
Characterization and Expression Analysis of the C-Type Lectin Ladderlectin in Litopenaeus vannamei Post-WSSV Infection

Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Experimental Animals
2.2. Tissue Distribution and Expression Profiles of LvLL after WSSV Challenge
2.3. RNA Extraction and cDNA Synthesis
2.4. Molecular Cloning and Sequencing of LvLL
2.5. Bioinformatics Analysis
2.6. RT-PCR and Statistical Analysis
2.7. Synthesis of Double-Stranded RNA
2.8. Analysis of dsRNA Interference Efficiency
2.9. Survival Rate Analysis
2.10. Investigating Immune Response and Gene Expression after LvLL Knockdown
2.10.1. Experimental Setting
2.10.2. TaqMan RT-PCR
2.10.3. Gene Expression Analysis
2.11. Screening of LvLL SNP and Correlation Analysis with WSSV Resistance
3. Results
3.1. Characterization and Analysis of LvLL
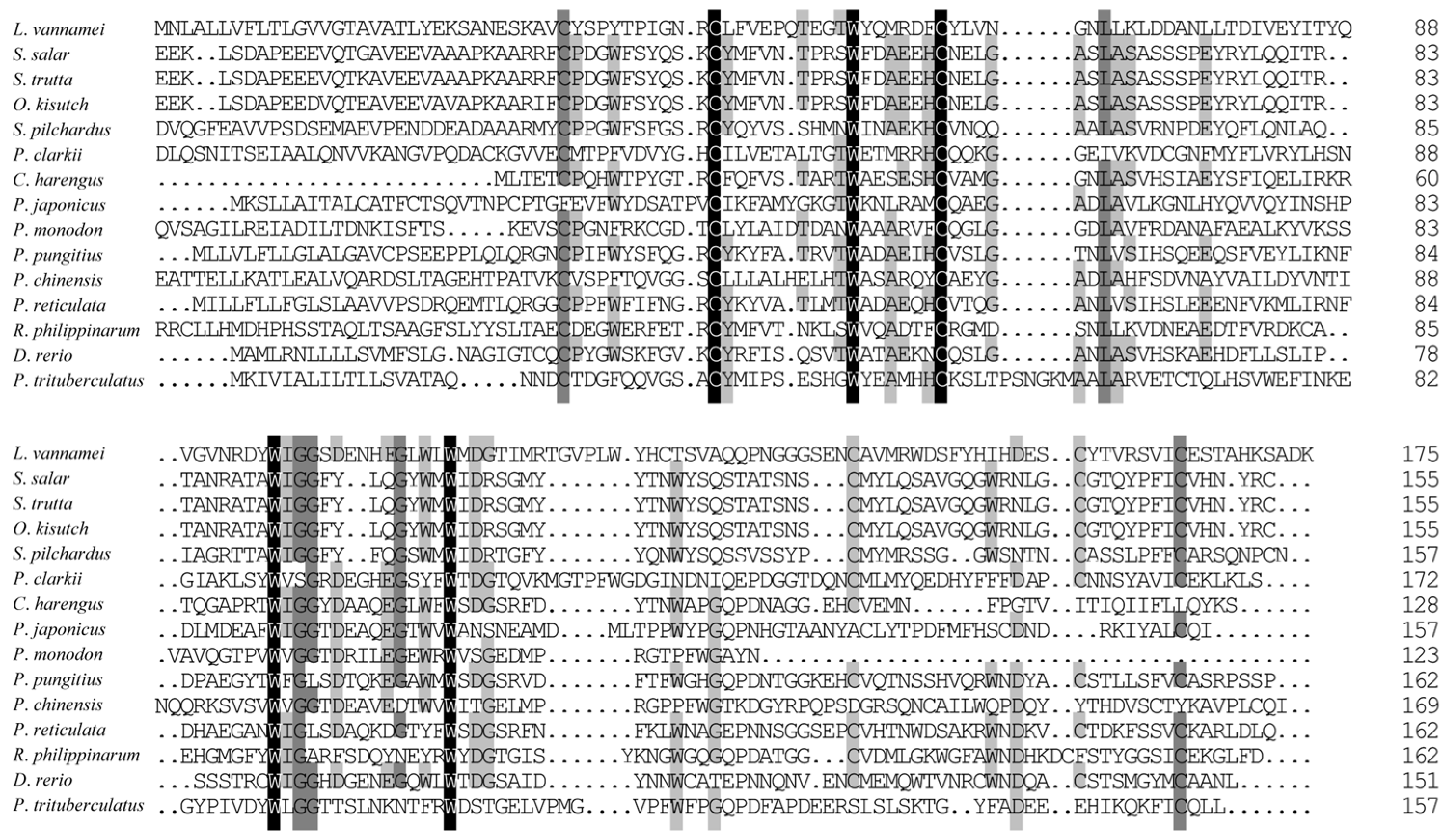
3.2. Homologous and Phylogenetic Analysis of Ladderlectin
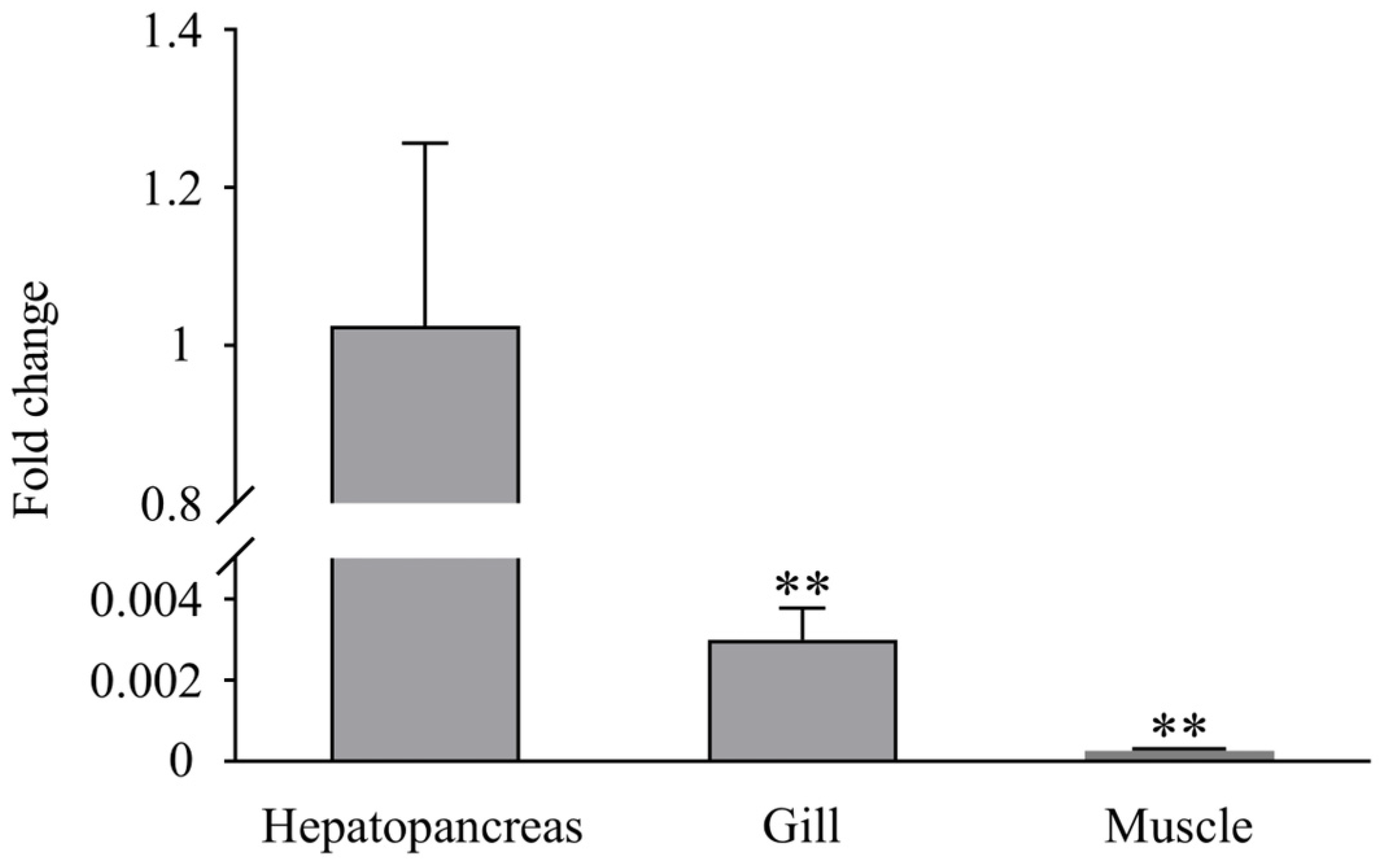
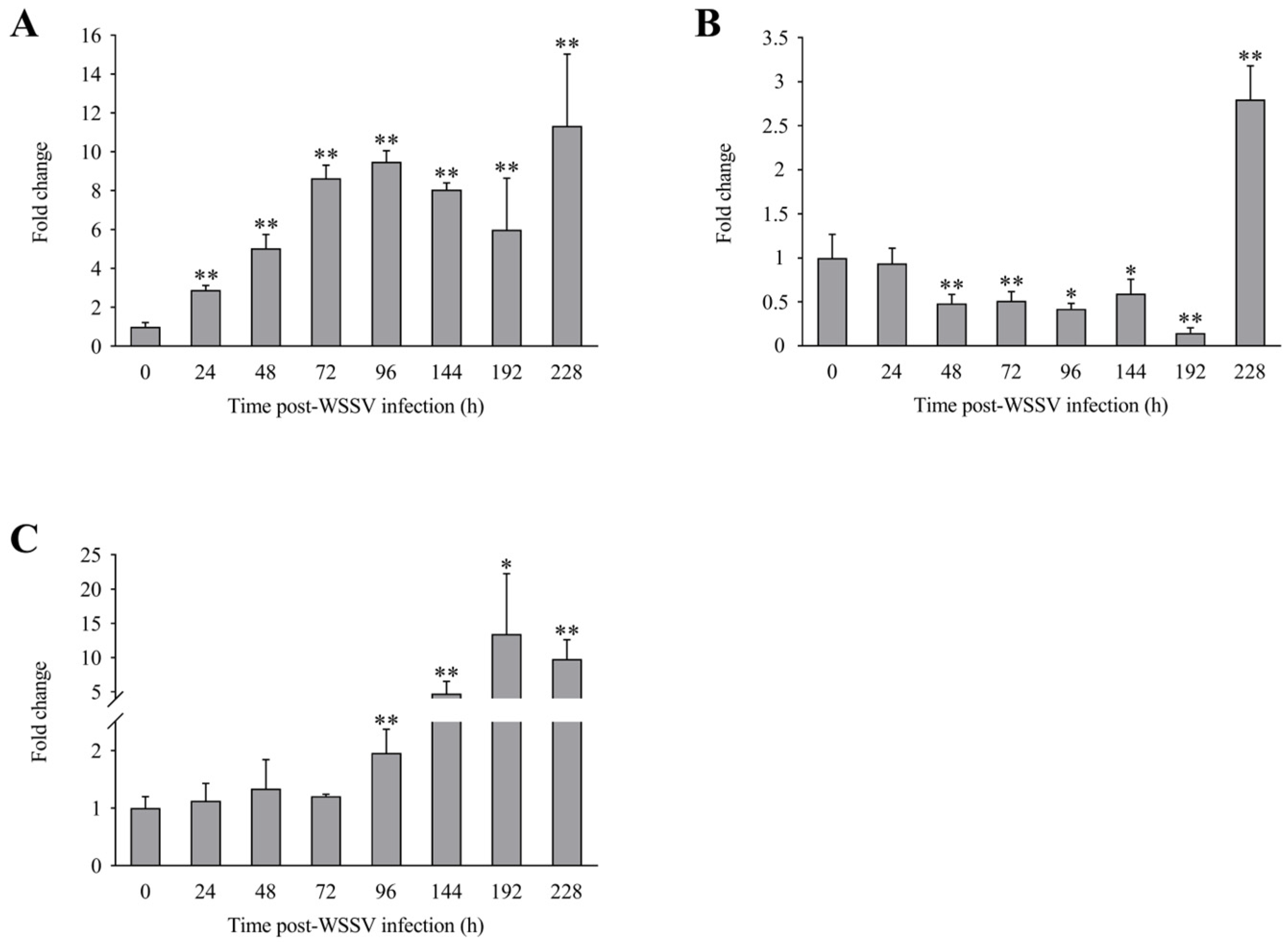
3.3. Expression Profiles of LvLL after WSSV Infection
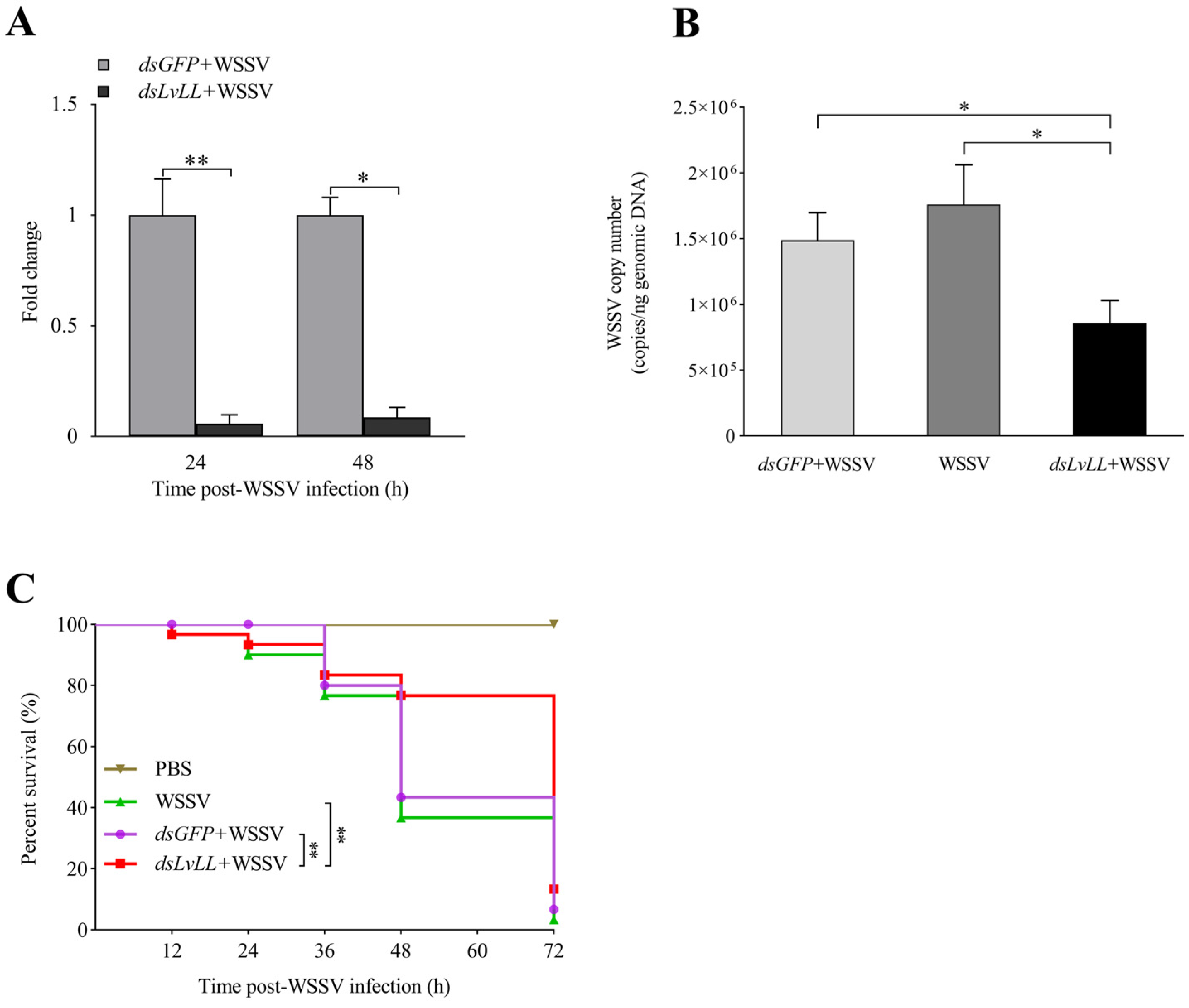
3.4. WSSV Infection Was Suppressed after LvLL Knockdown
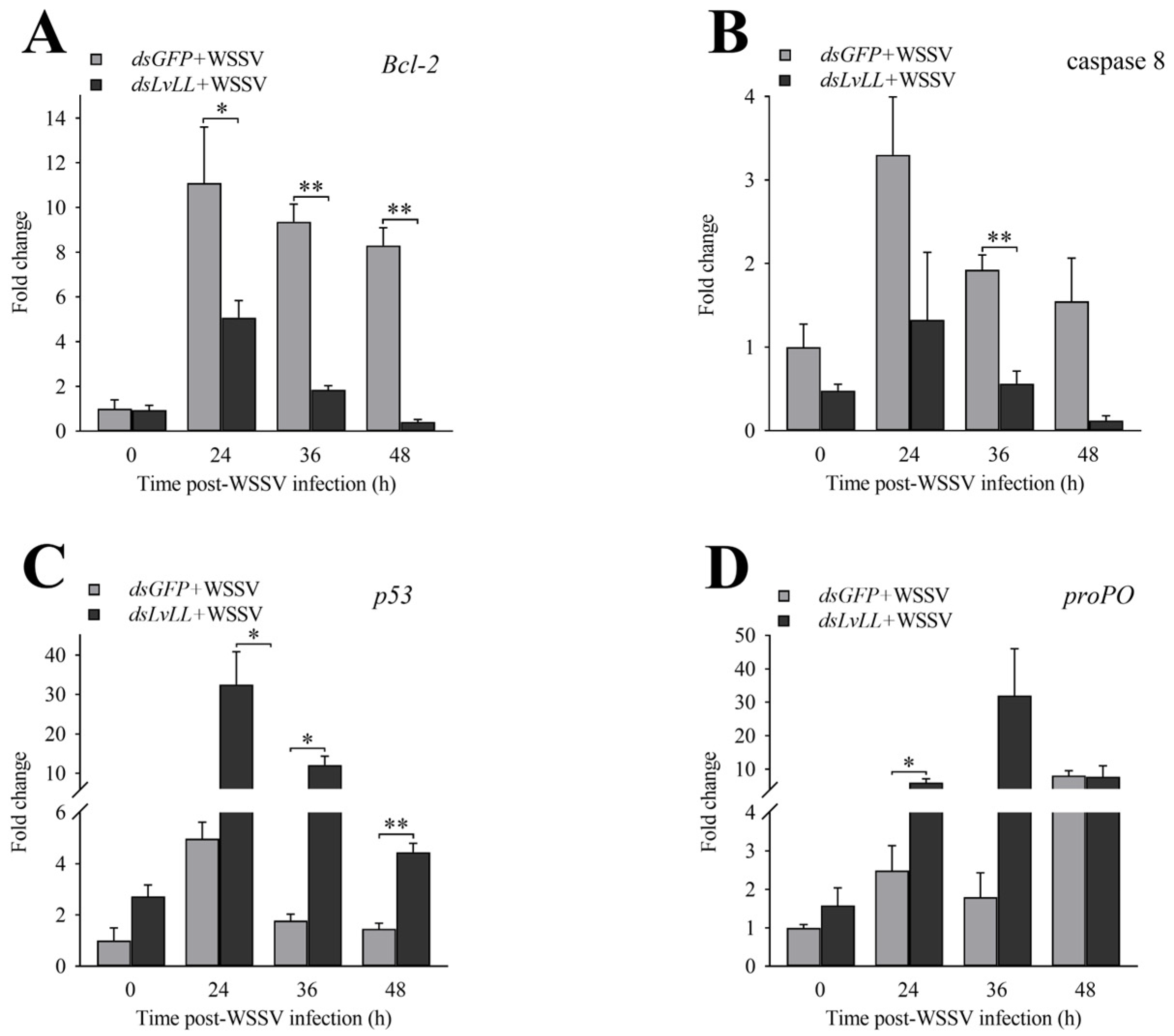
3.5. Expression Profiles of Genes Related to Innate Immunity after LvLL Knockdown
3.6. SNP Detection and Polymorphism Analysis
3.7. SNPs Associated with WSSV Resistance
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Flegel, T.W. Historic emergence, impact and current status of shrimp pathogens in Asia. J. Invertebr. Pathol. 2012, 110, 166–173. [Google Scholar] [CrossRef] [PubMed]
- Seibert, C.H.; Pinto, A.R. Challenges in shrimp aquaculture due to viral diseases: Distribution and biology of the five major penaeid viruses and interventions to avoid viral incidence and dispersion. Braz. J. Microbiol. 2012, 43, 857–864. [Google Scholar] [CrossRef]
- Lightner, D.V. Virus diseases of farmed shrimp in the Western Hemisphere (the Americas): A review. J. Invertebr. Pathol. 2011, 106, 110–130. [Google Scholar] [CrossRef]
- Pradeep, B.; Rai, P.; Mohan, S.A.; Shekhar, M.S.; Karunasagar, I. Biology, Host Range, Pathogenesis and Diagnosis of White spot syndrome virus. Indian J. Virol. 2012, 23, 161–174. [Google Scholar] [CrossRef] [PubMed]
- Wongteerasupaya, C.; Vickers, J.E.; Sriurairatana, S.; Nash, G.L.; Akarajamorn, A.; Boonsaeng, V.; Panyim, S.; Tassanakajon, A.; Withyachumnarnkul, B.; Flegel, T.W. A non-occluded systemic baculovirus that occurs in cells of ectodermal and mesodermal origin and causes high mortality in the black tiger prawn P. monodon. Dis. Aquat. Org. 1995, 21, 69–77. [Google Scholar] [CrossRef]
- Akira, S.; Uematsu, S.; Takeuchi, O. Pathogen recognition and innate immunity. Cell 2006, 124, 783–801. [Google Scholar] [CrossRef] [PubMed]
- Hoffmann, J.A.; Kafatos, F.C.; Janeway, C.A.; Ezekowitz, R.A.B. Phylogenetic perspectives in innate immunity. Science 1999, 284, 1313–1318. [Google Scholar] [CrossRef]
- De Gregorio, E.; Spellman, P.T.; Rubin, G.M.; Lemaitre, B. Genome-wide analysis of the Drosophila immune response by using oligonucleotide microarrays. Proc. Natl. Acad. Sci. USA 2001, 98, 12590–12595. [Google Scholar] [CrossRef]
- Luo, M.; Yang, L.; Wang, Z.A.; Zuo, H.; Weng, S.; He, J.; Xu, X. A novel C-type lectin with microbiostatic and immune regulatory functions from Litopenaeus vannamei. Fish Shellfish Immunol. 2019, 93, 361–368. [Google Scholar] [CrossRef]
- Wang, L.; Wang, L.; Zhang, D.; Li, F.; Wang, M.; Huang, M.; Zhang, H.; Song, L. A novel C-type lectin from crab Eriocheir sinensis functions as pattern recognition receptor enhancing cellular encapsulation. Fish Shellfish Immunol. 2013, 34, 832–842. [Google Scholar] [CrossRef]
- Zelensky, A.N.; Gready, J.E. The C-type lectin-like domain superfamily. FEBS J. 2005, 272, 6179–6217. [Google Scholar] [CrossRef]
- Drickamer, K. Evolution of Ca(2+)-dependent animal lectins. Prog. Nucl. Res. Molec. Biol. 1993, 45, 207–232. [Google Scholar]
- Chen, D.; Meng, X.; Xu, J.; Yu, J.; Meng, M.; Wang, J. PcLT, a novel C-type lectin from Procambarus clarkii, is involved in the innate defense against Vibrio alginolyticus and WSSV. Dev. Comp. Immunol. 2013, 39, 255–264. [Google Scholar] [CrossRef]
- Zhang, X.; Pan, L.; Yu, J.; Huang, H. One recombinant C-type lectin (LvLec) from white shrimp Litopenaeus vannamei affected the haemocyte immune response in vitro. Fish Shellfish Immunol. 2019, 89, 35–42. [Google Scholar] [CrossRef] [PubMed]
- Rogers, N.C.; Slack, E.C.; Edwards, A.D.; Nolte, M.A.; Schulz, O.; Schweighoffer, E.; Williams, D.L.; Gordon, S.; Tybulewicz, V.L.; Brown, G.D.; et al. Syk-Dependent Cytokine Induction by Dectin-1 Reveals a Novel Pattern Recognition Pathway for C Type Lectins. Immunity 2005, 22, 507–517. [Google Scholar] [CrossRef] [PubMed]
- van Vliet, S.J.; García-Vallejo, J.J.; van Kooyk, Y. Dendritic cells and C-type lectin receptors: Coupling innate to adaptive immune responses. Immunol. Cell Biol. 2008, 86, 580–587. [Google Scholar] [CrossRef] [PubMed]
- Junkunlo, K.; Prachumwat, A.; Tangprasittipap, A.; Senapin, S.; Borwornpinyo, S.; Flegel, T.W.; Sritunyalucksana, K. A novel lectin domain-containing protein (LvCTLD) associated with response of the whiteleg shrimp Penaeus (Litopenaeus) vannamei to yellow head virus (YHV). Dev. Comp. Immunol. 2012, 37, 334–341. [Google Scholar] [CrossRef]
- Su, Y.; Liu, Y.; Gao, F.; Cui, Z. A novel C-type lectin with a YPD motif from Portunus trituberculatus (PtCLec1) mediating pathogen recognition and opsonization. Dev. Comp. Immunol. 2020, 106, 103609. [Google Scholar] [CrossRef]
- Kwankaew, P.; Praparatana, R.; Runsaeng, P.; Utarabhand, P. An alternative function of C-type lectin comprising low-density lipoprotein receptor domain from Fenneropenaeus merguiensis to act as a binding receptor for viral protein and vitellogenin. Fish Shellfish Immunol. 2018, 74, 295–308. [Google Scholar] [CrossRef]
- Luo, Z.; Zhang, J.; Li, F.; Zhang, X.; Liu, C.; Xiang, J. Identification of a novel C-type lectin from the shrimp Litopenaeus vannamei and its role in defense against pathogens infection. Chin. J. Oceanol. Limnol. 2011, 29, 942–951. [Google Scholar] [CrossRef]
- Zhao, Z.Y.; Yin, Z.X.; Xu, X.P.; Weng, S.P.; Rao, X.Y.; Dai, Z.X.; Luo, Y.W.; Yang, G.; Li, Z.S.; Guan, H.J.; et al. A novel C-type lectin from the shrimp Litopenaeus vannamei possesses anti-white spot syndrome virus activity. J. Virol. 2009, 83, 347–356. [Google Scholar] [CrossRef] [PubMed]
- Huang, X.; Xu, Y.; Zhao, Y.; Cao, X.; Wang, D.; Yan, J.; Wei, T.; Dai, X.; Xu, Z.; Ren, Q. Characterization of four spliced isoforms of a transmembrane C-type lectin from Procambarus clarkii and their function in facilitating WSSV infection. Fish Shellfish Immunol. 2022, 127, 1127–1138. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.; Kumar, R.; Liu, C.; Lin, S.; Wang, H. A novel C-type lectin LvCTL 4.2 has antibacterial activity but facilitates WSSV infection in shrimp (L. vannamei). Dev. Comp. Immunol. 2022, 126, 104239. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.W.; Xu, Y.H.; Xu, J.D.; Zhao, X.F.; Wang, J.X. Collaboration between a soluble C-type lectin and calreticulin facilitates white spot syndrome virus infection in shrimp. J. Immunol. 2014, 193, 2106–2117. [Google Scholar] [CrossRef] [PubMed]
- Purcell, S.; Neale, B.; Todd-Brown, K.; Thomas, L.; Ferreira, M.A.R.; Bender, D.; Maller, J.; Sklar, P.; de Bakker, P.I.W.; Daly, M.J.; et al. PLINK: A Tool Set for Whole-Genome Association and Population-Based Linkage Analyses. Am. J. Hum. Genet. 2007, 81, 559–575. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Zheng, M.; Liu, J.; Jin, Z.; Wang, C.; Gao, M.; Zhang, H.; Zhang, X.; Xia, X. LDLa containing C-type lectin mediates phagocytosis of V. anguillarum and regulates immune effector genes in shrimp. Fish Shellfish Immunol. 2024, 145, 109361. [Google Scholar] [CrossRef]
- Xia, X.; You, M.; Rao, X.J.; Yu, X.Q. Insect C-type lectins in innate immunity. Dev. Comp. Immunol. 2018, 83, 70–79. [Google Scholar] [CrossRef]
- Ma, T.H.; Tiu, S.H.; He, J.G.; Chan, S.M. Molecular cloning of a C-type lectin (LvLT) from the shrimp Litopenaeus vannamei: Early gene down-regulation after WSSV infection. Fish Shellfish Immunol. 2007, 23, 430–437. [Google Scholar] [CrossRef]
- Hoi-Tung Ma, T.; Benzie, J.A.H.; He, J.; Chan, S. PmLT, a C-type lectin specific to hepatopancreas is involved in the innate defense of the shrimp Penaeus monodon. J. Invertebr. Pathol. 2008, 99, 332–341. [Google Scholar] [CrossRef]
- Luo, T.; Zhang, X.; Shao, Z.; Xu, X. PmAV, a novel gene involved in virus resistance of shrimp Penaeus monodon. FEBS Lett. 2003, 551, 53–57. [Google Scholar] [CrossRef]
- Sun, Y.D.; Fu, L.D.; Jia, Y.P.; Du, X.J.; Wang, J.X. A hepatopancreas-specific C-type lectin from the Chinese shrimp Fenneropenaeus chinensis exhibits antimicrobial activity. Mol. Immunol. 2008, 45, 348–361. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.C.; Li, F.H.; Dong, B.; Wang, B.; Luan, W.; Zhang, X.J.; Zhang, L.S.; Xiang, J.H. Molecular cloning, characterization and expression analysis of a putative C-type lectin (Fclectin) gene in Chinese shrimp Fenneropenaeus chinensis. Mol. Immunol. 2007, 44, 598–607. [Google Scholar] [CrossRef]
- Luo, T.; Yang, H.; Li, F.; Zhang, X.; Xu, X. Purification, characterization and cDNA cloning of a novel lipopolysaccharide-binding lectin from the shrimp Penaeus monodon. Dev. Comp. Immunol. 2006, 30, 607–617. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Vasta, G.R.; Wang, J. The functional relevance of shrimp C-type lectins in host-pathogen interactions. Dev. Comp. Immunol. 2020, 109, 103708. [Google Scholar] [CrossRef] [PubMed]
- Wei, X.; Liu, X.; Yang, J.; Fang, J.; Qiao, H.; Zhang, Y.; Yang, J. Two C-type lectins from shrimp Litopenaeus vannamei that might be involved in immune response against bacteria and virus. Fish Shellfish Immunol. 2012, 32, 132–140. [Google Scholar] [CrossRef]
- Wang, X.W.; Xu, W.T.; Zhang, X.W.; Zhao, X.F.; Yu, X.Q.; Wang, J.X. A C-type lectin is involved in the innate immune response of Chinese white shrimp. Fish Shellfish Immunol. 2009, 27, 556–562. [Google Scholar] [CrossRef]
- Xu, W.T.; Wang, X.W.; Zhang, X.W.; Zhao, X.F.; Yu, X.Q.; Wang, J.X. A new C-type lectin (FcLec5) from the Chinese white shrimp Fenneropenaeus chinensis. Amino Acids 2010, 39, 1227–1239. [Google Scholar] [CrossRef]
- Senghoi, W.; Runsaeng, P.; Utarabhand, P. FmLC5, a putative galactose-binding C-type lectin with two QPD motifs from the hemocytes of Fenneropenaeus merguiensis participates in shrimp immune defense. J. Invertebr. Pathol. 2017, 150, 136–144. [Google Scholar] [CrossRef]
- Liang, Z.; Yang, L.; Zheng, J.; Zuo, H.; Weng, S.; He, J.; Xu, X. A low-density lipoprotein receptor (LDLR) class A domain-containing C-type lectin from Litopenaeus vannamei plays opposite roles in antibacterial and antiviral responses. Dev. Comp. Immunol. 2019, 92, 29–34. [Google Scholar] [CrossRef]
- Gramberg, T.; Hofmann, H.; Möller, P.; Lalor, P.F.; Marzi, A.; Geier, M.; Krumbiegel, M.; Winkler, T.; Kirchhoff, F.; Adams, D.H.; et al. LSECtin interacts with filovirus glycoproteins and the spike protein of SARS coronavirus. Virology 2005, 340, 224–236. [Google Scholar] [CrossRef]
- Lambert, A.A.; Gilbert, C.; Richard, M.; Beaulieu, A.D.; Tremblay, M.J. The C-type lectin surface receptor DCIR acts as a new attachment factor for HIV-1 in dendritic cells and contributes to trans- and cis-infection pathways. Blood 2008, 112, 1299–1307. [Google Scholar] [CrossRef] [PubMed]
- Ke, F.; Grabow, S.; Kelly, G.L.; Lin, A.; O’Reilly, L.A.; Strasser, A. Impact of the combined loss of BOK, BAX and BAK on the hematopoietic system is slightly more severe than compound loss of BAX and BAK. Cell Death Discov. 2015, 6, e1938. [Google Scholar] [CrossRef] [PubMed]
- Dawson, S.J.; Makretsov, N.; Blows, F.M.; Driver, K.E.; Provenzano, E.; Le Quesne, J.; Baglietto, L.; Severi, G.; Giles, G.G.; McLean, C.A.; et al. BCL2 in breast cancer: A favourable prognostic marker across molecular subtypes and independent of adjuvant therapy received. Br. J. Cancer 2010, 103, 668–675. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Gong, Y.; Zheng, H.; Ma, H.; Aweya, J.J.; Zhang, Y.; Chen, X.; Li, S.-k. SpBcl2 promotes WSSV infection by suppressing apoptotic activity of hemocytes in mud crab, Scylla paramamosain. Dev. Comp. Immunol. 2019, 100, 103421. [Google Scholar] [CrossRef] [PubMed]
- Su, H.; Bidère, N.; Zheng, L.; Cubre, A.; Sakai, K.; Dale, J.; Salmena, L.; Hakem, R.; Straus, S.; Lenardo, M. Requirement for caspase-8 in NF-kappaB activation by antigen receptor. Science 2005, 307, 1465–1468. [Google Scholar] [CrossRef]
- Meulmeester, E.; Jochemsen, A.G. p53: A guide to apoptosis. Curr. Cancer Drug Targets 2008, 8, 87–97. [Google Scholar] [CrossRef]
- Gong, Y.; Ju, C.; Zhang, X. The miR-1000-p53 pathway regulates apoptosis and virus infection in shrimp. Fish Shellfish Immunol. 2015, 46, 516–522. [Google Scholar] [CrossRef]
- Sutthangkul, J.; Amparyup, P.; Charoensapsri, W.; Senapin, S.; Phiwsaiya, K.; Tassanakajon, A. Suppression of shrimp melanization during white spot syndrome virus infection. J. Biol. Chem. 2015, 290, 6470–6481. [Google Scholar] [CrossRef]
- Amparyup, P.; Charoensapsri, W.; Tassanakajon, A. Prophenoloxidase system and its role in shrimp immune responses against major pathogens. Fish Shellfish Immunol. 2013, 34, 990–1001. [Google Scholar] [CrossRef]
- Fang, Z.Y.; Li, D.; Li, X.J.; Zhang, X.; Zhu, Y.T.; Li, W.W.; Wang, Q. A single CRD C-type lectin from Eriocheir sinensis (EsLecB) with microbial-binding, antibacterial prophenoloxidase activation and hem-encapsulation activities. Fish Shellfish Immunol. 2016, 50, 175–190. [Google Scholar] [CrossRef]
- Chen, B.; Cole, J.W.; Grond-Ginsbach, C. Departure from Hardy Weinberg Equilibrium and Genotyping Error. Front. Genet. 2017, 8, 167. [Google Scholar] [CrossRef] [PubMed]
- Selvam, R.; Murali, N.; Thiruvenkadan, A.K.; Saravanakumar, R.; Ponnudurai, G.; Jawahar, T.P. Single-nucleotide polymorphism-based genetic diversity analysis of the Kilakarsal and Vembur sheep breeds. Vet. World 2017, 10, 549–555. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer or Probe Name | Sequences (5′–3′) |
|---|---|
| LvLL-F | CTTGAAGGCAACACAAACGC |
| LvLL-R | TTTGCATTCCTCAACTAAAACTACA |
| qLvLL-F | TTCGTGCTCCTGCTGTCCTT |
| qLvLL-R | CAAACTCTCTGGGCGTTGGGT |
| 18S-F | TATACGCTAGTGGAGCTGGAA |
| 18S-R | GGGGAGGTAGTGACGAAAAAT |
| WSSV-F | TGGTCCCGTCCTCATCTCAG |
| WSSV-R | GCTGCCTTGCCGGAAATTA |
| WSSV-probe | AGCCATGAAGAATGCCGTCTATCACACA |
| Bcl-2-F | GCTATGTGTCCTTTGTGGCT |
| Bcl-2-R | TGAACTTGGCAATGGTAACTG |
| p53-F | CCCCACATCCACGGAGATA |
| p53-R | CAATGGCGGCTGATACACC |
| caspase 8-F | GGCGACAAGATGAGGCAA |
| caspase 8-R | CAGGGTGAGGGAGAGAAAACT |
| proPO-F | TACATGCACCAGCAAATTATCG |
| proPO-R | AGTTTGGGGAAGTAGCCGTC |
| dsGFP-Fi | GCGTAATACGACTCACTATAGGTGGTCCCAATTCTCGTGGAAC |
| dsGFP-Ri | GCGTAATACGACTCACTATAGGCTTGAAGTTGACCTTGATGCC |
| dsLvLL-Fi | GCGTAATACGACTCACTATAGGCGCGCAAAATGATGTTCTTCGT |
| dsLvLL-Ri | GCGTAATACGACTCACTATAGGTGGAGCGCTACAATCATAATCAAA |
| Name | Location (bp) | Type of Base | Ho | He | MAF | PIC | HWE (p-Value) | |||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| S Group | R Group | S Group | R Group | S Group | R Group | S Group | R Group | S Group | R Group | |||
| SNP1 | ORF-23 | T/C | 0.200 | 0.275 | 0.455 | 0.492 | 0.350 | 0.438 | 0.351 | 0.371 | 0.000 | 0.009 |
| SNP2 | ORF-204 | C/T | 0.450 | 0.600 | 0.455 | 0.495 | 0.350 | 0.450 | 0.351 | 0.372 | 1.000 | 0.335 |
| SNP3 | ORF-492 | T/A | 0.025 | 0.000 | 0.025 | 0.000 | 0.013 | 0.000 | 0.024 | 0.000 | 1.000 | 1.000 |
| Name | Genotype | S Group | R Group | χ2 | p-Value |
|---|---|---|---|---|---|
| SNP1 | TT | 22 | 12 | 5.230 | 0.073 |
| CC | 10 | 17 | |||
| CT | 8 | 11 | |||
| SNP2 | TT | 17 | 10 | 2.763 | 0.251 |
| CC | 5 | 6 | |||
| CT | 18 | 24 | |||
| SNP3 | TT | 39 | 40 | 1.013 | 0.314 |
| AT | 1 | 0 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xue, Q.; Yang, B.; Luo, K.; Luan, S.; Kong, J.; Fu, Q.; Cao, J.; Chen, B.; Dai, P.; Xing, Q.; et al. Characterization and Expression Analysis of the C-Type Lectin Ladderlectin in Litopenaeus vannamei Post-WSSV Infection. Biology 2024, 13, 758. https://doi.org/10.3390/biology13100758
Xue Q, Yang B, Luo K, Luan S, Kong J, Fu Q, Cao J, Chen B, Dai P, Xing Q, et al. Characterization and Expression Analysis of the C-Type Lectin Ladderlectin in Litopenaeus vannamei Post-WSSV Infection. Biology. 2024; 13(10):758. https://doi.org/10.3390/biology13100758
Chicago/Turabian StyleXue, Qian, Bingbing Yang, Kun Luo, Sheng Luan, Jie Kong, Qiang Fu, Jiawang Cao, Baolong Chen, Ping Dai, Qun Xing, and et al. 2024. "Characterization and Expression Analysis of the C-Type Lectin Ladderlectin in Litopenaeus vannamei Post-WSSV Infection" Biology 13, no. 10: 758. https://doi.org/10.3390/biology13100758