Exploration of Protease Resources in the Gut of Omnivorous Gryllotalpa orientalis (Orthoptera: Gryllotalpidae)

 , ,
, ,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Analysis of Proteases Derived from Gut Extract
2.2. Analysis of Proteases Derived from Gut Bacteria
2.3. Statistical Analysis
3. Results
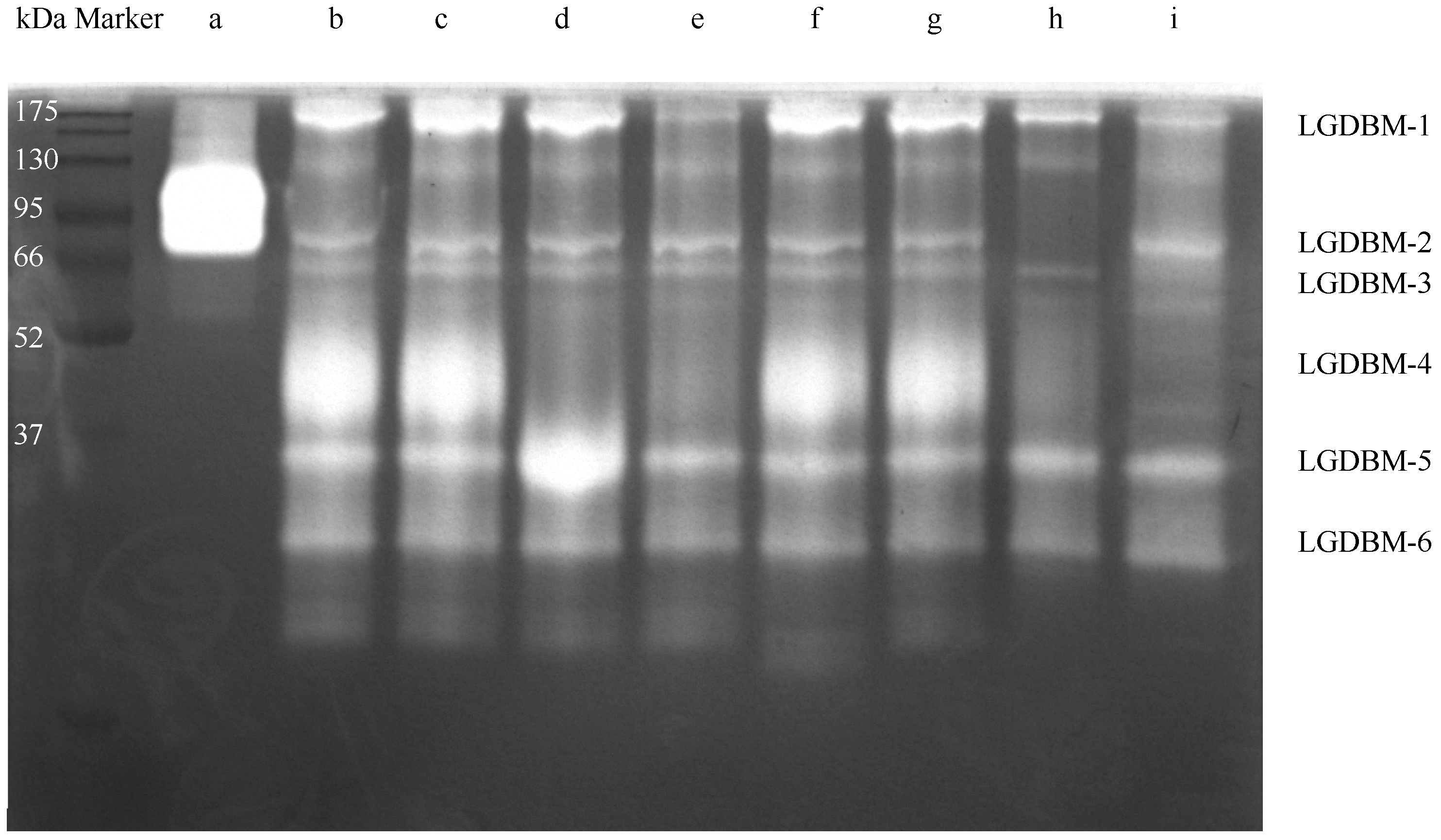
3.1. Exploration of Protease Resources from the Gut Extract
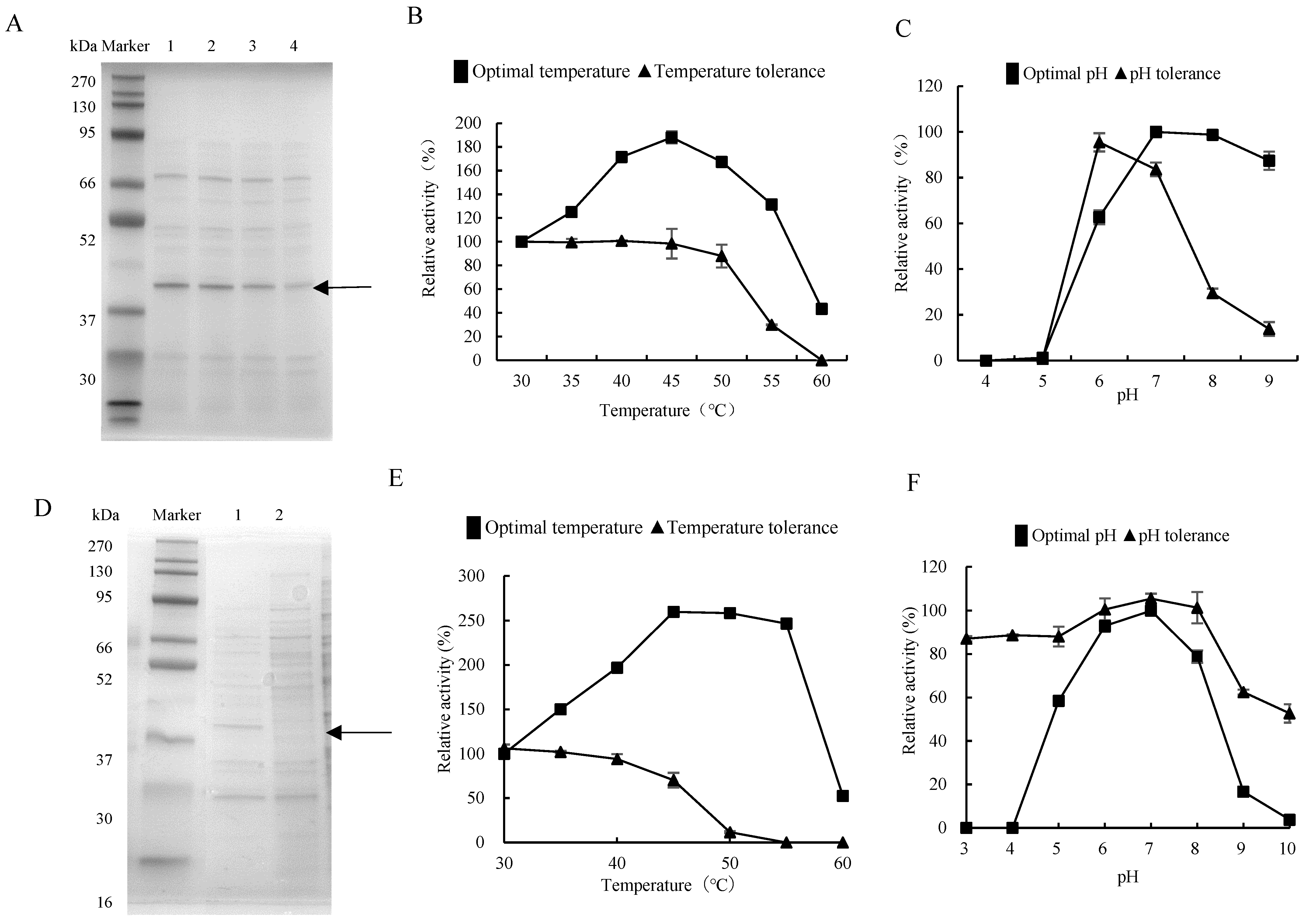
3.2. Exploration of Protease from Gut Bacteria
3.3. Homology Modeling and Binding Pocket Analysis of the DX-3-htpX Protease
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Douglas, A.E. The microbial dimension in insect nutritional ecology. Funct. Ecol. 2009, 23, 38–47. [Google Scholar] [CrossRef]
- Engel, P.; Moran, N.A. The gut microbiota of insects-diversity in structure and function. FEMS Microbiol. Rev. 2013, 37, 699–735. [Google Scholar] [CrossRef]
- Iyeh, C.I.; Ademolu, K.O.; Osipitan, A.A.; Idowu, A.B. Circadian Variations in the Digestive Enzyme Activities of Gut Homogenates of Adult American Cockroach, Periplaneta americana. Afr. Entomol. 2021, 29, 317–323. [Google Scholar] [CrossRef]
- Perera, O.P.; Shelby, K.S.; Pierce, C.A.; Snodgrass, G.L. Expression Profiles of Digestive Genes in the Gut and Salivary Glands of Tarnished Plant Bug (Hemiptera: Miridae). J. Insect Sci. 2021, 21, 3. [Google Scholar] [CrossRef]
- Shelomi, M.; Wipfler, B.; Zhou, X.; Pauchet, Y. Multifunctional cellulase enzymes are ancestral in Polyneoptera. Insect Mol. Biol. 2020, 29, 124–135. [Google Scholar] [CrossRef] [PubMed]
- Grover, S.; Kaur, S.; Gupta, A.K.; Taggar, G.K.; Kaur, J. Characterization of Trypsin Like Protease from Helicoverpa armigera (Hubner) and Its Potential Inhibitors. Proc. Natl. Acad. Sci. India B 2016, 88, 49–56. [Google Scholar] [CrossRef]
- MsangoSoko, K.; Gandotra, S.; Bhattacharya, R.; Ramakrishnan, B.; Sharma, K.; Subramanian, S. Screening and characterization of lipase producing bacteria isolated from the gut of a lepidopteran larvae, Samia ricini. J. Asia-Pac. Entomol. 2022, 25, 101856. [Google Scholar] [CrossRef]
- Kannan, M.; Mubarakali, D.; Thiyonila, B.; Krishnan, M.; Padmanaban, B.; Shantkriti, S. Insect gut as a bioresource for potential enzymes-an unexploited area for industrial biotechnology. Biocatal. Agric. Biotechnol. 2019, 18, 101010. [Google Scholar] [CrossRef]
- Hori, C.; Song, R.; Matsumoto, K.; Matsumoto, R.; Minkoff, B.B.; Oita, S.; Hara, H.; Takasuka, T.E. Proteomic Characterization of Lignocellulolytic Enzymes Secreted by the Insect-Associated Fungus Daldinia decipiens oita, Isolated from a Forest in Northern Japan. Appl. Environ. Microbiol. 2020, 86, e02350-19. [Google Scholar] [CrossRef] [PubMed]
- Sanatan, P.T.; Lomate, P.R.; Giri, A.P.; Hivrale, V.K. Characterization of a chemostable serine alkaline protease from Periplaneta americana. BMC Biochem. 2013, 14, 32. [Google Scholar] [CrossRef]
- Geib, S.M.; Tien, M.; Hoover, K. Identification of proteins involved in lignocellulose degradation using in gel zymogram analysis combined with mass spectroscopy-based peptide analysis of gut proteins from larval Asian longhorned beetles, Anoplophora glabripennis. Insect Sci. 2010, 17, 253–264. [Google Scholar] [CrossRef]
- Javaheri-Kermani, M.; Asoodeh, A. A novel beta-1,4 glucanase produced by symbiotic Bacillus sp. CF96 isolated from termite (Anacanthotermes). Int. J. Biol. Macromol. 2019, 131, 752–759. [Google Scholar] [CrossRef]
- Hatefi, A.; Makhdoumi, A.; Asoodeh, A.; Mirshamsi, O. Characterization of a bi-functional cellulase produced by a gut bacterial resident of Rosaceae branch borer beetle, Osphranteria coerulescens (Coleoptera: Cerambycidae). Int. J. Biol. Macromol. 2017, 103, 158–164. [Google Scholar] [CrossRef] [PubMed]
- Vilcinskas, A.; Schwabe, M.; Brinkrolf, K.; Plarre, R.; Wielsch, N.; Vogel, H. Larvae of the Clothing Moth Tineola bisselliella Maintain Gut Bacteria that Secrete Enzyme Cocktails to Facilitate the Digestion of Keratin. Microorganisms 2020, 8, 1415. [Google Scholar] [CrossRef] [PubMed]
- Sun, J.; Li, P.; Liu, Z.; Huang, W.; Mao, X. A novel thermostable serine protease from a metagenomic library derived from marine sediments in the East China Sea. Appl. Microbiol. Biotechnol. 2020, 104, 9229–9238. [Google Scholar] [CrossRef] [PubMed]
- Kannan, M.; Ramya, T.; Anbalagan, S.; Suriya, J.; Krishnan, M. Proteomic analysis of pupal gut serine protease of Silkworm, Bombyx mori: Partial purification and biochemical characterization. Biocatal. Agric. Biotechnol. 2017, 12, 159–165. [Google Scholar] [CrossRef]
- Banerjee, S.; Maiti, T.K.; Roy, R.N. Enzyme producing insect gut microbes: An unexplored biotechnological aspect. Crit. Rev. Biotechnol. 2021, 42, 384–402. [Google Scholar] [CrossRef]
- Chauhan, J.V.; Mathukiya, R.P.; Singh, S.P.; Gohel, S.D. Two steps purification, biochemical characterization, thermodynamics and structure elucidation of thermostable alkaline serine protease from Nocardiopsis alba strain OM-5. Int. J. Biol. Macromol. 2021, 169, 39–50. [Google Scholar] [CrossRef]
- Wilkesman, J. Cysteine Protease Zymography: Brief Review. Methods Mol. Biol. 2017, 1626, 25–31. [Google Scholar]
- Budatha, M.; Meur, G.; Dutta-Gupta, A. Identification and characterization of midgut proteases in Achaea janata and their implications. Biotechnol. Lett. 2008, 30, 305–310. [Google Scholar] [CrossRef]
- Ralhan, R.; Masui, O.; DeSouza, L.V.; Matta, A.; Macha, M.; Siu, K.W.M. Identification of proteins secreted by head and neck cancer cell lines using LC-MS/MS: Strategy for discovery of candidate serological biomarkers. Proteomics 2011, 11, 2363–2376. [Google Scholar] [CrossRef] [PubMed]
- Liu, Q.; Zhang, T.; Song, N.; Li, Q.; Wang, Z.; Zhang, X.; Lu, X.; Fang, J.; Chen, J. Purification and characterization of four key enzymes from a feather-degrading Bacillus subtilis from the gut of tarantula Chilobrachys guangxiensis. Int. Biodeterior. Biodegrad. 2014, 96, 26–32. [Google Scholar] [CrossRef]
- Paysan-Lafosse, T.; Blum, M.; Chuguransky, S.; Grego, T.; Pinto, B.L.; Salazar, G.A.; Bileschi, M.L.; Bork, P.; Bridge, A.; Colwell, L.; et al. InterPro in 2022. Nucleic Acids Res. 2023, 51, D418–D427. [Google Scholar] [CrossRef]
- Abramson, J.; Adler, J.; Dunger, J.; Evans, R.; Green, T.; Pritzel, A.; Ronneberger, O.; Willmore, L.; Ballard, A.J.; Bambrick, J.; et al. Accurate structure prediction of biomolecular interactions with AlphaFold 3. Nature 2024, 630, 493–500. [Google Scholar] [CrossRef]
- Ye, B.; Tian, W.; Wang, B.; Liang, J. CASTpFold: Computed Atlas of Surface Topography of the universe of protein Folds. Nucleic Acids Res. 2024, 52, W194–W199. [Google Scholar] [CrossRef] [PubMed]
- Ren, H.N.; Zhuo, T.X.; Bai, S.J.; Bai, Y.; Sun, X.Y.; Dan Liu, R.; Long, S.R.; Cui, J.; Wang, Z.Q. Proteomic analysis of hydrolytic proteases in excretory/secretory proteinsff from Trichinella spiralis intestinal infective larvae using zymography combined with shotgun LC-MS/MS approach. Acta Trop. 2021, 216, 105825. [Google Scholar] [CrossRef] [PubMed]
- Thanh Ha, D.T.; Kim Thoa, L.T.; Phuong Thao, T.T.; Dung, T.T.; Minh Ha, T.T.; Phuong Lan, T.T.; Khoo, K.S.; Show, P.L.; Huy, N.D. Production of extracellular agarase from Priestia megaterium AT7 and evaluation on marine algae hydrolysis. Enzym. Microb. Technol. 2024, 172, 110339. [Google Scholar] [CrossRef]
- Adlakha, N.; Rajagopal, R.; Kumar, S.; Reddy, V.S.; Yazdani, S.S. Synthesis and characterization of chimeric proteins based on cellulase and xylanase from an insect gut bacterium. Appl. Environ. Microbiol. 2011, 77, 4859–4866. [Google Scholar] [CrossRef]
- Petkauskaite, R.; Lukosius, D.; Dębski, J.; Jasilionis, A.; Dadlez, M.; Kieraite, I.; Timonina, A.; Kuisiene, N. Identification of proteins involved in starch and polygalacturonic acid degradation using LC/MS. Open Life Sci. 2014, 9, 708–716. [Google Scholar] [CrossRef]
- Robinson, S.L.; Piel, J.; Sunagawa, S. A roadmap for metagenomic enzyme discovery. Nat. Prod. Rep. 2021, 38, 1994–2023. [Google Scholar] [CrossRef]
- Munjal, N.; Jawed, K.; Wajid, S.; Yazdani, S.S. A constitutive expression system for cellulase secretion in Escherichia coli and its use in bioethanol production. PLoS ONE 2015, 10, e0119917. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
{kind=link}
| Gut Protease | Effective Protease Inhibitors | Protease Prediction |
|---|---|---|
| LGDBM-1 | Pepsin inhibitor, PMSF, trypsin inhibitor | Pepsin, serine protease, trypsin |
| LGDBM-2 | PMSF | Serine protease |
| LGDBM-3 | Trypsin inhibitor | Trypsin |
| LGDBM-4 | E-64, pepsin inhibitor, PMSF, trypsin inhibitor | Cysteine protease, pepsin, serine protease, trypsin |
| LGDBM-5 | - | - |
| LGDBM-6 | - | - |
| Gut Protease | Gene Names | Protein Names | Accession No. | Protein Identification/% | Score |
|---|---|---|---|---|---|
| LGDBM-1 | Glutathione S-transferase | P08515 | 35 | 141 | |
| DCD | Dermcidin | P81605 | 19 | 110 | |
| PSMA6 | Proteasome subunit alpha type-6 | P28072 | 7 | 74 | |
| LGDBM-3 | DCD | Dermcidin | P81605 | 12 | 92 |
| FTSH8 | ATP-dependent zinc metalloprotease | Q8W585 | 4 | 57 | |
| Glutathione S-transferase | P08515 | 12 | 46 | ||
| GGCT | Gamma-glutamyl cyclotransferase | Q32LE4 | 5 | 33 | |
| LGDBM-4 | Glutathione S-transferase | P08515 | 20 | 75 | |
| DCD | Dermcidin | P81605 | 12 | 74 | |
| groEL | Chaperonin GroEL | Q0KDR7 | 5 | 69 | |
| CTSD | Cathepsin D | Q4LAL9 | 2 | 27 | |
| LGDBM-5 | DCD | Dermcidin | P81605 | 12 | 74 |
| Glutathione S-transferase | P08515 | 26 | 66 | ||
| groEL | Chaperonin GroEL 1 | A0K536 | 2 | 23 | |
| LGDBM-6 | Glutathione S-transferase | P08515 | 26 | 110 | |
| DCD | Dermcidin | P81605 | 12 | 85 |
| Protease | Area (Å2) | Volume (Å3) | Active Sites in the D3 Pocket |
|---|---|---|---|
| HtpX | 557.472 | 837.241 | ARG4, LEU7, PHE8, VAL11, ALA52, SER55, LEU56, SER59, MET62, ALA63, TRP65, MET66, MET67, ASN113, ALA114, PHE115, ALA116, THR117, GLY118, MET132, VAL144, HIS147, GLU148, HIS151, MET157, THR160, THR161, LEU162, GLN164, ILE214, HIS217, SER218, ARG221, GLU222, MET238, ALA241, LEU242, LEU254, THR276, HIS277, ARG283 |
| HtpX-Ca2+ | 918.154 | 1378.221 | LEU1, LYS3, ALA110, GLU111, VAL112, ASN113, ALA114, MET132, VAL144, HIS147, GLU148, ARG219, MET238, ALA241, LEU245, ARG246, THR248, THR249, SER250, VAL252, ASP253, GLN256, LYS257, ALA260, LYS263, ILE264, SER265, LYS267, GLU268, PHE270, SER271, ARG272, PHE274, SER275, HIS277, PRO278, PRO279, LEU280 |
| HtpX-Cl− | 714.286 | 867.364 | LEU1, LYS3, GLU111, VAL112, ASN113, ALA114, MET132, VAL144, HIS147, GLU148, ARG219, MET238, ALA241, LEU242, LEU245, ARG246, THR248, THR249, SER250, VAL252, ASP253, GLN256, LYS257, ALA260, LYS263, ILE264, SER265, LYS267, GLU268, SER271, ARG272, SER275, HIS277, PRO278, PRO279, LEU280, GLU281, LYS282, ARG283 |
| HtpX-K+ | 925.544 | 1335.237 | LEU1, LEU2, LYS3, ALA110, GLU111, VAL112, ASN113, ALA114, MET132, VAL144, HIS147, GLU148, ARG219, MET238, ALA241, LEU242, LEU245, ARG246, THR248, THR249, SER250, VAL252, ASP253, GLN256, LYS257, ALA260, LYS263, ILE264, SER265, LYS267, GLU268, PHE270, SER271, ARG272, PHE274, SER275, HIS277, PRO278, PRO279, LEU280, GLU281, LYS282, ARG283 |
| HtpX-Zn2+ | 811.023 | 1179.127 | LEU1, LEU2, LYS3, ALA110, GLU111, VAL112, ASN113, ALA114, MET132, VAL144, HIS147, GLU148, ARG219, MET238, ALA241, LEU242, LEU245, ARG246, THR248, THR249, VAL252, ASP253, GLN256, LYS257, ALA260, LYS263, ILE264, SER265, LYS267, GLU268, SER271, ARG272, SER275, HIS277, PRO278, PRO279, LEU280, GLU281, ARG283 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zheng, X.; Wu, F.; Zhao, L.; Zhou, H.; Zhou, Z.; Jia, Z.; Shi, F. Exploration of Protease Resources in the Gut of Omnivorous Gryllotalpa orientalis (Orthoptera: Gryllotalpidae). Biology 2024, 13, 650. https://doi.org/10.3390/biology13090650
Zheng X, Wu F, Zhao L, Zhou H, Zhou Z, Jia Z, Shi F. Exploration of Protease Resources in the Gut of Omnivorous Gryllotalpa orientalis (Orthoptera: Gryllotalpidae). Biology. 2024; 13(9):650. https://doi.org/10.3390/biology13090650
Chicago/Turabian StyleZheng, Xiang, Fangtong Wu, Lu Zhao, He Zhou, Zhijun Zhou, Zhenhua Jia, and Fuming Shi. 2024. "Exploration of Protease Resources in the Gut of Omnivorous Gryllotalpa orientalis (Orthoptera: Gryllotalpidae)" Biology 13, no. 9: 650. https://doi.org/10.3390/biology13090650