bondy, S.C.; Kitazawa, M. Environmental and Dietary Exposure to Copper and Its Cellular Mechanisms Linking to Alzheimer’s Disease. Toxicol. Sci. 2018, 163, 338–345. [Google Scholar] [CrossRef]
- Cunha, I.; Mangas-Ramírez, E.; Guilhermino, L. Effects of copper and cadmium on cholinesterase and glutathione S-transferase activities of two marine gastropods (Monodonta lineata and Nucella lapillus). Comp. Biochem. Physiol. Part C Toxicol. Pharmacol. 2007, 145, 648–657. [Google Scholar] [CrossRef] [PubMed]
- Bocquené, G.; Galgani, F.; Truquet, P. Characterization and assay conditions for use of ACHe activity from several marine species in pollution monitoring. Mar. Environ. Res. 1990, 30, 75–89. [Google Scholar] [CrossRef]
- Curtis, T.M.; Williamson, R.; Depledge, M.H. Simultaneous, long-term monitoring of valve and cardiac activity in the blue mussel Mytilus edulis exposed to copper. Mar. Biol. 2000, 136, 837–846. [Google Scholar] [CrossRef]
- Brown, R.; Galloway, T.; Lowe, D.; Browne, M.; Dissanayake, A.; Jones, M.; Depledge, M. Differential sensitivity of three marine invertebrates to copper assessed using multiple biomarkers. Aquat. Toxicol. 2004, 66, 267–278. [Google Scholar] [CrossRef]
- Orihuela, R.; Domènech, J.; Bofill, R.; You, C.; Mackay, E.A.; Kägi, J.H.R.; Capdevila, M.; Atrian, S. The metal-binding features of the recombinant mussel Mytilus edulis MT-10-IV metallothionein. JBIC J. Biol. Inorg. Chem. 2008, 13, 801–812. [Google Scholar] [CrossRef]
- Cammilleri, G.; Galluzzo, P.; Pulvirenti, A.; Giangrosso, I.E.; Dico, G.M.L.; Montana, G.; Lampiasi, N.; Mobilia, M.A.; Lastra, A.; Vazzana, M.; et al. Toxic mineral elements in Mytilus galloprovincialis from Sicilian coasts (Southern Italy). Nat. Prod. Res. 2020, 34, 177–182. [Google Scholar] [CrossRef] [PubMed]
- Calvo, J.; Jung, H.; Meloni, G. Copper metallothioneins. IUBMB Life 2017, 69, 236–245. [Google Scholar] [CrossRef] [Green Version]
- Khedher, S.B.; Houas, Z.; Boussetta, H. Cholinesterase activity and histopatological changes in the Mediterranean crab, Carcinus maenas, exposed to environmental contaminants. Int. J. Aquat. Biol. 2017, 5, 348–359. [Google Scholar]
- Elumalai, M.; Antunes, C.; Guilhermino, L. Effects of Single Metals and their Mixtures on Selected Enzymes of Carcinus Maenas. Water Air Soil Pollut. 2002, 141, 273–280. [Google Scholar] [CrossRef]
- Oliva, M.; De Marchi, L.; Cuccaro, A.; Casu, V.; Tardelli, F.; Monni, G.; Freitas, R.; Caliani, I.; Fossi, M.; Fratini, S.; et al. Effects of copper on larvae of the marbled crab Pachygrapsus marmoratus (Decapoda, Grapsidae): Toxicity test and biochemical marker responses. Comp. Biochem. Physiol. Part C Toxicol. Pharmacol. 2019, 223, 71–77. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.-H.; Kang, J.-C. The immune responses in juvenile rockfish, Sebastes schlegelii for the stress by the exposure to the dietary lead (II). Environ. Toxicol. Pharmacol. 2016, 46, 211–216. [Google Scholar] [CrossRef]
- Lee, J.-W.; Choi, H.; Hwang, U.-K.; Kang, J.-C.; Kang, Y.J.; Kim, K.I.; Kim, J.-H. Toxic effects of lead exposure on bioaccumulation, oxidative stress, neurotoxicity, and immune responses in fish: A review. Environ. Toxicol. Pharmacol. 2019, 68, 101–108. [Google Scholar] [CrossRef]
- Rocha, A.; Trujillo, K.A. Neurotoxicity of low-level lead exposure: History, mechanisms of action, and behavioral effects in humans and preclinical models. Physiol. Behav. 2017, 176, 139–148. [Google Scholar] [CrossRef]
- Wang, B.; Du, Y. Cadmium and Its Neurotoxic Effects. Oxidative Med. Cell. Longev. 2013, 2013, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Chin-Chan, M.; Navarro-Yepes, J.; Quintanilla-Vega, B. Environmental pollutants as risk factors for neurodegenerative dis-orders: Alzheimer and Parkinson diseases. Front. Cell. Neurosci. 2015, 9, 1–22. [Google Scholar] [CrossRef] [Green Version]
- Roccheri, M.C.; Matranga, V. Cellular, biochemical and molecular effects of cadmium on marine invertebrates: Focus on Pa-racentrotus lividus sea urchin development. In Cadmium in the Environment; Reini, G.P., Ed.; Nova Science Publishers Inc.: New York, NY, USA, 2009; pp. 1–30. [Google Scholar]
- Bonaventura, R.; Russo, R.; Zito, F.; Matranga, V. Combined Effects of Cadmium and UVB Radiation on Sea Urchin Embryos: Skeleton Impairment Parallels p38 MAPK Activation and Stress Genes Overexpression. Chem. Res. Toxicol. 2015, 28, 1060–1069. [Google Scholar] [CrossRef] [PubMed]
- Bonacci, S.; Corsi, I.; Focardi, S. Cholinesterase activities in the scallop Pecten jacobaeus: Characterization and effects of ex-posure to aquatic contaminants. Sci. Total Environ. 2008, 392, 99–109. [Google Scholar]
- Mansour, C.; Guibbolini, M.; Hacene, O.R.; Mosbahi, D.S.; Faverney, C.R.-D. Oxidative Stress and Damage Biomarkers in Clam Ruditapes decussatus Exposed to a Polluted Site: The Reliable Biomonitoring Tools in Hot and Cold Seasons. Arch. Environ. Contam. Toxicol. 2020, 78, 478–494. [Google Scholar] [CrossRef] [PubMed]
- Aaseth, J.; Wallace, D.R.; Vejrup, K.; Alexander, J. Methylmercury and developmental neurotoxicity: A global concern. Curr. Opin. Toxicol. 2020, 19, 80–87. [Google Scholar] [CrossRef]
- Bjørklund, G.; Mutter, J.; Aaseth, J. Metal chelators and neurotoxicity: Lead, mercury, and arsenic. Arch. Toxicol. 2017, 91, 3787–3797. [Google Scholar] [CrossRef]
- Chiarelli, R.; Roccheri, M.C. Marine Invertebrates as Bioindicators of Heavy Metal Pollution. Open J. Met. 2014, 4, 93–106. [Google Scholar] [CrossRef] [Green Version]
- Chalghmi, H.; Bourdineaud, J.-P.; Haouas, Z.; Gourves, P.-Y.; Zrafi, I.; Saidane-Mosbahi, D. Transcriptomic, Biochemical, and Histopathological Responses of the Clam Ruditapes decussatus from a Metal-Contaminated Tunis Lagoon. Arch. Environ. Contam. Toxicol. 2016, 70, 241–256. [Google Scholar] [CrossRef]
- Prakash, C.; Soni, M.; Kumar, V. Mitochondrial oxidative stress and dysfunction in arsenic neurotoxicity: A review. J. Appl. Toxicol. 2015, 36, 179–188. [Google Scholar] [CrossRef] [PubMed]
- Neff, J.M. Ecotoxicology of arsenic in the marine environment. Environ. Toxicol. Chem. 1997, 16, 917–927. [Google Scholar] [CrossRef]
- Freitas, A.R.; Coppsola, F.; De Marchi, L.; Codela, V.; Pretti, C.; Chiellini, F.; Polese, G.S. The influence of Arsenic on the toxicity of carbon nanoparticles in bivalves. J. Hazard. Mater. 2018, 358, 484–493. [Google Scholar] [CrossRef]
- Blewett, T.A.; Leonard, E.M. Mechanisms of nickel toxicity to fish and invertebrates in marine and estuarine waters. Environ. Pollut. 2017, 223, 311–322. [Google Scholar] [CrossRef]
- Attig, H.; Dagnino, A.; Negri, A.; Jebali, J.; Boussetta, H.; Viarengo, A.; Dondero, F.; Banni, M. Uptake and biochemical re-sponses of mussels Mytilus galloprovincialis exposed to sublethal nickel concentrations. Ecotoxicol. Environ. Saf. 2010, 73, 1712–1719. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.; Wang, G. Oxidative damage effects in the copepod Tigriopus japonicus Mori experimentally exposed to nickel. Ecotoxicology 2009, 19, 273–284. [Google Scholar] [CrossRef] [Green Version]
- Olsvik, P.A.; Larsen, A.K.; Berntssen, M.H.G.; Goksøyr, A.; Karlsen, O.A.; Yadetie, F.; Sanden, M.; Kristensen, T. Effects of Agricultural Pesticides in Aquafeeds on Wild Fish Feeding on Leftover Pellets Near Fish Farms. Front. Genet. 2019, 10, 1–18. [Google Scholar] [CrossRef]
- Richardson, J.R.; Fitsanakis, V.; Westerink, R.H.S.; Kanthasamy, A.G. Neurotoxicity of pesticides. Acta Neuropathol. 2019, 138, 343–362. [Google Scholar] [CrossRef]
- Jayaraj, R.; Megha, P.; Sreedev, P. Review Article. Organochlorine pesticides, their toxic effects on living organisms and their fate in the environment. Interdiscip. Toxicol. 2016, 9, 90–100. [Google Scholar] [CrossRef] [Green Version]
- Kwong, T.C. Organophosphate Pesticides: Biochemistry and Clinical Toxicology. Ther. Drug Monit. 2002, 24, 144–149. [Google Scholar] [CrossRef]
- Zhang, X.; Xu, L.; Tian, H.; Wang, C.; Wang, W.; Ru, S. Monocrotophos, an organophosphorus insecticide, disrupts the ex-pression of HpNetrin and its receptor neogenin during early development in the sea urchin (Hemicentrotus pulcherrimus). Neurotoxicology 2017, 62, 130–137. [Google Scholar] [CrossRef]
- Yao, D.; Ru, S.; Katow, H. The neurotoxic effects of monocrotophos on the formation of the serotonergic nervous system and swimming activity in the larvae of the sea urchin Hemicentrotus pulcherrimus. Environ. Toxicol. Pharmacol. 2010, 30, 181–187. [Google Scholar] [CrossRef]
- Zhang, X.; Li, S.; Wang, C.; Tian, H.; Wang, W.; Ru, S. Effects of monocrotophos pesticide on cholinergic and dopaminergic neurotransmitter systems during early development in the sea urchin Hemicentrotus pulcherrimus. Toxicol. Appl. Pharmacol. 2017, 328, 46–53. [Google Scholar] [CrossRef]
- Buznikov, G.A.; Nikitina, L.A.; Bezuglov, V.V.; Lauder, J.M.; Padilla, S.; Slotkin, T.A. An invertebrate model of the devel-opmental neurotoxicity of insecticides: Effects of chlorpyrifos and dieldrin in sea urchin embryos and larvae. Environ. Health Perspect. 2001, 109, 651–661. [Google Scholar] [CrossRef] [PubMed]
- Amaroli, A.; Aluigi, M.G.; Falugi, C.; Chessa, M.G. Effects of the neurotoxic thionophosphate pesticide chlorpyrifos on dif-ferentiating alternative models. Chemosphere 2013, 90, 2115–2122. [Google Scholar] [CrossRef] [PubMed]
- Pesando, D.; Huitorel, P.; Dolcini, V.; Angelini, C.; Guidetti, P.; Falugi, C. Biological targets of neurotoxic pesticides analysed by alteration of developmental events in the Mediterranean sea urchin, Paracentrotus lividus. Mar. Environ. Res. 2003, 55, 39–57. [Google Scholar] [CrossRef]
- Le Bris, H.; Maffart, P.; Bocquené, G.; Buchet, V.; Galgani, F.; Blanc, G. Laboratory study on the effect of dichlorvos on two commercial bivalves. Aquaculture 1995, 138, 139–144. [Google Scholar] [CrossRef]
- Anguiano, G.A.; Amador, A.; Moreno-Legorreta, M.; Arcos-Ortega, F.; Vazquez-Boucard, C. Effects of exposure to oxamyl, carbofuran, dichlorvos, and lindane on acetylcholinesterase activity in the gills of the Pacific oyster Crassostrea gigas. Environ. Toxicol. 2009, 25, 327–332. [Google Scholar] [CrossRef]
- El Ayari, T.; Mhadhbi, L.; Debara, N.; Znati, M.; Dab, H. The detoxifying effect of Polygonum equisetiforme extracts against dichlorvos (DDVP)-induced oxidative stress and neurotoxicity in the commercial clam Ruditapes decussatus. Environ. Sci. Pollut. Res. 2020, 27, 10449–10458. [Google Scholar] [CrossRef]
- McHenery, J.G.; Linley-Adams, G.E.; Moore, D.C.; Rodger, G.K.; Davies, I.M. Experimental and field studies of effects of di-chlorvos exposure on acetylcholinesterase activity in the gills of the mussel, Mytilus edulis L. Aquat. Toxicol. 1997, 38, 125–143. [Google Scholar] [CrossRef]
- Bocquené, G.; Roig, A.; Fournier, D. Cholinesterases from the common oyster (Crassostrea gigas). Evidence for the presence of a soluble acetylcholinesterase insensitive to organophosphate and carbamate inhibitors. FEBS Lett. 1997, 407, 261–266. [Google Scholar] [CrossRef] [Green Version]
- Canty, M.N.; Hagger, J.A.; Moore, R.T.B.; Cooper, L.; Galloway, T.S. Sublethal impact of short term exposure to the organo-phosphate pesticide azamethiphos in the marine mollusc Mytilus edulis. Mar. Pollut. Bull. 2007, 54, 396–402. [Google Scholar] [CrossRef]
- Galloway, T.S.; Millward, N.; Browne, M.A.; Depledge, M.H. Rapid assessment of organophosphorous/carbamate exposure in the bivalve mollusc Mytilus edulis using combined esterase activities as biomarkers. Aquat. Toxicol. 2002, 61, 169–180. [Google Scholar] [CrossRef]
- Rickwood, C.J.; Galloway, T.S. Acetylcholinesterase inhibition as a biomarker of adverse effect: A study of Mytilus edulis ex-posed to the priority pollutant chlorfenvinphos. Aquat. Toxicol. 2004, 67, 45–56. [Google Scholar] [CrossRef] [PubMed]
- Srivatsan, M. Effects of organophosphates on cholinesterase activity and neurite regeneration in Aplysia. Chem. Interact. 1999, 119, 371–378. [Google Scholar] [CrossRef]
- Filbert, M.G.; Apland, J.P.; Petrali, J.P.; Adler, M. Paraoxon block of chloride conductance in cell R2 of Aplysia californica. Brain Res. Bull. 1992, 28, 473–477. [Google Scholar] [CrossRef]
- Chen, Y.-Y.; Huang, L.; Zhang, Y.; Ke, C.-H.; Huang, H.-Q. Differential expression profile of membrane proteins in Aplysia pleural–pedal ganglia under the stress of methyl parathion. Environ. Sci. Pollut. Res. 2014, 21, 3371–3385. [Google Scholar] [CrossRef]
- Juhel, G.; Bayen, S.; Goh, C.; Lee, W.K.; Kelly, B.C. Use of a suite of biomarkers to assess the effects of carbamazepine, bisphenol A, atrazine, and their mixtures on green mussels, Perna viridis. Environ. Toxicol. Chem. 2017, 36, 429–441. [Google Scholar] [CrossRef] [PubMed]
- McHenery, J.G.; Saward, D.; Seaton, D.D. Lethal and sub-lethal effects of the salmon delousing agent dichlorvos on the larvae of the lobster (Homarus gammarus L.) and herring (Clupea harengus L.). Aquaculture 1991, 98, 331–347. [Google Scholar] [CrossRef]
- Mchenery, J.G.; Francis, C.; Davies, I.M. Threshold toxicity and repeated exposure studies of dichlorvos to the larvae of the common lobster (Homarus gammarus L.). Aquat. Toxicol. 1996, 34, 237–251. [Google Scholar]
- Murison, D.J.; Moore, D.C.; McHenery, J.G.; Robertson, N.A.; Davies, I.M. Epiphytic invertebrate assemblages and dichlorvos usage at salmon farms. Aquaculture 1997, 159, 53–66. [Google Scholar] [CrossRef]
- La Parra, L.G.-D.; Bautista-Covarrubias, J.; La Rosa, N.R.-D.; Betancourt-Lozano, M.; Guilhermino, L. Effects of methamidophos on acetylcholinesterase activity, behavior, and feeding rate of the white shrimp (Litopenaeus vannamei). Ecotoxicol. Environ. Saf. 2006, 65, 372–380. [Google Scholar] [CrossRef] [PubMed]
- Frasco, M.F.; Fournier, D.; Carvalho, F.; Guilhermino, L. Cholinesterase from the common prawn (Palaemon serratus) eyes: Catalytic properties and sensitivity to organophosphate and carbamate compounds. Aquat. Toxicol. 2006, 77, 412–421. [Google Scholar] [CrossRef] [PubMed]
- Frasco, M.F.; Fournier, D.; Carvalho, F.; Guilhermino, L. Does mercury interact with the inhibitory effect of dichlorvos on Palaemon serratus (Crustacea: Decapoda) cholinesterase? Sci. Total Environ. 2008, 404, 88–93. [Google Scholar] [CrossRef]
- Oliveira, C.; Almeida, J.R.; Guilhermino, L.; Soares, A.M.V.M.; Gravato, C. Swimming velocity, avoidance behavior and bi-omarkers in Palaemon serratus exposed to fenitrothion. Chemosphere 2013, 90, 936–944. [Google Scholar] [CrossRef]
- Couillard, C.; Burridge, L. Sublethal exposure to azamethiphos causes neurotoxicity, altered energy allocation and high mortality during simulated live transport in American lobster. Ecotoxicol. Environ. Saf. 2015, 115, 291–299. [Google Scholar] [CrossRef]
- Fallang, A.; Ramsay, J.M.; Sevatdal, S.; Burka, J.F.; Jewess, P.; Hammell, K.L.; Horsberg, T.E. Evidence for occurrence of an organophosphate-resistant type of acetylcholinesterase in strains of sea lice (Lepeophtheirus salmonis Krøyer). Pest Manag. Sci. 2004, 60, 1163–1170. [Google Scholar] [CrossRef] [PubMed]
- Varó, I.; Redon, S.; Roger, E.M.G.; Amat, F.; Guinot, D.; Serrano, R.; Navarro, J.C. Aquatic pollution may favor the success of the invasive species A. franciscana. Aquat. Toxicol. 2015, 161, 208–220. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Duarte-Restrepo, E.; Jaramillo-Colorado, B.E.; Duarte-Jaramillo, L. Effects of chlorpyrifos on the crustacean Litopenaeus vannamei. PLoS ONE 2020, 15, e0231310. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gambardella, C.; Nichino, D.; Iacometti, C.; Ferrando, S.; Falugi, C.; Faimali, M. Comparison with cholinesterase inhibitors shows that most inhibition is exerted on AChE activity SC. Aquat. Toxicol. 2018, 196, 79–89. [Google Scholar] [CrossRef]
- Lee, J.W.; Kim, B.-M.; Jeong, C.-B.; Won, E.-J.; Rhee, J.-S.; Lee, J.-S. Inhibitory effects of biocides on transcription and protein activity of acetylcholinesterase in the intertidal copepod Tigriopus japonicus. Comp. Biochem. Physiol. Part C Toxicol. Pharmacol. 2015, 167, 147–156. [Google Scholar] [CrossRef] [PubMed]
- Oliveira, C.; Almeida, J.; Guilhermino, L.; Soares, A.M.; Gravato, C.A. Acute effects of deltamethrin on swimming velocity and biomarkers of the common prawn Palaemon serratus. Aquat. Toxicol. 2012, 2012, 209–216. [Google Scholar] [CrossRef]
- Tu, H.T.; Silvestre, F.; De Meulder, B.; Thome, J.-P.; Phuong, N.T.; Kestemont, P. Combined effects of deltamethrin, temperature and salinity on oxidative stress biomarkers and acetylcholinesterase activity in the black tiger shrimp (Penaeus monodon). Chemosphere 2012, 86, 83–91. [Google Scholar] [CrossRef] [PubMed]
- Alimba, C.G.; Faggio, C. Microplastics in the marine environment: Current trends in environmental pollution and mechanisms of toxicological profile. Environ. Toxicol. Pharmacol. 2019, 68, 61–74. [Google Scholar] [CrossRef]
- Sana, S.S.; Dogiparthi, L.K.; Gangadhar, L.; Chakravorty, A.; Abhishek, N. Effects of microplastics and nanoplastics on marine environment and human health. Environ. Sci. Pollut. Res. 2020, 27, 44743–44756. [Google Scholar] [CrossRef]
- Chae, Y.; An, Y.-J. Effects of micro- and nanoplastics on aquatic ecosystems: Current research trends and perspectives. Mar. Pollut. Bull. 2017, 124, 624–632. [Google Scholar] [CrossRef] [PubMed]
- Prüst, M.; Meijer, J.; Westerink, R.H.S. The plastic brain: Neurotoxicity of micro and nanoplastics. Part. Fibre Toxicol. 2020, 17, 1–16. [Google Scholar] [CrossRef]
- de Sá, L.C.; Oliveira, M.; Ribeiro, F.; Rocha, T.L.; Futter, M.N. Studies of the effects of microplastics on aquatic organisms: What do we know and where should we focus our efforts in the future? Sci. Total Environ. 2018, 645, 1029–1039. [Google Scholar] [CrossRef] [PubMed]
- Avio, C.G.; Gorbi, S.; Milan, M.; Benedetti, M.; Fattorini, D.; D’Errico, G.; Pauletto, M.; Bargelloni, L.; Regoli, F. Pollutants bioavailability and toxicological risk from microplastics to marine mussels. Environ. Pollut. 2015, 198, 211–222. [Google Scholar] [CrossRef]
- Brandts, I.; Teles, M.; Gonçalves, A.P.; Barreto, A.; Franco-Martinez, L.; Tvarijonaviciute, A.; Martins, M.A.; Soares, A.M.V.M.; Tort, L.; Oliveira, M. Effects of nanoplastics on Mytilus galloprovincialis after individual and combined exposure with car-bamazepine. Sci. Total Environ. 2018, 643, 775–784. [Google Scholar] [CrossRef]
- Ribeiro, S.; Torres, T.; Martins, R.; Santos, M.M. Toxicity screening of Diclofenac, Propranolol, Sertraline and Simvastatin using Danio rerio and Paracentrotus lividus embryo bioassays. Ecotoxicol. Environ. Saf. 2015, 114, 67–74. [Google Scholar] [CrossRef] [Green Version]
- O’Donovan, S.; Mestre, N.C.; Abel, S.; Fonseca, T.G.; Carteny, C.C.; Willems, T.; Prinsen, E.; Cormier, B.; Keiter, S.S.; Bebianno, M.J. Effects of the UV filter, oxybenzone, adsorbed to microplastics in the clam Scrobicularia plana. Sci. Total Environ. 2020, 733, 139102. [Google Scholar] [CrossRef]
- Tlili, S.; Jemai, D.; Brinis, S.; Regaya, I. Microplastics mixture exposure at environmentally relevant conditions induce oxidative stress and neurotoxicity in the wedge clam Donax trunculus. Chemosphere 2020, 258, 127344. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Feng, C.; Wu, Y.; Guo, X. Impacts of nanoplastics on bivalve: Fluorescence tracing of organ accumulation, oxidative stress and damage. J. Hazard. Mater. 2020, 392, 122418. [Google Scholar] [CrossRef] [PubMed]
- Tang, Y.; Zhou, W.; Sun, S.; Du, X.; Han, Y.; Shi, W.; Liu, G. Immunotoxicity and neurotoxicity of bisphenol A and microplastics alone or in combination to a bivalve species, Tegillarca granosa. Environ. Pollut. 2020, 265, 115115. [Google Scholar] [CrossRef]
- Gambardella, C.; Morgana, S.; Ferrando, S.; Bramini, M.; Piazza, V.; Costa, E.; Garaventa, F.; Faimali, M. Effects of polystyrene microbeads in marine planktonic crustaceans. Ecotoxicol. Environ. Saf. 2017, 145, 250–257. [Google Scholar] [CrossRef] [PubMed]
- Varó, I.; Perini, A.; Torreblanca, A.; Garcia, Y.; Bergami, E.; Vannuccini, M.L.; Corsi, I. Time-dependent effects of polystyrene nanoparticles in brine shrimp Artemia franciscana at physiological, biochemical and molecular levels. Sci. Total Environ. 2019, 675, 570–580. [Google Scholar] [CrossRef]
- Shi, B.; Huang, Z.; Xiang, X.; Huang, M.; Wang, W.X.; Ke, C. Transcriptome analysis of the key role of GAT2 gene in the hy-per-accumulation of copper in the oyster Crassostrea angulata. Sci. Rep. 2015, 5, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Huster, D.; Purnat, T.D.; Burkhead, J.L.; Ralle, M.; Fiehn, O.; Stuckert, F.; Olson, N.E.; Teupser, D.; Lutsenko, S. High Copper Selectively Alters Lipid Metabolism and Cell Cycle Machinery in the Mouse Model of Wilson Disease. J. Biol. Chem. 2007, 282, 8343–8355. [Google Scholar] [CrossRef] [Green Version]
- Filimonova, V.; Gonçalves, F.; Marques, J.C.; De Troch, M.; Gonçalves, A.M. Fatty acid profiling as bioindicator of chemical stress in marine organisms: A review. Ecol. Indic. 2016, 67, 657–672. [Google Scholar] [CrossRef]
- Naffaa, V.; Laprévote, O.; Schang, A.-L. Effects of endocrine disrupting chemicals on myelin development and diseases. NeuroToxicology 2021, 83, 51–68. [Google Scholar] [CrossRef]
- Chan, C.Y.; Wang, W.-X. A lipidomic approach to understand copper resilience in oyster Crassostrea hongkongensis. Aquat. Toxicol. 2018, 204, 160–170. [Google Scholar] [CrossRef] [PubMed]
- Tollefsen, K.E.; Song, Y.; Høgåsen, T.; Øverjordet, I.B.; Altin, D.; Hansen, B.H. Mortality and transcriptional effects of inorganic mercury in the marine copepod Calanus finmarchicus. J. Toxicol. Environ. Health Part A 2017, 80, 845–861. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cappello, T.; Vitale, V.; Oliva, S.; Villari, V.; Mauceri, A.; Fasulo, S.; Maisano, M. Alteration of neurotransmission and skele-togenesis in sea urchin Arbacia lixula embryos exposed to copper oxide nanoparticles. Comp. Biochem. Physiol. Part C Toxicol. Pharmacol. 2017, 199, 20–27. [Google Scholar] [CrossRef]
- Maisano, M.; Cappello, T.; Catanese, E.; Vitale, V.; Natalotto, A.; Giannetto, A.; Barreca, D.; Brunelli, E.; Mauceri, A.; Fasulo, S. Developmental abnormalities and neurotoxicological effects of CuO NPs on the black sea urchin Arbacia lixula by embryo-toxicity assay. Mar. Environ. Res. 2015, 111, 121–127. [Google Scholar] [CrossRef] [PubMed]
- Zhao, C.; Ding, J.; Yang, M.; Shi, D.; Yin, D.; Hu, F.; Sun, J.; Chi, X.; Zhang, L.; Chang, Y. Transcriptomes reveal genes involved in covering and sheltering behaviors of the sea urchin Strongylocentrotus intermedius exposed to UV-B radiation. Ecotoxicol. Environ. Saf. 2019, 167, 236–241. [Google Scholar] [CrossRef] [PubMed]
- Adams, N.L.; Campanale, J.P.; Foltz, K.R. Proteomic Responses of Sea Urchin Embryos to Stressful Ultraviolet Radiation. Integr. Comp. Biol. 2012, 52, 665–680. [Google Scholar] [CrossRef] [Green Version]
- Clapham, D.E. TRP channels as cellular sensors. Nat. Cell Biol. 2003, 426, 517–524. [Google Scholar] [CrossRef]
- Gobron, S.; Creveaux, I.; Meiniel, R.; Didier, R.; Dastugue, B.; Meiniel, A. The evolution of metazoan aα-carbonic anhydrases and their roles in calcium carbonate biomineralization. Neuroscience 1998, 88, 655–664. [Google Scholar] [CrossRef]
- Bonaventura, R.; Zito, F.; Russo, R.; Costa, C. A preliminary gene expression analysis on Paracentrotus lividus embryos exposed to UVB, Cadmium and their combination. Aquat. Toxicol. 2021, 232, 105770. [Google Scholar] [CrossRef]
- Wu, H.; Wang, W.-X. NMR-based metabolomic studies on the toxicological effects of cadmium and copper on green mussels Perna viridis. Aquat. Toxicol. 2010, 100, 339–345. [Google Scholar] [CrossRef]
- Wu, H.; Wang, W.X. Tissue-specific toxicological effects of cadmium in green mussels (Perna viridis): Nuclear magnetic reso-nance-based metabolomics study. Environ. Toxicol. Chem. 2011, 30, 806–812. [Google Scholar] [CrossRef] [PubMed]
- Dumas, T.; Boccard, J.; Gomez, E.; Fenet, H.; Courant, F. Multifactorial analysis of environmental metabolomic data in eco-toxicology: Wild marine mussel exposed to wwtp effluent as a case study. Metabolites 2020, 10, 269. [Google Scholar] [CrossRef] [PubMed]
- Porterfield, S.P.; Hendry, L.B. Impact of Pcbs On Thyroid Hormone Directed Brain Development. Toxicol. Ind. Heal. 1998, 14, 103–120. [Google Scholar] [CrossRef]
- Miller, V.; Kahnke, T.; Neu, N.; Sanchez-Morrissey, S.; Brosch, K.; Kelsey, K.; Seegal, R. Developmental PCB exposure induces hypothyroxinemia and sex-specific effects on cerebellum glial protein levels in rats. Int. J. Dev. Neurosci. 2010, 28, 553–560. [Google Scholar] [CrossRef] [Green Version]
- Buskey, E.J.; Strickler, J.R.; Bradley, C.J.; Hartline, D.K.; Lenz, P.H. Escapes in copepods: Comparison between myelinate and amyelinate species. J. Exp. Biol. 2017, 220, 754–758. [Google Scholar] [CrossRef] [Green Version]


{kind=link}
| Metals | |
|---|---|
| Essential | Non-Essential |
| Copper | Arsenic * |
| Iron | Cadmium |
| Manganese | Lead |
| Zinc | Mercury |
| Nickel | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Deidda, I.; Russo, R.; Bonaventura, R.; Costa, C.; Zito, F.; Lampiasi, N. Neurotoxicity in Marine Invertebrates: An Update. Biology 2021, 10, 161. https://doi.org/10.3390/biology10020161
Deidda I, Russo R, Bonaventura R, Costa C, Zito F, Lampiasi N. Neurotoxicity in Marine Invertebrates: An Update. Biology. 2021; 10(2):161. https://doi.org/10.3390/biology10020161
Chicago/Turabian StyleDeidda, Irene, Roberta Russo, Rosa Bonaventura, Caterina Costa, Francesca Zito, and Nadia Lampiasi. 2021. "Neurotoxicity in Marine Invertebrates: An Update" Biology 10, no. 2: 161. https://doi.org/10.3390/biology10020161
APA StyleDeidda, I., Russo, R., Bonaventura, R., Costa, C., Zito, F., & Lampiasi, N. (2021). Neurotoxicity in Marine Invertebrates: An Update. Biology, 10(2), 161. https://doi.org/10.3390/biology10020161