
Drug Resistance in Nontuberculous Mycobacteria: Mechanisms and Models


Abstract
:Simple Summary
Abstract
1. The Rise of Nontuberculous Mycobacteria
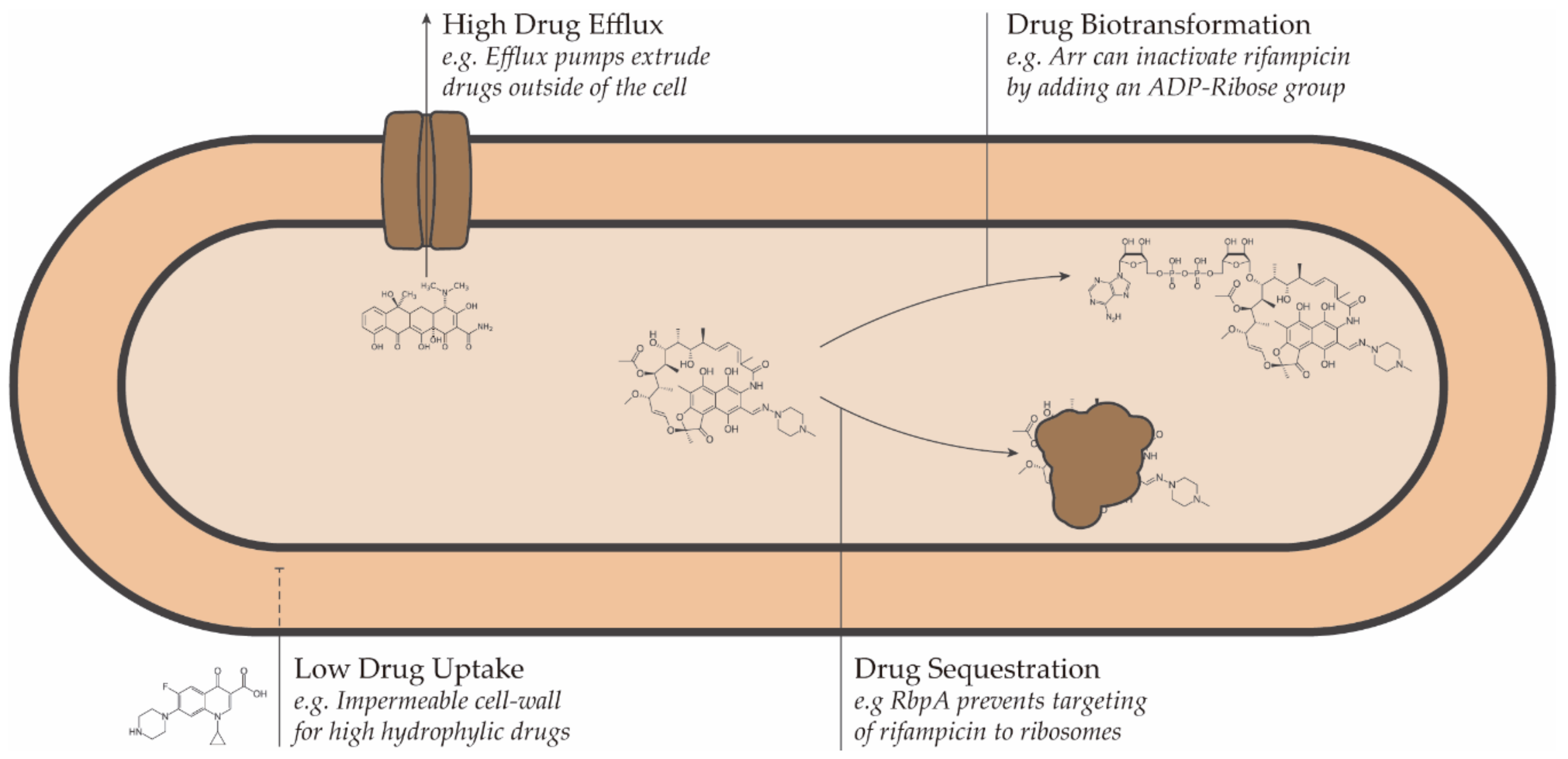
2. Mechanisms of Drug Resistance in Nontuberculous Mycobacteria
2.1. Drug Uptake
2.2. Drug Efflux
2.3. Drug Transformation and Sequestration
3. Models for Drug Discovery against NTM
3.1. In Silico Predictions
3.2. In Vitro Susceptibility Testing
3.3. In Vivo Models
3.4. Iterative Approach to Drug Design
4. Considerations for the Design of Therapies against NTMs
4.1. Optimization of Known Compounds Relevant for Combatting NTM
4.2. Synergies and Combination Therapies
4.3. Host-Directed Therapies
5. Summary and Future Perspectives
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Global Tuberculosis Report 2020. Available online: https://www.who.int/teams/global-tuberculosis-programme/tb-reports (accessed on 23 January 2021).
- Leprosy. Available online: https://www.who.int/news-room/fact-sheets/detail/leprosy (accessed on 23 January 2021).
- Yates, V.M. Mycobacterial Infections. In Rook’s Textbook of Dermatology; Wiley-Blackwell: Oxford, UK, 2010; pp. 1–41. ISBN 9781405161695. [Google Scholar]
- Griffith, D.E.; Aksamit, T.; Brown-Elliott, B.A.; Catanzaro, A.; Daley, C.; Gordin, F.; Holland, S.M.; Horsburgh, R.; Huitt, G.; Iademarco, M.F.; et al. An official ATS/IDSA Statement: Diagnosis, treatment, and prevention of nontuberculous mycobacterial diseases. Am. J. Respir. Crit. Care Med. 2007, 175, 367–416. [Google Scholar] [CrossRef] [PubMed]
- Kendall, B.; Winthrop, K. Update on the epidemiology of pulmonary nontuberculous mycobacterial infections. Semin. Respir. Crit. Care Med. 2013, 34, 087–094. [Google Scholar] [CrossRef] [PubMed]
- Ratnatunga, C.N.; Lutzky, V.P.; Kupz, A.; Doolan, D.L.; Reid, D.W.; Field, M.; Bell, S.C.; Thomson, R.M.; Miles, J.J. The rise of non-tuberculosis mycobacterial lung disease. Front. Immunol. 2020, 11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gopalaswamy, R.; Shanmugam, S.; Mondal, R.; Subbian, S. Of tuberculosis and non-tuberculous mycobacterial infections–a comparative analysis of epidemiology, diagnosis and treatment. J. Biomed. Sci. 2020, 27, 74. [Google Scholar] [CrossRef] [PubMed]
- Runyon, E.H. Anonymous mycobacteria in pulmonary disease. Med. Clin. N. Am. 1959. [Google Scholar] [CrossRef]
- Kim, C.-J.; Kim, N.-H.; Song, K.-H.; Choe, P.G.; Kim, E.S.; Park, S.W.; Kim, H.-B.; Kim, N.-J.; Kim, E.-C.; Park, W.B.; et al. Differentiating rapid- and slow-growing mycobacteria by difference in time to growth detection in liquid media. Diagn. Microbiol. Infect. Dis. 2013, 75, 73–76. [Google Scholar] [CrossRef] [PubMed]
- Porvaznik, I.; Solovič, I.; Mokrý, J. Non-tuberculous mycobacteria: Classification, diagnostics, and therapy. Adv. Exp. Med. Biol. 2017. [Google Scholar] [CrossRef]
- Johnson, M.M.; Odell, J.A. Nontuberculous mycobacterial pulmonary infections. J. Thorac. Dis. 2014, 6, 210. [Google Scholar] [PubMed]
- Nessar, R.; Cambau, E.; Reyrat, J.M.; Murray, A.; Gicquel, B. Mycobacterium abscessus: A new antibiotic nightmare. J. Antimicrob. Chemother. 2012. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Devulder, G.; de Montclos, M.P.; Flandrois, J.P. A multigene approach to phylogenetic analysis using the genus Mycobacterium as a model. Int. J. Syst. Evol. Microbiol. 2005, 55, 293–302. [Google Scholar] [CrossRef] [PubMed]
- Gupta, R.S.; Lo, B.; Son, J. Phylogenomics and comparative genomic studies robustly support division of the genus mycobacterium into an emended genus mycobacterium and four novel genera. Front. Microbiol. 2018, 9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wee, W.Y.; Dutta, A.; Choo, S.W. Comparative genome analyses of mycobacteria give better insights into their evolution. PLoS ONE 2017, 12, e0172831. [Google Scholar] [CrossRef] [PubMed]
- Jagielski, T.; Borówka, P.; Bakuła, Z.; Lach, J.; Marciniak, B.; Brzostek, A.; Dziadek, J.; Dziurzyński, M.; Pennings, L.; van Ingen, J.; et al. Genomic insights into the mycobacterium kansasii complex: An update. Front. Microbiol. 2020, 10, 2918. [Google Scholar] [CrossRef] [Green Version]
- Hamieh, A.; Tayyar, R.; Tabaja, H.; Zein, S.E.L.; Bou Khalil, P.; Kara, N.; Kanafani, Z.A.; Kanj, N.; Bou Akl, I.; Araj, G.; et al. Emergence of Mycobacterium simiae: A retrospective study from a tertiary care center in Lebanon. PLoS ONE 2018, 13, e0195390. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Franco-Paredes, C.; Marcos, L.A.; Henao-Martínez, A.F.; Rodríguez-Morales, A.J.; Villamil-Gómez, W.E.; Gotuzzo, E.; Bonifaz, A. Cutaneous Mycobacterial infections. Clin. Microbiol. Rev. 2018, 32. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Douglas, J.G.; Calder, M.A.; Choo-Kang, Y.F.J.; Leitch, A.G. Mycobacterium gordonae: A new pathogen? Thorax 1986, 41, 152–153. [Google Scholar] [CrossRef] [Green Version]
- Suzuki, S.; Morino, E.; Ishii, M.; Namkoong, H.; Yagi, K.; Asakura, T.; Asami, T.; Fujiwara, H.; Uwamino, Y.; Nishimura, T.; et al. Clinical characteristics of pulmonary Mycobacterium scrofulaceum disease in 2001-2011: A case series and literature review. J. Infect. Chemother. 2016, 22, 611–616. [Google Scholar] [CrossRef]
- Han, X.Y.; Tarrand, J.J.; Infante, R.; Jacobson, K.L.; Truong, M. Clinical significance and epidemiologic analyses of Mycobacterium avium and Mycobacterium intracellulare among patients without AIDS. J. Clin. Microbiol. 2005, 43, 4407–4412. [Google Scholar] [CrossRef] [Green Version]
- Doig, C.; Muckersie, L.; Watt, B.; Forbes, K.J. Molecular epidemiology of Mycobacterium malmoense infections in Scotland. J. Clin. Microbiol. 2002, 40, 1103–1105. [Google Scholar] [CrossRef] [Green Version]
- To, K.; Cao, R.; Yegiazaryan, A.; Owens, J.; Venketaraman, V. General Overview of nontuberculous mycobacteria opportunistic pathogens: Mycobacterium avium and Mycobacterium abscessus. J. Clin. Med. 2020, 9. [Google Scholar] [CrossRef]
- Jones, R.S.; Shier, K.L.; Master, R.N.; Bao, J.R.; Clark, R.B. Current significance of the Mycobacterium chelonae-abscessus group. Diagn. Microbiol. Infect. Dis. 2019, 94, 248–254. [Google Scholar] [CrossRef]
- Máiz Carro, L.; Barbero Herranz, E.; Nieto Royo, R. Respiratory infections due to nontuberculous mycobacterias. Med. Clin. 2018, 150, 191–197. [Google Scholar] [CrossRef] [PubMed]
- Nishiuchi, Y.; Iwamoto, T.; Maruyama, F. Infection sources of a common non-tuberculous mycobacterial pathogen, Mycobacterium avium complex. Front. Med. 2017, 4, 27. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shah, N.M.; Davidson, J.A.; Anderson, L.F.; Lalor, M.K.; Kim, J.; Thomas, H.L.; Lipman, M.; Abubakar, I. Pulmonary Mycobacterium avium-intracellulare is the main driver of the rise in non-tuberculous mycobacteria incidence in England, Wales and Northern Ireland, 2007–2012. BMC Infect. Dis. 2016. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Johansen, M.D.; Herrmann, J.L.; Kremer, L. Non-tuberculous mycobacteria and the rise of Mycobacterium abscessus. Nat. Rev. Microbiol. 2020, 18, 392–407. [Google Scholar] [CrossRef] [PubMed]
- Fleshner, M.; Olivier, K.N.; Shaw, P.A.; Adjemian, J.; Strollo, S.; Claypool, R.J.; Folio, L.; Zelazny, A.; Holland, S.M.; Prevots, D.R.; et al. Mortality among patients with pulmonary non-tuberculous mycobacteria disease. Int. J. Tuberc. Lung Dis. 2016. [Google Scholar] [CrossRef] [PubMed]
- Jarand, J.; Levin, A.; Zhang, L.; Huitt, G.; Mitchell, J.D.; Daley, C.L. Clinical and microbiologic outcomes in patients receiving treatment for Mycobacterium abscessus pulmonary disease. Clin. Infect. Dis. 2011. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Taylor, J.L.; Palmer, S.M. Mycobacterium abscessus chest wall and pulmonary infection in a cystic fibrosis lung transplant recipient. J. Hear. Lung Transplant. 2006. [Google Scholar] [CrossRef]
- Bryant, J.M.; Grogono, D.M.; Rodriguez-Rincon, D.; Everall, I.; Brown, K.P.; Moreno, P.; Verma, D.; Hill, E.; Drijkoningen, J.; Gilligan, P.; et al. Emergence and spread of a humantransmissible multidrug-resistant nontuberculous mycobacterium. Science 2016. [Google Scholar] [CrossRef] [Green Version]
- Hermansen, T.S.; Ravn, P.; Svensson, E.; Lillebaek, T. Nontuberculous mycobacteria in Denmark, incidence and clinical importance during the last quarter-century. Sci. Rep. 2017. [Google Scholar] [CrossRef] [Green Version]
- Baldwin, S.L.; Larsenid, S.E.; Ordway, D.; Cassell, G.; Coler, R.N. The complexities and challenges of preventing and treating nontuberculous mycobacterial diseases. PLoS Negl. Trop. Dis. 2019, 13, e0007083. [Google Scholar] [CrossRef] [PubMed]
- Faverio, P.; Stainer, A.; Bonaiti, G.; Zucchetti, S.C.; Simonetta, E.; Lapadula, G.; Marruchella, A.; Gori, A.; Blasi, F.; Codecasa, L.; et al. Characterizing non-tuberculous mycobacteria infection in bronchiectasis. Int. J. Mol. Sci. 2016. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Santin, M.; Dorca, J.; Alcaide, F.; Gonzalez, L.; Casas, S.; Lopez, M.; Guerra, M.R. Long-term relapses after 12-month treatment for Mycobacterium kansasii lung disease. Eur. Respir. J. 2009. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sim, Y.S.; Park, H.Y.; Jeon, K.; Suh, G.Y.; Kwon, O.J.; Koh, W.-J. Standardized combination antibiotic treatment of Mycobacterium avium complex lung disease. Yonsei Med. J. 2010, 51, 888. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wallace, R.J.; Dukart, G.; Brown-Elliott, B.A.; Griffith, D.E.; Scerpella, E.G.; Marshall, B. Clinical experience in 52 patients with tigecycline-containing regimens for salvage treatment of Mycobacterium abscessus and Mycobacterium chelonae infections. J. Antimicrob. Chemother. 2014, 69, 1945–1953. [Google Scholar] [CrossRef] [PubMed]
- Diagnosis and treatment of disease caused by nontuberculous mycobacteria. This official statement of the American Thoracic Society was approved by the Board of Directors, March 1997. Medical Section of the American Lung Association. Am. J. Respir. Crit. Care Med. 1997, 156, S1–S25. [CrossRef]
- Mirsaeidi, M.; Farshidpour, M.; Allen, M.B.; Ebrahimi, G.; Falkinham, J.O. Highlight on advances in nontuberculous mycobacterial disease in North America. Biomed Res. Int. 2014, 2014, 919474. [Google Scholar] [CrossRef] [PubMed]
- Wu, M.-L.; Aziz, D.B.; Dartois, V.; Dick, T. NTM drug discovery: Status, gaps and the way forward. Drug Discov. Today 2018, 23, 1502–1519. [Google Scholar] [CrossRef]
- Munita, J.M.; Arias, C.A.; Unit, A.R.; Santiago, A. De HHS Public access mechanisms of antibiotic resistance. HHS Public Access 2016, 4, 1–37. [Google Scholar] [CrossRef]
- Moon, S.M.; Park, H.Y.; Kim, S.-Y.; Jhun, B.W.; Lee, H.; Jeon, K.; Kim, D.H.; Huh, H.J.; Ki, C.-S.; Lee, N.Y.; et al. Clinical characteristics, treatment outcomes, and resistance mutations associated with macrolide-resistant Mycobacterium avium complex lung disease. Antimicrob. Agents Chemother. 2016, 60, 6758–6765. [Google Scholar] [CrossRef] [Green Version]
- Nasiri, M.J.; Haeili, M.; Ghazi, M.; Goudarzi, H.; Pormohammad, A.; Imani Fooladi, A.A.; Feizabadi, M.M. New insights in to the intrinsic and acquired drug resistance mechanisms in mycobacteria. Front. Microbiol. 2017, 8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hayes, J.D.; Wolf, C.R. Molecular mechanisms of drug resistance. Biochem. J. 1990, 272, 281–295. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jarlier, V.; Nikaido, H. Mycobacterial cell wall: Structure and role in natural resistance to antibiotics. FEMS Microbiol. Lett. 1994, 123, 11–18. [Google Scholar] [CrossRef] [PubMed]
- Falkinham, J.O. Growth in catheter biofilms and antibiotic resistance of Mycobacterium avium. J. Med. Microbiol. 2007, 56, 250–254. [Google Scholar] [CrossRef] [PubMed]
- Falkinham, J.O. Challenges of NTM drug development. Front. Microbiol. 2018, 9. [Google Scholar] [CrossRef]
- Helguera-Repetto, A.C.; Chacon-Salinas, R.; Cerna-Cortes, J.F.; Rivera-Gutierrez, S.; Ortiz-Navarrete, V.; Estrada-Garcia, I.; Gonzalez-y-Merchand, J.A. Differential macrophage response to slow- and fast-growing pathogenic mycobacteria. Biomed. Res. Int. 2014, 2014, 916521. [Google Scholar] [CrossRef] [Green Version]
- Ren, H.; Liu, J. AsnB is involved in natural resistance of Mycobacterium smegmatis to multiple drugs. Antimicrob. Agents Chemother. 2006, 50, 250–255. [Google Scholar] [CrossRef] [Green Version]
- Wolff, K.A.; Nguyen, H.T.; Cartabuke, R.H.; Singh, A.; Ogwang, S.; Nguyen, L. Protein Kinase G Is required for intrinsic antibiotic resistance in mycobacteria. Antimicrob. Agents Chemother. 2009, 53, 3515–3519. [Google Scholar] [CrossRef] [Green Version]
- Cangelosi, G.A.; Palermo, C.O.; Laurent, J.-P.; Hamlin, A.M.; Brabant, W.H. Colony morphotypes on Congo red agar segregate along species and drug susceptibility lines in the Mycobacterium avium-intracellulare complex. Microbiology 1999, 145, 1317–1324. [Google Scholar] [CrossRef] [Green Version]
- Cangelosi, G.A.; Do, J.S.; Freeman, R.; Bennett, J.G.; Semret, M.; Behr, M.A. The two-component regulatory system mtrAB is required for morphotypic multidrug resistance in Mycobacterium avium. Antimicrob. Agents Chemother. 2006, 50, 461–468. [Google Scholar] [CrossRef] [Green Version]
- Philalay, J.S.; Palermo, C.O.; Hauge, K.A.; Rustad, T.R.; Cangelosi, G.A. Genes required for intrinsic multidrug resistance in Mycobacterium avium. Antimicrob. Agents Chemother. 2004, 48, 3412–3418. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smith, T.; Wolff, K.A.; Nguyen, L. Molecular biology of drug resistance in Mycobacterium tuberculosis. Curr. Top. Microbiol. Immunol. 2013, 374, 53–80. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nguyen, L.; Chinnapapagari, S.; Thompson, C.J. FbpA-dependent biosynthesis of trehalose dimycolate is required for the intrinsic multidrug resistance, cell wall structure, and colonial morphology of Mycobacterium smegmatis. J. Bacteriol. 2005, 187, 6603–6611. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Niederweis, M. Nutrient acquisition by mycobacteria. Microbiology 2008, 154, 679–692. [Google Scholar] [CrossRef] [Green Version]
- Sharbati, S.; Schramm, K.; Rempel, S.; Wang, H.; Andrich, R.; Tykiel, V.; Kunisch, R.; Lewin, A. Characterisation of porin genes from Mycobacterium fortuitum and their impact on growth. BMC Microbiol. 2009, 9, 31. [Google Scholar] [CrossRef] [Green Version]
- Lambert, P.A. Cellular impermeability and uptake of biocides and antibiotics in Gram-positive bacteria and mycobacteria. J. Appl. Microbiol. 2002, 92, 46S–54S. [Google Scholar] [CrossRef]
- Sharbati-Tehrani, S.; Stephan, J.; Holland, G.; Appel, B.; Niederweis, M.; Lewin, A. Porins limit the intracellular persistence of Mycobacterium smegmatis. Microbiology 2005, 151, 2403–2410. [Google Scholar] [CrossRef] [Green Version]
- Stephan, J.; Mailaender, C.; Etienne, G.; Daffeé, M.; Niederweis, M. Multidrug resistance of a porin deletion mutant of Mycobacterium smegmatis. Antimicrob. Agents Chemother. 2004, 48, 4163–4170. [Google Scholar] [CrossRef] [Green Version]
- Danilchanka, O.; Pavlenok, M.; Niederweis, M. Role of porins for uptake of antibiotics by Mycobacterium smegmatis. Antimicrob. Agents Chemother. 2008, 52, 3127–3134. [Google Scholar] [CrossRef] [Green Version]
- Falkinham, J.O. Nontuberculous mycobacteria from household plumbing of patients with nontuberculous mycobacteria disease. Emerg. Infect. Dis. 2011, 17, 419–424. [Google Scholar] [CrossRef]
- Van der Wielen, P.W.J.J.; van der Kooij, D. Nontuberculous mycobacteria, fungi, and opportunistic pathogens in unchlorinated drinking water in The Netherlands. Appl. Environ. Microbiol. 2013, 79, 825–834. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kostakioti, M.; Hadjifrangiskou, M.; Hultgren, S.J. Bacterial biofilms: Development, dispersal, and therapeutic strategies in the dawn of the postantibiotic era. Cold Spring Harb. Perspect. Med. 2013, 3, a010306. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yamazaki, Y.; Danelishvili, L.; Wu, M.; Hidaka, E.; Katsuyama, T.; Stang, B.; Petrofsky, M.; Bildfell, R.; Bermudez, L.E. The ability to form biofilm influences Mycobacterium avium invasion and translocation of bronchial epithelial cells. Cell. Microbiol. 2006, 8, 806–814. [Google Scholar] [CrossRef] [PubMed]
- Carter, G.; Wu, M.; Drummond, D.C.; Bermudez, L.E. Characterization of biofilm formation by clinical isolates of Mycobacterium avium. J. Med. Microbiol. 2003, 52, 747–752. [Google Scholar] [CrossRef]
- Brodlie, M.; Aseeri, A.; Lordan, J.L.; Robertson, A.G.N.; McKean, M.C.; Corris, P.A.; Griffin, S.M.; Manning, N.J.; Pearson, J.P.; Ward, C. Bile acid aspiration in people with cystic fibrosis before and after lung transplantation. Eur. Respir. J. 2015, 46, 1820–1823. [Google Scholar] [CrossRef] [Green Version]
- Simoes, M. Antimicrobial strategies effective against infectious bacterial biofilms. Curr. Med. Chem. 2011, 18, 2129–2145. [Google Scholar] [CrossRef]
- Faria, S.; Joao, I.; Jordao, L. General overview on nontuberculous mycobacteria, biofilms, and human Infection. J. Pathog. 2015, 2015, 809014. [Google Scholar] [CrossRef] [Green Version]
- Mah, T.-F. Biofilm-specific antibiotic resistance. Future Microbiol. 2012, 7, 1061–1072. [Google Scholar] [CrossRef] [Green Version]
- Casadevall, A.; Pirofski, L. Virulence factors and their mechanisms of action: The view from a damage–response framework. J. Water Health 2009, 7, S2–S18. [Google Scholar] [CrossRef]
- Steed, K.A.; Falkinham, J.O. Effect of growth in biofilms on chlorine susceptibility of Mycobacterium avium and Mycobacterium intracellulare. Appl. Environ. Microbiol. 2006, 72, 4007–4011. [Google Scholar] [CrossRef] [Green Version]
- Ehlers, S.; Schaible, U.E. The granuloma in tuberculosis: Dynamics of a host–pathogen collusion. Front. Immunol. 2013, 3. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gonzalez-Juarrero, M.; Turner, O.C.; Turner, J.; Marietta, P.; Brooks, J.V.; Orme, I.M. Temporal and spatial arrangement of lymphocytes within lung granulomas induced by aerosol infection with Mycobacterium tuberculosis. Infect. Immun. 2001, 69, 1722–1728. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Puissegur, M.-P.; Botanch, C.; Duteyrat, J.-L.; Delsol, G.; Caratero, C.; Altare, F. An in vitro dual model of mycobacterial granulomas to investigate the molecular interactions between mycobacteria and human host cells. Cell. Microbiol. 2004, 6, 423–433. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Lin, W.M.; Li, H.; Dai, X.D.; Ma, S.P.; Ren, W.H.; Jeon, S.K.; Lee, J.M. Hepatic nontuberculous mycobacterial granulomas in patients with cancer mimicking metastases: An analysis of three cases. Quant. Imaging Med. Surg. 2019, 9, 1126–1131. [Google Scholar] [CrossRef]
- Ufimtseva, E. Mycobacterium -host cell relationships in granulomatous lesions in a mouse model of latent tuberculous infection. Biomed Res. Int. 2015, 2015, 948131. [Google Scholar] [CrossRef] [Green Version]
- Dutta, N.K.; Karakousis, P.C. Latent tuberculosis infection: Myths, models, and molecular mechanisms. Microbiol. Mol. Biol. Rev. 2014, 78, 343–371. [Google Scholar] [CrossRef] [Green Version]
- Machado, D.; Lecorche, E.; Mougari, F.; Cambau, E.; Viveiros, M. Insights on Mycobacterium leprae Efflux Pumps and their implications in drug resistance and virulence. Front. Microbiol. 2018, 9. [Google Scholar] [CrossRef]
- Li, X.-Z.; Zhang, L.; Nikaido, H. Efflux pump-mediated intrinsic drug resistance in Mycobacterium smegmatis. Antimicrob. Agents Chemother. 2004, 48, 2415–2423. [Google Scholar] [CrossRef] [Green Version]
- Liu, J.; Takiff, H.E.; Nikaido, H. Active efflux of fluoroquinolones in Mycobacterium smegmatis mediated by LfrA, a multidrug efflux pump. J. Bacteriol. 1996, 178, 3791–3795. [Google Scholar] [CrossRef] [Green Version]
- De Rossi, E.; Blokpoel, M.C.J.; Cantoni, R.; Branzoni, M.; Riccardi, G.; Young, D.B.; De Smet, K.A.L.; Ciferri, O. Molecular cloning and functional analysis of a novel tetracycline resistance determinant, tet(V), from Mycobacterium smegmatis. Antimicrob. Agents Chemother. 1998, 42, 1931–1937. [Google Scholar] [CrossRef] [Green Version]
- Silva, P.E.A.; Bigi, F.; de la Paz Santangelo, M.; Romano, M.I.; Martín, C.; Cataldi, A.; Aínsa, J.A. Characterization of P55, a multidrug efflux pump in Mycobacterium bovis and Mycobacterium tuberculosis. Antimicrob. Agents Chemother. 2001, 45, 800–804. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kwon, H.H.; Tomioka, H.; Saito, H. Distribution and characterization of β-lactamases of mycobacteria and related organisms. Tuber. Lung Dis. 1995, 76, 141–148. [Google Scholar] [CrossRef]
- Rominski, A.; Schulthess, B.; Müller, D.M.; Keller, P.M.; Sander, P. Effect of β-lactamase production and β-lactam instability on MIC testing results for Mycobacterium abscessus. J. Antimicrob. Chemother. 2017, 72, 3070–3078. [Google Scholar] [CrossRef] [PubMed]
- Lavollay, M.; Dubée, V.; Heym, B.; Herrmann, J.-L.; Gaillard, J.-L.; Gutmann, L.; Arthur, M.; Mainardi, J.-L. In vitro activity of cefoxitin and imipenem against Mycobacterium abscessus complex. Clin. Microbiol. Infect. 2014, 20, O297–O300. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pandey, R.; Chen, L.; Manca, C.; Jenkins, S.; Glaser, L.; Vinnard, C.; Stone, G.; Lee, J.; Mathema, B.; Nuermberger, E.L.; et al. Dual β-lactam combinations highly active against Mycobacterium abscessus complex in vitro. MBio 2019, 10. [Google Scholar] [CrossRef] [Green Version]
- Lefebvre, A.-L.; Dubée, V.; Cortes, M.; Dorchêne, D.; Arthur, M.; Mainardi, J.-L. Bactericidal and intracellular activity of β-lactams against Mycobacterium abscessus. J. Antimicrob. Chemother. 2016, 71, 1556–1563. [Google Scholar] [CrossRef] [Green Version]
- Adjei, M.D.; Heinze, T.M.; Deck, J.; Freeman, J.P.; Williams, A.J.; Sutherland, J.B. Acetylation and nitrosation of ciprofloxacin by environmental strains of mycobacteria. Can. J. Microbiol. 2007, 53, 144–147. [Google Scholar] [CrossRef]
- Payton, M.; Auty, R.; Delgoda, R.; Everett, M.; Sim, E. Cloning and characterization of arylamine N -acetyltransferase genes from Mycobacterium smegmatis and Mycobacterium tuberculosis: Increased expression results in isoniazid resistance. J. Bacteriol. 1999, 181, 1343–1347. [Google Scholar] [CrossRef] [Green Version]
- Aínsa, J.A.; Pérez, E.; Pelicic, V.; Berthet, F.; Gicquel, B.; Martín, C. Aminoglycoside 2′- N -acetyltransferase genes are universally present in mycobacteria: Characterization of the aac(2′)-Ic gene from Mycobacterium tuberculosis and the aac(2 ′ )-Id gene from Mycobacterium smegmatis. Mol. Microbiol. 1997, 24, 431–441. [Google Scholar] [CrossRef]
- Ripoll, F.; Pasek, S.; Schenowitz, C.; Dossat, C.; Barbe, V.; Rottman, M.; Macheras, E.; Heym, B.; Herrmann, J.-L.; Daffé, M.; et al. Non mycobacterial virulence genes in the genome of the emerging pathogen Mycobacterium abscessus. PLoS ONE 2009, 4, e5660. [Google Scholar] [CrossRef] [Green Version]
- Rominski, A.; Roditscheff, A.; Selchow, P.; Böttger, E.C.; Sander, P. Intrinsic rifamycin resistance of Mycobacterium abscessus is mediated by ADP-ribosyltransferase MAB_0591. J. Antimicrob. Chemother. 2017, 72, 376–384. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baysarowich, J.; Koteva, K.; Hughes, D.W.; Ejim, L.; Griffiths, E.; Zhang, K.; Junop, M.; Wright, G.D. Rifamycin antibiotic resistance by ADP-ribosylation: Structure and diversity of Arr. Proc. Natl. Acad. Sci. USA 2008, 105, 4886–4891. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blair, J.M.A.; Webber, M.A.; Baylay, A.J.; Ogbolu, D.O.; Piddock, L.J.V. Molecular mechanisms of antibiotic resistance. Nat. Rev. Microbiol. 2015, 13, 42–51. [Google Scholar] [CrossRef]
- Adizie, J.; Qasim, M.; Pagaria, M. S39 Risk of NTM (non tuberculous mycobacterium) infection in patients on long term prophylactic macrolide antibiotics. Thorax 2016, 71, A24–A25. [Google Scholar] [CrossRef] [Green Version]
- Meier, A.; Kirschner, P.; Springer, B.; Steingrube, V.A.; Brown, B.A.; Wallace, R.J.; Böttger, E.C. Identification of mutations in 23S rRNA gene of clarithromycin-resistant Mycobacterium intracellulare. Antimicrob. Agents Chemother. 1994, 38, 381–384. [Google Scholar] [CrossRef] [Green Version]
- Bastian, S.; Veziris, N.; Roux, A.L.; Brossier, F.; Gaillard, J.L.; Jarlier, V.; Cambau, E. Assessment of clarithromycin susceptibility in strains belonging to the Mycobacterium abscessus group by erm(41) and rrl sequencing. Antimicrob. Agents Chemother. 2011, 55, 775–781. [Google Scholar] [CrossRef] [Green Version]
- Sander, P.; Prammananan, T.; Meier, A.; Frischkorn, K.; Böttger, E.C. The role of ribosomal RNAs in macrolide resistance. Mol. Microbiol. 1997, 26, 469–480. [Google Scholar] [CrossRef]
- Brown-Elliott, B.A.; Nash, K.A.; Wallace, R.J. Antimicrobial susceptibility testing, drug resistance mechanisms, and therapy of infections with nontuberculous mycobacteria. Clin. Microbiol. Rev. 2012, 25, 545–582. [Google Scholar] [CrossRef] [Green Version]
- Obata, S.; Zwolska, Z.; Toyota, E.; Kudo, K.; Nakamura, A.; Sawai, T.; Kuratsuji, T.; Kirikae, T. Association of rpoB mutations with rifampicin resistance in Mycobacterium avium. Int. J. Antimicrob. Agents 2006, 27, 32–39. [Google Scholar] [CrossRef]
- Klein, J.L.; Brown, T.J.; French, G.L. Rifampin resistance in Mycobacterium kansasii is associated with rpoB mutations. Antimicrob. Agents Chemother. 2001, 45, 3056–3058. [Google Scholar] [CrossRef] [Green Version]
- Zhu, J.-H.; Wang, B.-W.; Pan, M.; Zeng, Y.-N.; Rego, H.; Javid, B. Rifampicin can induce antibiotic tolerance in mycobacteria via paradoxical changes in rpoB transcription. Nat. Commun. 2018, 9, 4218. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dey, A.; Verma, A.K.; Chatterji, D. Role of an RNA polymerase interacting protein, MsRbpA, from Mycobacterium smegmatis in phenotypic tolerance to rifampicin. Microbiology 2010, 156, 873–883. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rampacci, E.; Stefanetti, V.; Passamonti, F.; Henao-Tamayo, M. Preclinical models of nontuberculous mycobacteria infection for early drug discovery and vaccine research. Pathogens 2020, 9, 641. [Google Scholar] [CrossRef] [PubMed]
- Iskar, M.; Zeller, G.; Zhao, X.-M.; van Noort, V.; Bork, P. Drug discovery in the age of systems biology: The rise of computational approaches for data integration. Curr. Opin. Biotechnol. 2012, 23, 609–616. [Google Scholar] [CrossRef]
- Khurshid Ahmad, M.H. Drug discovery and in silico techniques: A mini-review. Enzym. Eng. 2014, 4. [Google Scholar] [CrossRef]
- Amberg, A. In Silico Methods. In Drug Discovery and Evaluation: Safety and Pharmacokinetic Assays; Springer: Berlin/Heidelberg, Germany, 2013; pp. 1273–1296. ISBN 9783642252402. [Google Scholar]
- Zloh, M.; Kirton, S.B. The benefits of in silico modeling to identify possible small-molecule drugs and their off-target interactions. Future Med. Chem. 2018, 10, 423–432. [Google Scholar] [CrossRef] [Green Version]
- Truman, R.W.; Ebenezer, G.J.; Pena, M.T.; Sharma, R.; Balamayooran, G.; Gillingwater, T.H.; Scollard, D.M.; McArthur, J.C.; Rambukkana, A. The armadillo as a model for peripheral neuropathy in leprosy. ILAR J. 2014, 54, 304–314. [Google Scholar] [CrossRef] [Green Version]
- Nisha, J.; Shanthi, V. Computational simulation techniques to understand rifampicin resistance mutation (S425L) of rpoB in M. leprae. J. Cell. Biochem. 2015, 116, 1278–1285. [Google Scholar] [CrossRef]
- Vedithi, S.C.; Lavania, M.; Kumar, M.; Kaur, P.; Turankar, R.P.; Singh, I.; Nigam, A.; Sengupta, U. A report of rifampin-resistant leprosy from northern and eastern India: Identification and in silico analysis of molecular interactions. Med. Microbiol. Immunol. 2015, 204, 193–203. [Google Scholar] [CrossRef]
- Nisha, J.; Shanthi, V. Characterization of ofloxacin interaction with mutated (A91V) Quinolone resistance determining region of DNA gyrase in Mycobacterium Leprae through computational simulation. Cell Biochem. Biophys. 2018, 76, 125–134. [Google Scholar] [CrossRef]
- Macalino, S.J.Y.; Billones, J.B.; Organo, V.G.; Carrillo, M.C.O. In Silico strategies in tuberculosis drug discovery. Molecules 2020, 25, 665. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sugiki, T.; Furuita, K.; Fujiwara, T.; Kojima, C. Current NMR techniques for structure-based drug discovery. Molecules 2018, 23, 148. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Batool, M.; Ahmad, B.; Choi, S. A structure-based drug discovery paradigm. Int. J. Mol. Sci. 2019, 20, 2783. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rehna, E.A.A.; Singh, S.K.; Dharmalingam, K. Functional insights by comparison of modeled structures of 18kDa small heat shock protein and its mutant in Mycobacterium leprae. Bioinformation 2008, 3, 230–234. [Google Scholar] [CrossRef] [Green Version]
- Ramis, I.B.; Vianna, J.S.; Silva Junior, L.; von Groll, A.; Ramos, D.F.; Lobo, M.M.; Zanatta, N.; Viveiros, M.; da Silva, P.E.A. In silico and in vitro evaluation of tetrahydropyridine compounds as efflux inhibitors in Mycobacterium abscessus. Tuberculosis 2019, 118, 101853. [Google Scholar] [CrossRef]
- Sotriffer, C.A. Molecular dynamics simulations in drug design. In Encyclopedic Reference of Genomics and Proteomics in Molecular Medicine; Springer: Berlin/Heidelberg, Germany, 2006; pp. 1153–1160. [Google Scholar]
- Kumar, A.; Sechi, L.A.; Caboni, P.; Marrosu, M.G.; Atzori, L.; Pieroni, E. Dynamical insights into the differential characteristics of Mycobacterium avium subsp. paratuberculosis peptide binding to HLA-DRB1 proteins associated with multiple sclerosis. New J. Chem. 2015, 39, 1355–1366. [Google Scholar] [CrossRef]
- Ferreira, L.L.G.; Andricopulo, A.D. Editorial: Chemoinformatics approaches to structure- and ligand-based drug design. Front. Pharmacol. 2018, 9. [Google Scholar] [CrossRef]
- Ruyck, J.; Dupont, C.; Lamy, E.; Le Moigne, V.; Biot, C.; Guérardel, Y.; Herrmann, J.; Blaise, M.; Grassin-Delyle, S.; Kremer, L.; et al. Structure-based design and synthesis of piperidinol-containing molecules as new Mycobacterium abscessus inhibitors. ChemistryOpen 2020, 9, 351–365. [Google Scholar] [CrossRef]
- Dupont, C.; Viljoen, A.; Dubar, F.; Blaise, M.; Bernut, A.; Pawlik, A.; Bouchier, C.; Brosch, R.; Guérardel, Y.; Lelièvre, J.; et al. A new piperidinol derivative targeting mycolic acid transport in Mycobacterium abscessus. Mol. Microbiol. 2016, 101, 515–529. [Google Scholar] [CrossRef] [Green Version]
- Bakour, S.; Sankar, S.A.; Rathored, J.; Biagini, P.; Raoult, D.; Fournier, P.-E. Identification of virulence factors and antibiotic resistance markers using bacterial genomics. Future Microbiol. 2016, 11, 455–466. [Google Scholar] [CrossRef]
- Le Moigne, V.; Belon, C.; Goulard, C.; Accard, G.; Bernut, A.; Pitard, B.; Gaillard, J.-L.; Kremer, L.; Herrmann, J.-L.; Blanc-Potard, A.-B. MgtC as a Host-induced factor and vaccine candidate against Mycobacterium abscessus infection. Infect. Immun. 2016, 84, 2895–2903. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Soni, I.; De Groote, M.A.; Dasgupta, A.; Chopra, S. Challenges facing the drug discovery pipeline for non-tuberculous mycobacteria. J. Med. Microbiol. 2016, 65, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Quan, T.P.; Bawa, Z.; Foster, D.; Walker, T.; Del Ojo Elias, C.; Rathod, P.; MMM Informatics Group; Iqbal, Z.; Bradley, P.; Mowbray, J.; et al. Evaluation of whole-genome sequencing for mycobacterial species identification and drug susceptibility testing in a clinical setting: A large-scale prospective assessment of performance against line probe assays and phenotyping. J. Clin. Microbiol. 2018, 56. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matsumoto, Y.; Kinjo, T.; Motooka, D.; Nabeya, D.; Jung, N.; Uechi, K.; Horii, T.; Iida, T.; Fujita, J.; Nakamura, S. Comprehensive subspecies identification of 175 nontuberculous mycobacteria species based on 7547 genomic profiles. Emerg. Microbes Infect. 2019, 8, 1043–1053. [Google Scholar] [CrossRef]
- National Research Council. Summary of Advantages and Disadvantages of in vitro and in vivo methods. In Monoclonal Antibody Production; National Academy Press: Washington, DC, USA, 2000; ISBN 978-0309075114. [Google Scholar]
- Jonkman, J.H.; van Bork, L.E.; Wijsbeek, J.; de Zeeuw, R.A.; Orie, N.G.; Cox, H.L. “First pass effect” after rectal administration of thiazinamium methylsulphate [proceedings]. J. Pharm. Pharmacol. 1976, 28, 56. [Google Scholar] [CrossRef]
- Bose, M.; Venugopal, D.; Kumar, S.; Isa, M. Drug resistance profile of human Mycobacterium avium complex strains from India. Indian J. Med. Microbiol. 2007, 25, 115. [Google Scholar] [CrossRef] [PubMed]
- Wallace, R.J.; Dalovisio, J.R.; Pankey, G.A. Disk diffusion testing of susceptibility of Mycobacterium fortuitum and Mycobacterium chelonei to antibacterial agents. Antimicrob. Agents Chemother. 1979, 16, 611–614. [Google Scholar] [CrossRef] [Green Version]
- Nair, D.; Verma, J.; Rawat, D.; Hasan, A.; Capoor, M.; Gupta, K.; Deb, M.; Aggarwal, P. The use of E-test for the drug susceptibility testing of Mycobacterium tuberculosis-A solution or an illusion? Indian J. Med. Microbiol. 2010, 28, 30. [Google Scholar] [CrossRef]
- Freixo, I.M.; Caldas, P.C.S.; Martins, F.; Brito, R.C.; Ferreira, R.M.C.; Fonseca, L.S.; Saad, M.H.F. Evaluation of etest strips for rapid susceptibility testing of Mycobacterium tuberculosis. J. Clin. Microbiol. 2002, 40, 2282–2284. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Falkinham, J.O.; Macri, R.V.; Maisuria, B.B.; Actis, M.L.; Sugandhi, E.W.; Williams, A.A.; Snyder, A.V.; Jackson, F.R.; Poppe, M.A.; Chen, L.; et al. Antibacterial activities of dendritic amphiphiles against nontuberculous mycobacteria. Tuberculosis 2012, 92, 173–181. [Google Scholar] [CrossRef]
- Khan, Z.A.; Siddiqui, M.F.; Park, S. Current and emerging methods of antibiotic susceptibility testing. Diagnostics 2019, 9, 49. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Malhotra, S.; Vedithi, S.C.; Blundell, T.L. Decoding the similarities and differences among mycobacterial species. PLoS Negl. Trop. Dis. 2017, 11, e0005883. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Andries, K.; Verhasselt, P.; Guillemont, J.; Göhlmann, H.W.H.; Neefs, J.-M.; Winkler, H.; Van Gestel, J.; Timmerman, P.; Zhu, M.; Lee, E.; et al. A diarylquinoline drug active on the ATP synthase of Mycobacterium tuberculosis. Science 2005, 307, 223–227. [Google Scholar] [CrossRef] [PubMed]
- Von Reyn, C.F.; Jacobs, N.J.; Arbeit, R.D.; Maslow, J.N.; Niemczyk, S. Polyclonal Mycobacterium avium infections in patients with AIDS: Variations in antimicrobial susceptibilities of different strains of M. avium isolated from the same patient. J. Clin. Microbiol. 1995, 33, 1008–1010. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Wijk, R.C.; Sar, A.M.; Krekels, E.H.J.; Verboom, T.; Spaink, H.P.; Simonsson, U.S.H.; Graaf, P.H. Quantification of natural growth of two strains of Mycobacterium Marinum for translational antituberculosis drug development. Clin. Transl. Sci. 2020, 13, 1060–1064. [Google Scholar] [CrossRef] [Green Version]
- Falkinham, J.O. Factors influencing the chlorine susceptibility of Mycobacterium avium, Mycobacterium intracellulare, and Mycobacterium scrofulaceum. Appl. Environ. Microbiol. 2003, 69, 5685–5689. [Google Scholar] [CrossRef] [Green Version]
- McCarthy, C. Spontaneous and induced mutation in Mycobacterium avium. Infect. Immun. 1970, 2, 223–228. [Google Scholar] [CrossRef] [Green Version]
- Van Ingen, J.; Boeree, M.J.; van Soolingen, D.; Mouton, J.W. Resistance mechanisms and drug susceptibility testing of nontuberculous mycobacteria. Drug Resist. Updat. 2012, 15, 149–161. [Google Scholar] [CrossRef]
- Nie, W.J.; Xie, Z.Y.; Gao, S.; Teng, T.L.; Zhou, W.Q.; Shang, Y.Y.; Jing, W.; Shi, W.H.; Wang, Q.F.; Huang, X.R.; et al. Efficacy of moxifloxacin against Mycobacterium abscessus in zebrafish model in vivo. Biomed. Environ. Sci. 2020, 33, 350–358. [Google Scholar] [CrossRef]
- Chan, E.D.; Bai, X. Animal models of non-tuberculous mycobacterial infections. Mycobact. Dis. 2016, 6. [Google Scholar] [CrossRef] [Green Version]
- Bernut, A.; Herrmann, J.-L.; Ordway, D.; Kremer, L. The diverse cellular and animal models to decipher the physiopathological traits of Mycobacterium abscessus infection. Front. Cell. Infect. Microbiol. 2017, 7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Groote, M.A.; Johnson, L.; Podell, B.; Brooks, E.; Basaraba, R.; Gonzalez-Juarrero, M. GM-CSF knockout mice for preclinical testing of agents with antimicrobial activity against Mycobacterium abscessus. J. Antimicrob. Chemother. 2014, 69, 1057–1064. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maggioncalda, E.C.; Story-Roller, E.; Mylius, J.; Illei, P.; Basaraba, R.J.; Lamichhane, G. A mouse model of pulmonary Mycobacteroides abscessus infection. Sci. Rep. 2020, 10, 3690. [Google Scholar] [CrossRef] [PubMed]
- Swenson, C.; Zerbe, C.S.; Fennelly, K. Host Variability in NTM disease: Implications for research needs. Front. Microbiol. 2018, 9. [Google Scholar] [CrossRef]
- Flynn, J.L. Lessons from experimental Mycobacterium tuberculosis infections. Microbes Infect. 2006, 8, 1179–1188. [Google Scholar] [CrossRef]
- Obregón-Henao, A.; Arnett, K.A.; Henao-Tamayo, M.; Massoudi, L.; Creissen, E.; Andries, K.; Lenaerts, A.J.; Ordway, D.J. Susceptibility of Mycobacterium abscessus to antimycobacterial drugs in preclinical models. Antimicrob. Agents Chemother. 2015, 59, 6904–6912. [Google Scholar] [CrossRef] [Green Version]
- Ordway, D.; Henao-Tamayo, M.; Smith, E.; Shanley, C.; Harton, M.; Troudt, J.; Bai, X.; Basaraba, R.J.; Orme, I.M.; Chan, E.D. Animal model of Mycobacterium abscessus lung infection. J. Leukoc. Biol. 2008, 83, 1502–1511. [Google Scholar] [CrossRef]
- Bernut, A.; Nguyen-Chi, M.; Halloum, I.; Herrmann, J.-L.; Lutfalla, G.; Kremer, L. Mycobacterium abscessus-Induced granuloma formation is strictly dependent on TNF signaling and neutrophil trafficking. PLOS Pathog. 2016, 12, e1005986. [Google Scholar] [CrossRef]
- Bolz, M.; Ruf, M.T. Buruli ulcer in animals and experimental infection models. In Buruli Ulcer: Mycobacterium Ulcerans Disease; Springer International Publishing: New York, NY, USA, 2019; pp. 159–181. ISBN 9783030111144. [Google Scholar]
- George, K.M.; Pascopella, L.; Welty, D.M.; Small, P.L.C. A Mycobacterium ulcerans toxin, mycolactone, causes apoptosis in guinea pig ulcers and tissue culture cells. Infect. Immun. 2000, 68, 877–883. [Google Scholar] [CrossRef] [Green Version]
- George, K.M.; Chatterjee, D.; Gunawardana, G.; Welty, D.; Hayman, J.; Lee, R.; Small, P.L.C. Mycolactone: A polyketide toxin from Mycobacterium ulcerans required for virulence. Science 1999, 283, 854–857. [Google Scholar] [CrossRef] [Green Version]
- Krieg, R.E.; Hockmeyer, W.T.; Connor, D.H. Toxin of Mycobacterium ulcerans. Production and effects in guinea pig skin. Arch. Dermatol. 1974, 110, 783–788. [Google Scholar] [CrossRef] [PubMed]
- Meijer, A.H.; Spaink, H.P. Host-pathogen interactions made transparent with the zebrafish model. Curr. Drug Targets 2011, 12, 1000–1017. [Google Scholar] [CrossRef] [PubMed]
- Broekhuizen, C.A.N.; Schultz, M.J.; van der Wal, A.C.; Boszhard, L.; de Boer, L.; Vandenbroucke-Grauls, C.M.J.E.; Zaat, S.A.J. Tissue around catheters is a niche for bacteria associated with medical device infection. Crit. Care Med. 2008, 36, 2395–2402. [Google Scholar] [CrossRef] [PubMed]
- Boelens, J.J.; Dankert, J.; Murk, J.L.; Weening, J.J.; van der Poll, T.; Dingemans, K.P.; Koole, L.; Laman, J.D.; Zaat, S.A.J. Biomaterial-associated persistence of Staphylococcus epidermidis in pericatheter macrophages. J. Infect. Dis. 2000, 181, 1337–1349. [Google Scholar] [CrossRef] [Green Version]
- Busscher, H.J.; van der Mei, H.C.; Subbiahdoss, G.; Jutte, P.C.; van den Dungen, J.J.A.M.; Zaat, S.A.J.; Schultz, M.J.; Grainger, D.W. Biomaterial-associated infection: Locating the finish line in the race for the surface. Sci. Transl. Med. 2012, 4, 153rv10. [Google Scholar] [CrossRef] [Green Version]
- Veneman, W.J.; Marín-Juez, R.; de Sonneville, J.; Ordas, A.; Jong-Raadsen, S.; Meijer, A.H.; Spaink, H.P. Establishment and optimization of a high throughput setup to study Staphylococcus epidermidis and Mycobacterium marinum infection as a model for drug discovery. J. Vis. Exp. 2014. [Google Scholar] [CrossRef]
- Davis, J.M.; Clay, H.; Lewis, J.L.; Ghori, N.; Herbomel, P.; Ramakrishnan, L. Real-Time visualization of mycobacterium-macrophage interactions leading to initiation of granuloma formation in zebrafish embryos. Immunity 2002, 17, 693–702. [Google Scholar] [CrossRef] [Green Version]
- Johansen, M.D.; Kremer, L. Large extracellular cord formation in a zebrafish model of Mycobacterium kansasii infection. J. Infect. Dis. 2020, 222, 1046–1050. [Google Scholar] [CrossRef]
- Dupont, C.; Viljoen, A.; Thomas, S.; Roquet-Banères, F.; Herrmann, J.-L.; Pethe, K.; Kremer, L. Bedaquiline inhibits the ATP synthase in Mycobacterium abscessus and is effective in infected zebrafish. Antimicrob. Agents Chemother. 2017, 61. [Google Scholar] [CrossRef] [Green Version]
- Bernut, A.; Le Moigne, V.; Lesne, T.; Lutfalla, G.; Herrmann, J.-L.; Kremer, L. In vivo assessment of drug efficacy against Mycobacterium abscessus using the embryonic zebrafish test system. Antimicrob. Agents Chemother. 2014, 58, 4054–4063. [Google Scholar] [CrossRef] [Green Version]
- Winthrop, K.; Rivera, A.; Engelmann, F.; Rose, S.; Lewis, A.; Ku, J.; Bermudez, L.; Messaoudi, I. A rhesus macaque model of pulmonary nontuberculous mycobacterial disease. Am. J. Respir. Cell Mol. Biol. 2016, 54, 170–176. [Google Scholar] [CrossRef] [PubMed]
- Peters, J.; Caceres, D.M.; Mangat, M.; Griffith, D. Non-human primate model of Non-TB Mycobacteria (NTM) pulmonary disease. Chest 2019, 156, A148–A149. [Google Scholar] [CrossRef]
- Van de Berg, J.L.; Williams-Blangero, S. Advantages and limitations of nonhuman primates as animal models in genetic research on complex diseases. J. Med. Primatol. 1997, 26, 113–119. [Google Scholar] [CrossRef] [PubMed]
- Vallender, E.J.; Miller, G.M. Nonhuman primate models in the genomic era: A paradigm shift. ILAR J. 2013, 54, 154–165. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Izumizono, Y.; Arevalo, S.; Koseki, Y.; Kuroki, M.; Aoki, S. Identification of novel potential antibiotics for tuberculosis by in silico structure-based drug screening. Eur. J. Med. Chem. 2011, 46, 1849–1856. [Google Scholar] [CrossRef]
- Gozalbes, R.; Brun-Pascaud, M.; Garciía-Domenech, R.; Gaálvez, J.; Girard, P.-M.; Doucet, J.-P.; Derouin, F. Prediction of quinolone activity against Mycobacterium avium by molecular topology and virtual computational screening. Antimicrob. Agents Chemother. 2000, 44, 2764–2770. [Google Scholar] [CrossRef] [Green Version]
- Garcia-Garcia, A. New agents active against Mycobacterium avium complex selected by molecular topology: A virtual screening method. J. Antimicrob. Chemother. 2003, 53, 65–73. [Google Scholar] [CrossRef]
- Schulthess, P.; van Wijk, R.C.; Krekels, E.H.J.; Yates, J.W.T.; Spaink, H.P.; van der Graaf, P.H. Outside-in systems pharmacology combines innovative computational methods with high-throughput whole vertebrate studies. CPT Pharmacomet. Syst. Pharmacol. 2018, 7, 285–287. [Google Scholar] [CrossRef]
- Pires, D.E.V.; Ascher, D.B. mycoCSM: Using graph-based signatures to identify safe potent hits against mycobacteria. J. Chem. Inf. Model. 2020, 60, 3450–3456. [Google Scholar] [CrossRef]
- Vente, D.; Arandjelović, O.; Baron, V.O.; Dombay, E.; Gillespie, S.H. Using machine learning for automatic estimation of M. smegmatis cell count from fluorescence microscopy images. In Studies in Computational Intelligence; Springer: Cham, Switzerland, 2020; pp. 57–68. [Google Scholar]
- Lee, J.; Rho, K.; Park, K.H.; Kim, J.-S.; Shin, S.; Kim, T.S.; Kim, S. Utilizing Negative Markers for Identifying Mycobacteria Species based on Mass Spectrometry with Machine Learning Methods. In Proceedings of the 2019 IEEE International Conference on Bioinformatics and Biomedicine (BIBM), San Diego, CA, USA, 18–21 November 2019; IEEE: Piscataway, NJ, USA, 2019; pp. 1331–1337. [Google Scholar]
- Rastogi, N.; Frehel, C.; Ryter, A.; Ohayon, H.; Lesourd, M.; David, H.L. Multiple drug resistance in Mycobacterium avium: Is the wall architecture responsible for exclusion of antimicrobial agents? Antimicrob. Agents Chemother. 1981, 20, 666–677. [Google Scholar] [CrossRef] [Green Version]
- Haemers, A.; Leysen, D.C.; Bollaert, W.; Zhang, M.Q.; Pattyn, S.R. Influence of N substitution on antimycobacterial activity of ciprofloxacin. Antimicrob. Agents Chemother. 1990, 34, 496–497. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Franzblau, S.G.; White, K.E. Comparative in vitro activities of 20 fluoroquinolones against Mycobacterium leprae. Antimicrob. Agents Chemother. 1990, 34, 229–231. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chopra, S.; Matsuyama, K.; Hutson, C.; Madrid, P. Identification of antimicrobial activity among FDA-approved drugs for combating Mycobacterium abscessus and Mycobacterium chelonae. J. Antimicrob. Chemother. 2011, 66, 1533–1536. [Google Scholar] [CrossRef] [PubMed]
- Jeong, J.; Kim, G.; Moon, C.; Kim, H.J.; Kim, T.H.; Jang, J. Pathogen box screening for hit identification against Mycobacterium abscessus. PLoS ONE 2018, 13, e0195595. [Google Scholar] [CrossRef]
- Low, J.L.; Wu, M.-L.; Aziz, D.B.; Laleu, B.; Dick, T. Screening of TB actives for activity against nontuberculous mycobacteria delivers high hit rates. Front. Microbiol. 2017, 8. [Google Scholar] [CrossRef]
- Cowman, S.; Burns, K.; Benson, S.; Wilson, R.; Loebinger, M.R. The antimicrobial susceptibility of non-tuberculous mycobacteria. J. Infect. 2016, 72, 324–331. [Google Scholar] [CrossRef]
- McGuffin, S.; Mullen, S.; Early, J.; Parish, T. 1341. Development of a series of high-throughput screens to identify leads for nontuberculous mycobacteria drug design. Open Forum Infect. Dis. 2019, 6, S485. [Google Scholar] [CrossRef] [Green Version]
- Zimmer, B.L.; DeYoung, D.R.; Roberts, G.D. In vitro synergistic activity of ethambutol, isoniazid, kanamycin, rifampin, and streptomycin against Mycobacterium avium-intracellulare complex. Antimicrob. Agents Chemother. 1982, 22, 148–150. [Google Scholar] [CrossRef] [Green Version]
- Mukherjee, D.; Wu, M.-L.; Teo, J.W.P.; Dick, T. Vancomycin and clarithromycin show synergy against Mycobacterium abscessus in vitro. Antimicrob. Agents Chemother. 2017, 61. [Google Scholar] [CrossRef] [Green Version]
- Machado, D.; Cannalire, R.; Santos Costa, S.; Manfroni, G.; Tabarrini, O.; Cecchetti, V.; Couto, I.; Viveiros, M.; Sabatini, S. Boosting Effect of 2-phenylquinoline efflux inhibitors in combination with macrolides against Mycobacterium smegmatis and Mycobacterium avium. ACS Infect. Dis. 2016, 1, 593–603. [Google Scholar] [CrossRef]
- Felicetti, T.; Machado, D.; Cannalire, R.; Astolfi, A.; Massari, S.; Tabarrini, O.; Manfroni, G.; Barreca, M.L.; Cecchetti, V.; Viveiros, M.; et al. Modifications on C6 and C7 positions of 3-phenylquinolone efflux pump inhibitors led to potent and safe antimycobacterial treatment adjuvants. ACS Infect. Dis. 2019, 5, 982–1000. [Google Scholar] [CrossRef] [PubMed]
- Blondiaux, N.; Moune, M.; Desroses, M.; Frita, R.; Flipo, M.; Mathys, V.; Soetaert, K.; Kiass, M.; Delorme, V.; Djaout, K.; et al. Reversion of antibiotic resistance in Mycobacterium tuberculosis by spiroisoxazoline SMARt-420. Science 2017, 355, 1206–1211. [Google Scholar] [CrossRef] [PubMed]
- Zheng, W.; Sun, W.; Simeonov, A. Drug repurposing screens and synergistic drug-combinations for infectious diseases. Br. J. Pharmacol. 2018, 175, 181–191. [Google Scholar] [CrossRef] [PubMed]
- Worthington, R.J.; Melander, C. Combination approaches to combat multidrug-resistant bacteria. Trends Biotechnol. 2013, 31, 177–184. [Google Scholar] [CrossRef] [Green Version]
- Story-Roller, E.; Maggioncalda, E.C.; Lamichhane, G. Select β-lactam combinations exhibit synergy against Mycobacterium abscessus in vitro. Antimicrob. Agents Chemother. 2019, 63. [Google Scholar] [CrossRef]
- Hetényi, C.; van der Spoel, D. Blind docking of drug-sized compounds to proteins with up to a thousand residues. FEBS Lett. 2006, 580, 1447–1450. [Google Scholar] [CrossRef] [Green Version]
- Johnson, E.O.; Office, E.; Kawate, T.; Orzechowski, M.; Hung, D.T. Discovery of a novel synergistic antimycobacterial combination targeting EfpA using large-scale chemical-genetics. bioRxiv 2019. [Google Scholar] [CrossRef]
- Wallis, R.S.; Hafner, R. Advancing host-directed therapy for tuberculosis. Nat. Rev. Immunol. 2015, 15, 255–263. [Google Scholar] [CrossRef]
- Tobin, D.M. Host-Directed therapies for tuberculosis. Cold Spring Harb. Perspect. Med. 2015, 5, a021196. [Google Scholar] [CrossRef]
- Schwegmann, A.; Brombacher, F. Host-Directed drug targeting of factors hijacked by pathogens. Sci. Signal. 2008, 1, re8. [Google Scholar] [CrossRef]
- Hawn, T.R.; Shah, J.A.; Kalman, D. New tricks for old dogs: Countering antibiotic resistance in tuberculosis with host-directed therapeutics. Immunol. Rev. 2015, 264, 344–362. [Google Scholar] [CrossRef] [Green Version]
- Cai, S.; Sato, K.; Shimizu, T.; Yamabe, S.; Hiraki, M.; Sano, C.; Tomioka, H. Antimicrobial activity of picolinic acid against extracellular and intracellular Mycobacterium avium complex and its combined activity with clarithromycin, rifampicin and fluoroquinolones. J. Antimicrob. Chemother. 2006, 57, 85–93. [Google Scholar] [CrossRef] [PubMed]
- Gomes, M.S.; Dom, G.; Pedrosa, J.; Boelaert, J.R.; Appelberg, R. Effects of iron deprivation on Mycobacterium avium growth. Tuber. Lung Dis. 1999, 79, 321–328. [Google Scholar] [CrossRef] [PubMed]
- Pais, T.F.; Appelberg, R. Macrophage control of mycobacterial growth induced by picolinic acid is dependent on host cell apoptosis. J. Immunol. 2000, 164, 389–397. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tanigawa, K.; Suzuki, K.; Nakamura, K.; Akama, T.; Kawashima, A.; Wu, H.; Hayashi, M.; Takahashi, S.-I.; Ikuyama, S.; Ito, T.; et al. Expression of adipose differentiation-related protein (ADRP) and perilipin in macrophages infected with Mycobacterium leprae. FEMS Microbiol. Lett. 2008, 289, 72–79. [Google Scholar] [CrossRef] [Green Version]
- Degang, Y.; Akama, T.; Hara, T.; Tanigawa, K.; Ishido, Y.; Gidoh, M.; Makino, M.; Ishii, N.; Suzuki, K. Clofazimine modulates the expression of lipid metabolism proteins in Mycobacterium leprae-infected macrophages. PLoS Negl. Trop. Dis. 2012, 6, e1936. [Google Scholar] [CrossRef] [Green Version]
- Tang, J.; Chen, Q.; Guo, J.; Yang, L.; Tao, Y.; Li, L.; Miao, H.; Feng, H.; Chen, Z.; Zhu, G. Minocycline attenuates neonatal germinal-matrix-hemorrhage-induced neuroinflammation and brain edema by activating cannabinoid receptor 2. Mol. Neurobiol. 2016, 53, 1935–1948. [Google Scholar] [CrossRef]
- Montes-Worboys, A.; Brown, S.; Regev, D.; Bellew, B.F.; Mohammed, K.A.; Faruqi, I.; Sharma, P.; Moudgil, B.; Antony, V.B. Targeted delivery of amikacin into granuloma. Am. J. Respir. Crit. Care Med. 2010, 182, 1546–1553. [Google Scholar] [CrossRef] [Green Version]
- Rao, M.; Ippolito, G.; Mfinanga, S.; Ntoumi, F.; Yeboah-Manu, D.; Vilaplana, C.; Zumla, A.; Maeurer, M. Improving treatment outcomes for MDR-TB—Novel host-directed therapies and personalised medicine of the future. Int. J. Infect. Dis. 2019, 80, S62–S67. [Google Scholar] [CrossRef] [Green Version]


{kind=link}
{kind=link}
| Runyon Classification | NTM Species | Pathogenesis in Humans |
|---|---|---|
| Photochromogens Runyon type I | M. kansasii [16], M. simiae [17] | Pulmonary infections Skin infections Disseminated infections |
| M. marinum [18] | Skin and soft tissue infections Disseminated infections | |
| Scotochromogens Runyon type II | M. gordonae [19] | Pulmonary infections Skin infections Disseminated infections |
| M. scrofulaceum [20] | Cervical lymphadenitis among children Pulmonary infections Disseminated infections | |
| Non-photochromogens Runyon type III | M. avium complex (M. avium and M. intracellulare) [21] | Pulmonary MAC infections Disseminated infections (mostly in AIDS patients) MAC associated lymphadenitis (in young kids and people with normal immune systems) |
| M. malmoense [22] | Pulmonary infections Disseminated infections | |
| M. ulcerans [18] | Skin diseases (Buruli ulcers) | |
| Rapid growing Runyon type IV | M. abscessus [23] | Pulmonary infections Skin and Soft tissue disease Central nervous system infections Disseminated infections |
| M. chelonae [24] | Skin and soft tissue infections Pulmonary infections Disseminated infections | |
| M. smegmatis | Widely regarded as nonpathogenic |
| In Silico | In Vitro | In Vivo | |
|---|---|---|---|
| Methods employed | Structure–activity relations, Molecular simulations, Comparative genomics | Antimicrobial effect tests on cultured cells | Tests on live infected animals |
| Main insights | Molecular basis for drug action | Molecular and cellular effect of drug action | Whole-organism level of drug action |
| Advantages | High throughput, low-cost, no need for actual chemical synthesis of compounds or bacterial growth | Relatively simple systems and lower cost and time involvement, easy to handle, scalable | Closer to the actual physiological environment |
| Limitations | Requires prior information and complicated models to simulate molecular events such as docking and drug-target interactions | Needs a high level of standardization and careful experimentation for reproducibility, may not reproduce clinical situations | Requires careful model selection, large organism response is less predictable, ethical considerations, high economical costs |
| Best-fit stage in drug discovery | Primary (for narrowing the search of potential candidates) or secondary (for optimizing compounds to species-specific targets) | Secondary (for screening initial targets and efficacy determination) | Tertiary (for preclinical evaluation) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Saxena, S.; Spaink, H.P.; Forn-Cuní, G. Drug Resistance in Nontuberculous Mycobacteria: Mechanisms and Models. Biology 2021, 10, 96. https://doi.org/10.3390/biology10020096
Saxena S, Spaink HP, Forn-Cuní G. Drug Resistance in Nontuberculous Mycobacteria: Mechanisms and Models. Biology. 2021; 10(2):96. https://doi.org/10.3390/biology10020096
Chicago/Turabian StyleSaxena, Saloni, Herman P. Spaink, and Gabriel Forn-Cuní. 2021. "Drug Resistance in Nontuberculous Mycobacteria: Mechanisms and Models" Biology 10, no. 2: 96. https://doi.org/10.3390/biology10020096