
Pepino (Solanum muricatum) Metabolic Profiles and Soil Nutrient Association Analysis in Three Growing Sites on the Loess Plateau of Northwestern China


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
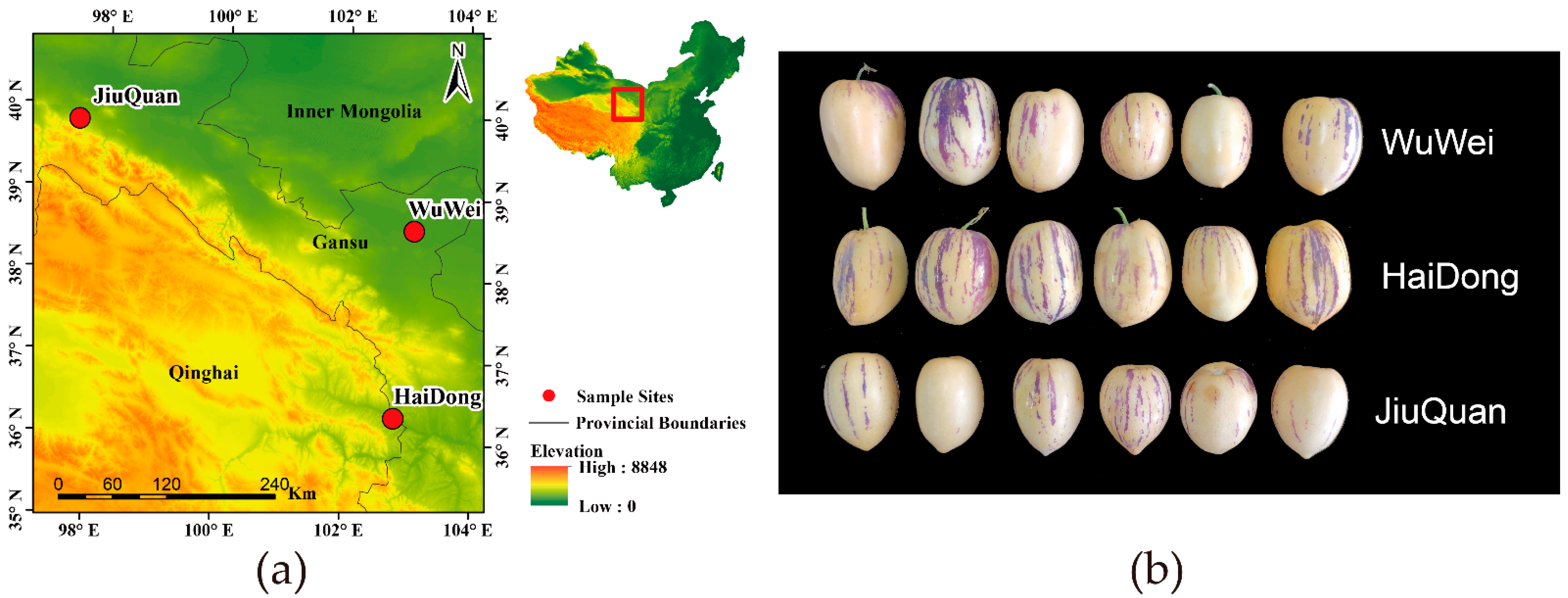
2.1. Sample Collection and Preparation
2.2. Metabolomic Analysis
2.2.1. Sample Processing
2.2.2. UPLC-Q-TOF/MS
2.3. Metabolomic Data Analysis
2.4. Soil Nutrients (NPK) Measurements in the Soils of the Three Regions
2.5. Statistical Analysis
3. Results
3.1. Metabolite Analysis
3.2. Differential Metabolite Analysis
3.3. Soil Nutrient Characteristics in the Three Planting Sites
3.4. Correlation Analysis of Soil Properties and Differential Metabolites
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Prohens, J.; Ruiz, J.J.; Nuez, F. The Pepino (Solanum Muricatum, Solanaceae): A “New” Crop with a History. Econ. Bot. 1996, 50, 355–368. [Google Scholar] [CrossRef]
- Anderson, G.J.; Jansen, R.K.; Kim, Y. The Origin and Relationships of the Pepino, Solanum Muricatum (Solanaceae): DNA Restriction Fragment Evidence. Econ. Bot. 1996, 50, 369–380. [Google Scholar] [CrossRef]
- Herraiz, F.J.; Blanca, J.; Ziarsolo, P.; Gramazio, P.; Plazas, M.; Anderson, G.J.; Prohens, J.; Vilanova, S. The First de Novo Transcriptome of Pepino (Solanum Muricatum): Assembly, Comprehensive Analysis and Comparison with the Closely Related Species S. Caripense, Potato and Tomato. BMC Genom. 2016, 17, 321. [Google Scholar] [CrossRef]
- Schaffer, A.A.; Rylski, I.; Fogelman, M. Carbohydrate Content and Sucrose Metabolism in Developing Solanum Muricatum Fruits. Phytochemistry 1989, 28, 737–739. [Google Scholar] [CrossRef]
- Rana, M.K.; Brar, A. Pepino. In Vegetable Crops Science; CRC Press: Boca Raton, FL, USA, 2017; pp. 901–914. ISBN 1-315-11620-0. [Google Scholar]
- Sudha, G.; Priya, M.S.; Shree, R.I.; Vadivukkarasi, S. In Vitro Free Radical Scavenging Activity of Raw Pepino Fruit (Solanum Muricatum Aiton). Int. J. Curr. Pharm. Res. 2011, 3, 137–140. [Google Scholar]
- Sudha, G.; Sangeetha Priya, M.; Shree, R.B.I.; Vadivukkarasi, S. Antioxidant Activity of Ripe and Unripe Pepino Fruit (Solanum Muricatum Aiton). J. Food Sci. 2012, 77, C1131–C1135. [Google Scholar] [CrossRef]
- Mahato, S.K.; Gurung, S.; Chakravarty, S.; Chhetri, B.; Khawas, T. An Introduction to Pepino (Solanum Muricatum Aiton). Int. J. Environ. Agric. Biotechnol. 2016, 1, 238513. [Google Scholar]
- Bravo, A.; Arias, E. Artichoke Cultivation Present Status and Prospects [Cynara Scolymus]. Campesino 1983, 114, 17–32. (In Spanish) [Google Scholar]
- Levy, D.; Kedar, N.; Levy, N. Pepino (Solanum Muricatum Aiton): Breeding in Israel for Better Taste and Aroma. Isr. J. Plant Sci. 2006, 54, 205–213. [Google Scholar] [CrossRef]
- Rodríguez-Burruezo, A.; Prohens, J.; Nuez, F. ‘Valencia’: A New Pepino (Solanum Muricatum) Cultivar with Improved Fruit Quality. HortScience 2004, 39, 1500–1502. [Google Scholar] [CrossRef]
- Ruiz, J.J.; Prohens, J.; Nuez, F. ‘Sweet Round’ and ‘Sweet Long’: Two Pepino Cultivars for Mediterranean, Climates. HortScience 1997, 32, 751–752. [Google Scholar] [CrossRef]
- Ge, B.; He, Z.; Zhang, Z.; Wang, H.; Li, S. Genetic Variation in Potato Virus M Isolates Infecting Pepino (Solanum Muricatum) in China. Arch. Virol. 2014, 159, 3197–3210. [Google Scholar] [CrossRef]
- She, X.; Yu, L.; Lan, G.; Tang, Y.; Deng, M.; Li, Z.; He, Z. Pantoea Agglomerans Causing Blight Disease on Pepino Melon (Solanum Muricatum) in China. Crop Prot. 2021, 139, 105385. [Google Scholar] [CrossRef]
- Hu, J.; Huang, W.; Zhang, F.; Luo, X.; Chen, Y.; Xie, J. Variability of Volatile Compounds in the Medicinal Plant Dendrobium Officinale from Different Regions. Molecules 2020, 25, 5046. [Google Scholar] [CrossRef]
- Wang, C.; Lv, S.; Wu, Y.; Gao, X.; Li, J.; Zhang, W.; Meng, Q. Oolong Tea Made from Tea Plants from Different Locations in Yunnan and Fujian, China Showed Similar Aroma but Different Taste Characteristics. SpringerPlus 2016, 5, 576. [Google Scholar] [CrossRef]
- Zhang, X.; Zhao, Y.; Guo, L.; Qiu, Z.; Huang, L.; Qu, X. Differences in Chemical Constituents of Artemisia Annua L from Different Geographical Regions in China. PLoS ONE 2017, 12, e0183047. [Google Scholar] [CrossRef]
- Davies, J.N.; Hobson, G.E.; McGlasson, W.B. The Constituents of Tomato Fruit—The Influence of Environment, Nutrition, and Genotype. Crit. Rev. Food Sci. Nutr. 1981, 15, 205–280. [Google Scholar] [CrossRef]
- Wert, T.W.; Williamson, J.G.; Chaparro, J.X.; Miller, E.P.; Rouse, R.E. The Influence of Climate on Fruit Development and Quality of Four Low-Chill Peach Cultivars. HortScience 2009, 44, 666–670. [Google Scholar] [CrossRef]
- Park, B.S.; Song, J.T.; Seo, H.S. Arabidopsis Nitrate Reductase Activity Is Stimulated by the E3 SUMO Ligase AtSIZ1. Nat. Commun. 2011, 2, 400. [Google Scholar] [CrossRef]
- Mao, Y.-B.; Liu, Y.-Q.; Chen, D.-Y.; Chen, F.-Y.; Fang, X.; Hong, G.-J.; Wang, L.-J.; Wang, J.-W.; Chen, X.-Y. Jasmonate Response Decay and Defense Metabolite Accumulation Contributes to Age-Regulated Dynamics of Plant Insect Resistance. Nat. Commun. 2017, 8, 13925. [Google Scholar] [CrossRef]
- Shi, Z.; Wei, F.; Wan, R.; Li, Y.; Wang, Y.; An, W.; Qin, K.; Dai, G.; Cao, Y.; Feng, J. Impact of Nitrogen Fertilizer Levels on Metabolite Profiling of the Lycium Barbarum L. Fruit. Molecules 2019, 24, 3879. [Google Scholar] [CrossRef]
- Mutua, C.M.; Ogweno, J.O.; Gesimba, R.M. Effect of NPK Fertilizer Rates on Growth and Yield of Field and Greenhouse Grown Pepino Melon (Solanum Muricatum Aiton). J. Hortic. Plant Res. 2021, 13, 10–23. [Google Scholar] [CrossRef]
- Mutua, C.M.; Ogweno, J.O.; Gesimba, R.M. Effect of NPK Fertilizer Rates on Secondary Metabolites of Pepino Melon (Solanum Muricatum Aiton). J. Hortic. For. 2021, 13, 25–34. [Google Scholar]
- Li, D.; Gaquerel, E. Next-Generation Mass Spectrometry Metabolomics Revives the Functional Analysis of Plant Metabolic Diversity. Annu. Rev. Plant Biol. 2021, 72, 867–891. [Google Scholar] [CrossRef]
- Jacobs, D.M.; Van Den Berg, M.A.; Hall, R.D. Towards Superior Plant-Based Foods Using Metabolomics. Curr. Opin. Biotechnol. 2021, 70, 23–28. [Google Scholar] [CrossRef]
- Fuentes, S.; Tongson, E.; Viejo, C.G. Novel Digital Technologies Implemented in Sensory Science and Consumer Perception. Curr. Opin. Food Sci. 2021, 41, 99–106. [Google Scholar] [CrossRef]
- Tsugawa, H.; Cajka, T.; Kind, T.; Ma, Y.; Higgins, B.; Ikeda, K.; Kanazawa, M.; VanderGheynst, J.; Fiehn, O.; Arita, M. MS-DIAL: Data-Independent MS/MS Deconvolution for Comprehensive Metabolome Analysis. Nat. Methods 2015, 12, 523–526. [Google Scholar] [CrossRef]
- Horai, H.; Arita, M.; Kanaya, S.; Nihei, Y.; Ikeda, T.; Suwa, K.; Ojima, Y.; Tanaka, K.; Tanaka, S.; Aoshima, K. MassBank: A Public Repository for Sharing Mass Spectral Data for Life Sciences. J. Mass Spectrom. 2010, 45, 703–714. [Google Scholar] [CrossRef]
- Van Lagen, B. Soil Analyses. In Manual for Soil and Water Analysis; Backhuys Publishers: Leiden, The Netherlands, 1996; pp. 1–20. [Google Scholar]
- Douglas, L.A.; Riazi, A.; Smith, C.J. A Semi-micro Method for Determining Total Nitrogen in Soils and Plant Material Containing Nitrite and Nitrate. Soil Sci. Soc. Am. J. 1980, 44, 431–433. [Google Scholar] [CrossRef]
- Yang, Z.; Zhang, Y.; Wang, Y.; Zhang, H.; Zhu, Q.; Yan, B.; Fei, J.; Xiangmin, R.; Peng, J.; Luo, G. Intercropping Regulation of Soil Phosphorus Composition and Microbially-Driven Dynamics Facilitates Maize Phosphorus Uptake and Productivity Improvement. Field Crops Res. 2022, 287, 108666. [Google Scholar] [CrossRef]
- Öborn, I.; Edwards, A.C.; Hillier, S. Quantifying Uptake Rate of Potassium from Soil in a Long-Term Grass Rotation Experiment. Plant Soil 2010, 335, 3–19. [Google Scholar] [CrossRef]
- Zhao, W.; Zhou, Q.; Tian, Z.; Cui, Y.; Liang, Y.; Wang, H. Apply Biochar to Ameliorate Soda Saline-Alkali Land, Improve Soil Function and Increase Corn Nutrient Availability in the Songnen Plain. Sci. Total Environ. 2020, 722, 137428. [Google Scholar] [CrossRef]
- Tiessen, H.; Moir, J.O. Characterization of Available P by Sequential Extraction. Soil Sampl. Methods Anal. 1993, 7, 5–229. [Google Scholar]
- Xingcui, X.; Rongxiu, X.I.E.; Zhihui, L.I.; Weijing, K.; Jiyou, W.U. Effects of Fertilization on Growth and Soil of Middle-Aged Slash Pine Forest. Agric. Sci. Technol. 2015, 16, 2386–2393. [Google Scholar]
- Berbecea, A.A.; Radulov, I.; Sala, F.; Crista, F.; Lato, A. Interrelation between Metal Availability, Soil PH and Mineral Fertilization. Res. J. Agric. Sci. 2011, 43, 19–22. [Google Scholar]
- Pang, Z.; Chong, J.; Zhou, G.; de Lima Morais, D.A.; Chang, L.; Barrette, M.; Gauthier, C.; Jacques, P.-É.; Li, S.; Xia, J. MetaboAnalyst 5.0: Narrowing the Gap between Raw Spectra and Functional Insights. Nucleic Acids Res. 2021, 49, W388–W396. [Google Scholar] [CrossRef]
- Kola, O. Physical and Chemical Characteristics of the Ripe Pepino (Solanum Muricatum) Fruit Grown in Turkey. J. Food Agric. Environ. 2010, 8, 168–171. [Google Scholar]
- Luykx, D.M.; Van Ruth, S.M. An Overview of Analytical Methods for Determining the Geographical Origin of Food Products. Food Chem. 2008, 107, 897–911. [Google Scholar] [CrossRef]
- Becerra-Martínez, E.; Florentino-Ramos, E.; Pérez-Hernández, N.; Zepeda-Vallejo, L.G.; Villa-Ruano, N.; Velázquez-Ponce, M.; García-Mendoza, F.; Bañuelos-Hernández, A.E. 1H NMR-Based Metabolomic Fingerprinting to Determine Metabolite Levels in Serrano Peppers (Capsicum Annum L.) Grown in Two Different Regions. Food Res. Int. 2017, 102, 163–170. [Google Scholar] [CrossRef]
- Mutua, C.; Ogweno, J.; Gesimba, R. Postharvest Quality of Pepino Melon (Solanum Muricatum Aiton) as Influenced by NPK Fertilizer Rates, Growing Environment and Storage Temperature. Adv. Hortic. Sci. 2021, 35, 1–96. [Google Scholar] [CrossRef]
- Kola, O.; Simsek, M.; Duran, H.; Bozkir, H. HPLC Determination of Carotenoid, Organic Acid, and Sugar Content in Pepino (Solanum Muricatum) Fruit during the Ripening Period. Chem. Nat. Compd. 2015, 51, 132–136. [Google Scholar] [CrossRef]
- Redgwell, R.J.; Turner, N.A. Pepino (Solanum Muricatum): Chemical Composition of Ripe Fruit. J. Sci. Food Agric. 1986, 37, 1217–1222. [Google Scholar] [CrossRef]
- Contreras, C.; Schwab, W.; Mayershofer, M.; Morales, I.; González-Agüero, M.; Defilippi, B. Study of Physiological and Quality Parameters during Development and Ripening of Pepino (Solanum Muricatum Aiton) Fruit. Chil. J. Agric. Res. 2019, 79, 385–395. [Google Scholar] [CrossRef]
- Huyskens-Keil, S.; Widayat, H.P.; Lüdders, P.; Schreiner, M.; Peters, P. Physiological Changes of Pepino (Solanum Muricatum Ait.) during Maturation and Ripening. In Proceedings of the II ISHS Conference on Fruit Production in the Tropics and Subtropics, Bonn-Röttgen, Germany, 24 June 1999; pp. 251–256. [Google Scholar]
- Ahumada, M.; Cantwell, M. Postharvest Studies on Pepino Dulce (Solanum Muricatum Ait.): Maturity at Harvest and Storage Behavior. Postharvest Biol. Technol. 1996, 7, 129–136. [Google Scholar] [CrossRef]
- Contreras, C.; González-Agüero, M.; Defilippi, B.G. A Review of Pepino (Solanum Muricatum Aiton) Fruit: A Quality Perspective. HortScience 2016, 51, 1127–1133. [Google Scholar] [CrossRef]
- Wang, Y.; Liang, X.; Li, Y.; Fan, Y.; Li, Y.; Cao, Y.; An, W.; Shi, Z.; Zhao, J.; Guo, S. Changes in Metabolome and Nutritional Quality of Lycium Barbarum Fruits from Three Typical Growing Areas of China as Revealed by Widely Targeted Metabolomics. Metabolites 2020, 10, 46. [Google Scholar] [CrossRef]
- Shi, Q.; Han, G.; Liu, Y.; Jiang, J.; Jia, Y.; Li, X. Nutrient Composition and Quality Traits of Dried Jujube Fruits in Seven Producing Areas Based on Metabolomics Analysis. Food Chem. 2022, 385, 132627. [Google Scholar] [CrossRef]
- Radi, M.; Mahrouz, M.; Jaouad, A.; Gaouji, A.; Tacchini, M.; Aubert, S.; Amiot, M.J. Effect of Mineral Fertilization (NPK) on the Quality of Apricot. In Proceedings of the III International Symposium on Mineral Nutrition of Deciduous Fruit Trees, Zaragoza, Spain, 27 May 1996; p. 508. [Google Scholar]
- Milošević, T.; Milošević, N.; Mladenović, J. The Influence of Organic, Organo-Mineral and Mineral Fertilizers on Tree Growth, Yielding, Fruit Quality and Leaf Nutrient Composition of Apple Cv. ‘Golden Delicious Reinders’. Sci. Hortic. 2022, 297, 110978. [Google Scholar] [CrossRef]
- Smoleń, S.; Sady, W. The Effect of Various Nitrogen Fertilization and Foliar Nutrition Regimes on the Concentrations of Sugars, Carotenoids and Phenolic Compounds in Carrot (Daucus Carota L.). Sci. Hortic. 2009, 120, 315–324. [Google Scholar] [CrossRef]
- Radi, M.; Mahrouz, M.; Jaouad, A.; Amiot, M. Influence of Mineral Fertilization (NPK) on the Quality of Apricot Fruit (Cv. Canino). The Effect of the Mode of Nitrogen Supply. Agronomie 2003, 23, 737–745. [Google Scholar] [CrossRef]
- Famiani, F.; Battistelli, A.; Moscatello, S.; Cruz-Castillo, J.G.; Walker, R.P. The Organic Acids That Are Accumulated in the Flesh of Fruits: Occurrence, Metabolism and Factors Affecting Their Contents—A Review. Rev. Chapingo Ser. Hortic. 2015, 21, 97–128. [Google Scholar] [CrossRef]
- Kyriacou, M.C.; Leskovar, D.I.; Colla, G.; Rouphael, Y. Watermelon and Melon Fruit Quality: The Genotypic and Agro-Environmental Factors Implicated. Sci. Hortic. 2018, 234, 393–408. [Google Scholar] [CrossRef]
- Minas, I.S.; Tanou, G.; Molassiotis, A. Environmental and Orchard Bases of Peach Fruit Quality. Sci. Hortic. 2018, 235, 307–322. [Google Scholar] [CrossRef]
- García-Gómez, B.E.; Salazar, J.A.; Nicolás-Almansa, M.; Razi, M.; Rubio, M.; Ruiz, D.; Martínez-Gómez, P. Molecular Bases of Fruit Quality in Prunus Species: An Integrated Genomic, Transcriptomic, and Metabolic Review with a Breeding Perspective. Int. J. Mol. Sci. 2020, 22, 333. [Google Scholar] [CrossRef]
- Çolak, N.G.; Eken, N.T.; Ülger, M.; Frary, A.; Doğanlar, S. Mapping of Quantitative Trait Loci for Antioxidant Molecules in Tomato Fruit: Carotenoids, Vitamins C and E, Glutathione and Phenolic Acids. Plant Sci. 2020, 292, 110393. [Google Scholar] [CrossRef]









Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sun, Z.; Wang, L.; Zhang, G.; Yang, S.; Zhong, Q. Pepino (Solanum muricatum) Metabolic Profiles and Soil Nutrient Association Analysis in Three Growing Sites on the Loess Plateau of Northwestern China. Metabolites 2022, 12, 885. https://doi.org/10.3390/metabo12100885
Sun Z, Wang L, Zhang G, Yang S, Zhong Q. Pepino (Solanum muricatum) Metabolic Profiles and Soil Nutrient Association Analysis in Three Growing Sites on the Loess Plateau of Northwestern China. Metabolites. 2022; 12(10):885. https://doi.org/10.3390/metabo12100885
Chicago/Turabian StyleSun, Zhu, Lihui Wang, Guangnan Zhang, Shipeng Yang, and Qiwen Zhong. 2022. "Pepino (Solanum muricatum) Metabolic Profiles and Soil Nutrient Association Analysis in Three Growing Sites on the Loess Plateau of Northwestern China" Metabolites 12, no. 10: 885. https://doi.org/10.3390/metabo12100885