Epicardial Lineages

Institut für Molekularbiologie, OE5250, Medizinische Hochschule Hannover, Carl-Neuberg-Str. 1, D-30625 Hannover, Germany
*
Author to whom correspondence should be addressed.
J. Dev. Biol. 2013, 1(1), 32-46; https://doi.org/10.3390/jdb1010032
Submission received: 16 May 2013
/
Revised: 10 June 2013
/
Accepted: 17 June 2013
/
Published: 21 June 2013
Abstract
:The epicardium is the mono-layered epithelium that covers the outer surface of the myocardium from early in cardiac development. Long thought to act merely passively to protect the myocardium from frictional forces in the pericardial cavity during the enduring contraction and expansion cycles of the heart, it is now considered to be a crucial source of cells and signals that direct myocardial growth and formation of the coronary vasculature during development and regeneration. Lineage tracing efforts in the chick, the mouse and the zebrafish unambiguously identified fibroblasts in interstitial and perivascular locations as well as coronary smooth muscle cells as the two major lineages that derive from epithelial-mesenchymal transition and subsequent differentiation from individual epicardial cells. However, controversies exist about an additional endothelial and myocardial fate of epicardial progenitor cells. Here, we review epicardial fate mapping efforts in three vertebrate model systems, describe their conceptual differences and discuss their methodological limitations to reach a consensus of the potential of (pro-)epicardial cells in vitro and in vivo.
1. The Epicardium—A Cardiac Tissue of Extracardiac Origin
The vertebrate heart with its chamber organization and three-layered tissue design is the result of a complex morphogenetic program that progressively transforms a simple organ rudiment into a large muscular pump. Endocardium and myocardium, i.e. the inner and the muscular layer of the heart, are present in the simple peristaltically active tube that is established shortly after gastrulation to support the need of the rapidly growing embryo. The epicardium, the outer tissue layer, is only added once this simple tube has undergone a complicated looping process and established the chambers.
The myocardium and endocardium arise from fusion of two bilateral fields of cardiac precursor cells in the anterior lateral plate mesoderm (the first heart field), and subsequent recruitment and delayed myocardial differentiation of pharyngeal mesoderm (the second heart field) [1]. The epicardium, in contrast, derives from an outgrowth of the coelomic epithelium that is established posterior to the growing heart tube from a subpool of the anterior lateral plate mesoderm [2,3] at embryonic day (E) 8.5 in the mouse [4,5], at Hamburger and Hamilton stage (HH) 14 in the chick, and 48 h post fertilization (hpf) in the zebrafish [3]. This mesothelial outgrowth, which gains a cauliflower-like shape, was coined proepicardial organ, or more frequently, proepicardium. The proepicardium is an unpaired midline structure, but it forms bilaterally in close proximity to the two wings of the sinus venosus myocardium. The left proepicardial anlage regresses during further development in the chick, whereas both proepicardial anlagen merge in the midline of the mouse embryo [6].
While the epicardium is thought to arise exclusively from the proepicardium, the proepicardium most likely does not solely contribute to the epicardium but seems to be a mixed population of epicardial precursors and endothelial cells [7,8,9,10]. The close spatial arrangement of different precursor tissues including that of the liver, the septum transversum, and the myocardium next to the proepicardium makes the analysis of the lineage relationships at the posterior heart pole even more complicated. This, and the lack of a unique genetic label for a proepicardial character of cells may at least partly explain the discrepancies between the different lineage tracing approaches performed in the different vertebrate models. Nonetheless, the proepicardium, is a transient embryonic tissue. Proepicardial cells are transferred to the myocardium via an extracellular matrix bridge in avian embryos [11] and by finger-like protrusions contacting the heart as well as proepicardial vesicles in the murine and zebrafish heart [5]. Once proepicardial cells attach to the myocardial surface of the looped heart, they proliferate and migrate to ensheath the heart down to the base of the outflow tract, at E10.5 in murine heart development (HH25 in the chick [12]). From this point in development, the epicardium supports myocardial growth by a cocktail of paracrine factors (for review see Olivey and Svensson 2010 [13]).
Shortly later at E11.5 to E12.5 (HH20 in the chick), the first cells start to leave the continuity of the mono-layered epithelial epicardium. This process progresses from the atrio-ventricular groove to the apex of the ventricles in parallel with the outgrowth of the coronary plexus. At around E15.5 (HH27 in the chick [14]) epicardial cells are no longer mobilized under developmental conditions, and are likely to differentiate terminally. The lineages to which epicardium-derived cells (EPDCs) [15] give rise have been analyzed in the development of chick, mouse and zebrafish embryos using a number of methods with a consensus on the main fates but with diverging views of additional minor fates.
2. Proepicardial Lineage Tracing in Chick Development—Fibroblasts, Coronary Smooth Muscle Cells and Possibly Endothelial Cells
The earliest study that analyzed the fate of proepicardial cells was performed by Mikawa and Gourdie in 1996. They used retroviral gene transfer to transduce a lacZ reporter gene into proepicardial cells both in ovo as well as in vitro with subsequent transplantation of a proepicardial cell cluster into isochronic recipients in the latter case. Using ß-galactosidase histochemistry on sections of E18 hearts they identified coronary smooth muscle cells and coronary endothelial cells as well as perivascular connective tissue as being proepicardium-derived, demonstrating for the first time that cells of the coronary vasculature (at least partly) derive from the proepicardium. Intriguingly, immunoconfocal microscopy identified both endothelial and smooth muscle cell marker gene expression within the proepicardium, suggesting that these lineages are already present within the proepicardium and that they migrate along with future epicardial cells onto the myocardium [16].
While the presence of endothelial cells in or at the proepicardium is likely (see also below), an immigration of differentiated smooth muscle cells from the proepicardium into the myocardium has not been confirmed. However, Perez-Pomares and co-workers identified delaminating cells between the epicardial and myocardial tissue layers [14], which are mesenchymal but lack differentiation markers of the smooth muscle and endothelial cell lineages [7,17]. Further, proepicardial explant cultures that were induced by angiogenic growth factors like fibroblast growth factor 2 (FGF2) and vascular endothelial growth factor (VEGF) become mesenchymal and invade collagen matrices [17]. Collectively, these results suggested that individual epicardial cells undergo an epithelial-to-mesenchymal conversion, and only subsequently differentiate further.
Fate mapping of quail epicardial cells (HH18) transduced with ß-galactosidase encoding adenoviruses after transplantation into HH15 chick recipients revealed the contribution of these epicardium-derived mesenchymal precursor cells to coronary smooth muscle cells, perivascular and intermyocardial fibroblasts, mesenchymal cells of the atrio-ventricular cushions and some ventricular endocardial cells [17]; similar results were obtained by quail to chick transplantation of proepicardia [15,18]. However, studies of these quail-chick chimeras additionally identified endothelial cells of the coronary vessels as proepicardium-derived [7,19]. An endothelial fate of epicardial cells is controversial as Poelmann and colleagues has provided evidence that endothelial cells found in proepicardial grafts are liver-derived [12] and Dettman and colleagues have shown that epicardial grafts do not contribute to the endothelial cell lineage of the avian heart [17]. Furthermore, immunohistochemical analysis of avian proepicardia and proepicardial explants indicate that endothelial cells are present within the proepicardium of quail and chick embryos [16] and their abundance varies with the age of the proepicardial graft [20]. In the end, we would like to point out the study by Männer in 1999 who investigated the contribution of epicardial cells to the myocardial lineage of the heart. He did not detect any quail-derived myocardial cells in quail-chick chimeras [18], which is in line with missing reports of epicardium-derived cardiomyocytes in avian embryos.
In conclusion, (pro-)epicardial fate mapping has revealed that embryonic epicardial cells are a multipotent precursor population. While most of them contribute to the mature epicardium, some become mesenchymal and differentiate into vascular lineages (coronary smooth muscle cells, perivascular fibroblasts), mesenchymal cells of the atrio-ventricular cushion tissue (mitral and triscupid valves), interstitial fibroblasts and in a minor fraction to endocardial and subendocardial cells in the chick embryo (see Table 1 for a summary). Endothelial cells of unknown origin exist in the proepicardium and may contribute to the coronary vasculature by co-migration with epicardial precursor cells. A differentiation of (pro-)epicardial cells into the endothelial cell lineage seems unlikely.
3. Epicardial Lineage Tracing in Mouse Development—An Epicardial Origin of Myocardial Cells?
As intrauterine development largely precludes or at least hinders the direct manipulation and labeling of the (pro-)epicardium in the mouse, lineage tracing of these cells has only become possible with the advent of suitable genetic labeling systems. Cre is a phage-encoded recombinase that specifically recognizes short loxP called DNA stretches to delete (if the two loxP sites are oriented in the same direction) or flip (if the loxP sites are reversed) the intervening sequences. If loxP sites are suitably placed around a reporter gene (such as GFP or lacZ), which has been introduced in a ubiquitously expressed locus such as Rosa26, a single cre mediated recombination event can irreversibly activate the reporter (e.g., by removing a loxP flanked stop cassette in front of the reporter) in all descendants of a founder cell. Hence, if cre is expressed under the control of an (pro-)epicardial-specific promoter, the fate of these cells can be analyzed at any time-point of development by a simple co-expression analysis of the reporter and the differentiation marker. Therefore, epicardial fate mapping in the mouse (and other species that allow efficient genetic manipulation such as the zebrafish) boils down to having a sensitive reporter line and a specific cre driver line at hand [21,22]. While the first task was quickly achieved, finding a suitable cre line turned out to be much more of a problem.
The first report on such a genetic fate mapping approach in the mouse came from Wilm and co-workers in 2005 who reported descendents of epicardial cells in the coronary vasculature (almost all smooth muscle cells, and a low degree of endothelial cells) based on a cre knock-in in a 280 kbp YAC of the Wilms tumor 1 (Wt1) locus [23]. Wt1 is a gene that is strongly expressed in the proepicardium and in cells of the coelomic lining including the epicardium [24]. Furthermore, a regulatory fragment of the chick Gata5 locus mediated cre expression in the mouse epicardium and identified cardiac fibroblasts and smooth muscle cells as epicardial descendants again confirming the data obtained in the chick [25].
Two additional independent lineage tracings by cre/loxP technology further confirmed fibroblasts and smooth muscle cells as epicardial fates and suggested, surprisingly, that epicardial cells albeit with different patterns also differentiate into cardiomyocytes in the mouse. The study by Cai and co-workers used a cre knock-in allele of T-box gene 18 (Tbx18) (Tbx18tm2(cre)Sev) for epicardial lineage tracing and observed that cardiomyocytes of the left ventricle and the interventricular septum were positive for reporter activity [26]. Lineage tracing using a creEGFP knock-in allele of Wt1 (Wt1tm1(EGFP/cre)Wtp) reported a substantial contribution of epicardial cells to the myocardium of all four cardiac chambers and in the interventricular septum, constituting 7–10% of cardiomyocytes in ventricles and 18% in atria starting from E10.5 [27]. As an epicardial origin of myocardial cells was incongruent with all previous fate-mapping efforts both in the chick and the mouse, the suitability of the cre lines was quickly challenged. Tbx18 is expressed in the proepicardium and its expression is maintained in epicardial cells until E16.5. However, Tbx18 is also expressed endogenously in cardiomyocytes of the interventricular septum and the left ventricle i.e., exactly in those regions which the study by Cai et al. suggested to derive from the epicardium [28]. Using an independent cre knock-in allele of Tbx18 (Tbx18tm4(cre)Akis) Grieskamp and colleagues showed that epicardial cells overlying the right ventricular myocardium do not differentiate into cardiomyocytes but give exclusively rise to smooth muscle cells and fibroblasts [29]. More recently, it has been shown that Wt1 is not specific for epicardial cells but is also expressed in coronary endothelial cells and possibly in cardiomyocytes [30,31]; an (pro-)epicardial origin of these cell types can therefore not be analyzed with Wt1-based cre lines. Independent of these limitations, it has recently been reported that Wt1tm1(EGFP/cre)Wtp mediates widespread ectopic expression in the early embryo including the heart, further questioning the results of earlier Wt1-based lineage tracing efforts [30,31]. However, similar to the chick heart, former Wt1-expressing EPDCs replace mesenchymal cells of the atrio-ventricular cushion tissue in a second invasion and contribute to mitral and tricuspid valves as well as to the annulus fibrosus [32,33].
Due to the mentioned limitations of cre lines based on Tbx18 and Wt1 regulatory elements for epicardial lineage tracing, several groups investigated the potential of other promoter elements to specifically label (pro-)epicardial cells. Transcription factor 21 (Tcf21) is expressed in the proepicardium, epicardium and a subset of epicardium-derived cells that contribute to the fibroblast lineage of murine hearts. Lineage tracing of Tcf21-expressing epicardial cells of E10.5 hearts with an inducible cre-driver line (Tcf21MerCreMer) revealed their contribution to coronary smooth muscle cells, interstitial and perivascular fibroblasts; Tcf21-expressing epicardial cells, however, did not account for coronary endothelial cells or cardiomyocytes [34].
In a recent study, Rinkevich and colleagues investigated the differentiation potential of mesothelial cells using a knock-in of a creERT2-IRES-LacZ cassette into the mesothelin locus (MslnCln), thereby providing a mesothelium-specific tool to trace the fate of epicardial cells. Although the study did not focus on the heart, the authors stated that they observed a restriction of descendants of Msln-expressing (i.e., mesothelial) cells to the fibroblast and smooth muscle lineage of the heart during embryogenesis as well as during postnatal life of mice. A contribution of the mesothelium to endothelial cells or cardiac muscle was not detected [35].
Katz and colleagues used cre lines based on regulatory elements of Semaphorin 3D (Sema3D) and Scleraxis (Scx) to further delineate the molecular and cellular complexity of the proepicardium. Both genes are initially expressed in the proepicardium but in a pattern only partially overlapping with that of Tbx18 and Wt1 and are maintained in the epicardium for a variable length of time; importantly, none of these genes is expressed in coronary endothelial cells. Lineage tracing of Sema3D-expressing (pro)epicardial cells (Sema3dtm1.1(GFP/cre)Cjt) revealed their contribution to coronary smooth muscle cells, interstitial fibroblasts, atrio-ventricular valves and coronary endothelial cells as well as a minor portion (0.36%) of cardiomyocytes. Endothelial cells derived from the Sema3D lineage appear in the sinus venosus and the coronary plexus growing into the heart after E11.5. These observations are in line with the derivation of coronary endothelial cells from the sinus venosus endothelium as described by Red-Horse and colleagues in 2010 [36]. However, it was known that endothelial cells that derive from the Scx-expressing proepicardial cell population (Tg(Scx-GFP/cre)1Stzr) emerged directly underneath the epicardium at E11, before the endothelial cells from the sinus venosus were detected by Red-Horse et al 2010. This finding indicates another route of proepicardial contribution to the endothelial cell lineage, possibly via endocardial cells [37]. Next to endothelial cells, Scx-expressing proepicardial cells also contribute to coronary smooth muscle cells and cardiomyocytes of the murine heart although in an unexplained pattern in the latter case.
In summary, genetic fate mappings showed that murine epicardial cells give rise to coronary smooth muscle cells, perivascular/interstitial fibroblasts and mesenchymal cells of the annulus fibrosus and atrio-ventricular valves [26,27,29,32,33]. Lineage-labeled cardiomyocytes have been observed in several studies, but their epicardial origin is controversial as myocardial recombination of the used cre-driver lines is likely [28,30]. Coronary endothelial cells are, as in the avian system, proepicardium-derived but their origin is distinct from the Tbx18-/Wt1-expressing epicardial cell population; further investigations are needed to determine their migration route. The fates of murine (pro-)epicardial cells are summarized in Table 1.
4. Epicardial Fates in Zebrafish Development—Fibroblasts and Smooth Muscle Cells and Nothing Else
The first researchers, that set out to investigate (pro-)epicardial fates in the zebrafish were Kikuchi and colleagues in 2011. Tbx18, wt1b and tcf21 were tested for epicardium-specific genetic lineage tracings. Both, tbx18 and wt1b regulatory elements did not drive epicardium-specific reporter gene expression, confirming the observations made in mice. The regulatory elements of tcf21, however, faithfully recapitulated specific epicardial tcf21 expression. (Pro-)epicardial cells of the tcf21-lineage (Tg(tcf21:DsRed2)pd37) contributed to the subepicardium and perivascular cells; lineage-labeled cardiomyocytes or endothelial cells were not identified. Tracing of epicardial cells of the zebrafish larva with an inducible tcf21-cre line (Tg(tcf21:CreER)pd42) revealed that epicardial cells contributed to the adult epicardium, the ventricular subepicardium but never to the trabeculated myocardium. Perivascular cells of larger vessels and smooth muscle cells of the bulbus arteriosus but not cardiomyocytes were labeled by this technique [38]. Hence, tcf21-expressing (pro-)epicardial cells give rise to subepicardial mesenchyme, perivascular cells and a few smooth muscle cells of the bulbus arteriosus in the zebrafish heart.
5. In Vitro Studies Reveal Proepicardial and Epicardial Potentials
Given the origin of the epicardium from the proepicardium and their shared molecular characteristics, it has turned out difficult to independently assess the fate of cells in both tissues. A possible way out of this dilemma is offered by explant cultures of proepicardium and epicardium, that have been established for mouse, rat and chick embryos. As these in vitro systems allow tracing the fate of these cells under natural conditions (as far as they are known) but also under artificial conditions, they provide additional insight into the developmental potential of these cells.
Primary proepicardial explants recapitulated all the cell fates observed in (pro-)epicardial cells in vivo. Smooth muscle cells, fibroblasts as well as endothelial cells and cardiomyocytes can be differentiated from proepicardial cells of mouse and chick embryos under certain conditions [19,39,40]. However, it is technically extremely challenging to isolate pure proepicardial cells devoid of contaminating surrounding tissues such as the sinus venosus myocardium. Furthermore, Katz and colleagues have shown in their studies of proepicardium-derived endothelial cells that Scx-positive proepicardial cells have a higher potential to differentiate into endothelial cells than Scx-negative proepicardial cells [37] supporting the notion that the “proepicardium” is a heterogenous cell population to start with.

{kind=link}
| Method | Reference | SE | cSMC | pCF | iCF | AVV | Endo | cEC | CM |
|---|---|---|---|---|---|---|---|---|---|
| Chick | |||||||||
| Dye labeling | [17] | X | |||||||
| [19] | X | X | |||||||
| Retroviral labeling | [16] | X | X | X (PE) | |||||
| [17] | X | X | X | X | X | ||||
| [19] | X | X | |||||||
| Quail-chick chimeras | [12] | - | |||||||
| [15] | X | X | X | X | X | ||||
| [18] | X | X | X | X | X | X | - | ||
| [19] | X | X | X | X | X | X | |||
| PE explant | [41] | X | |||||||
| [19] | X | ||||||||
| [40] | X | X | |||||||
| [42] | X | ||||||||
| [20] | X | ||||||||
| Epicardial explant | [43] | X | |||||||
| Mouse | |||||||||
| Dye labeling | [26] | X | |||||||
| [44] | X | X | |||||||
| Lineage-tracing | |||||||||
| MslnCln | [35] | X | X | - | - | ||||
| Sema3dtm1.1(GFP/cre)Cjt | [37] | X | X | X | X | X | X | ||
| Tbx18tm2(cre)Sev | [26] | X | X | - | X | ||||
| Tbx18tm4(cre)Akis | [28] | - | |||||||
| [29] | X | X | X | - | |||||
| Tcf21MerCreMer | [34] | X | X | X | X | - | - | ||
| Tg(Scx-GFP/cre)1Stzr | [36] | X | X | X | X | ||||
| Wt1tm1(EGFP/cre)Wtp | [27] | X | X | X | |||||
| [32] | X | ||||||||
| Wt1tm2(cre/ERT2)Wtp | [26] | X | X | X | |||||
| [30] | X | X* | - | ||||||
| [31] | X* | ||||||||
| PE explant | [45] | X | |||||||
| [36] | X | ||||||||
| [39] | X | X | X | X | X | ||||
| Epicardial explant | [46] | X | |||||||
| [47] | X | ||||||||
| [39] | X | X | X | - | - | ||||
| Zebrafish | |||||||||
| Tg(tcf21:DsRed2)pd3 | [38] | X | X | X | - | - | |||
Abbreviations: AVV: atrio-ventricular valve; CM: cardiomyocyte; cEC: coronary endothelial cell; cSMC: coronary smooth muscle cell (pSC: perivascular support cell in zebrafish); Endo: endocardium; iCF: interstitial cardiac fibroblast; pCF: perivascular cardiac fibroblast; PE: proepicardium; SE: subepicardium; * endogenous expression in case of Wt1.
Primary epicardial explants are derived from epithelial outgrowths of embryonic ventricles. These cells differentiate into fibroblasts and smooth muscle cells but not into cardiomyocytes and endothelial cells in culture arguing that epicardial cells have lost this potential compared to their proepicardial precursors, or that endothelial cells have been removed by this method [39].
Nonetheless, the combination of proepicardial and epicardial fate mapping efforts in vitro and in vivo points towards the character of the epicardium as a bipotential source of fibroblasts and smooth muscle cells. The proepicardium may have an additional endothelial fate but it is more likely that the proepicardium is a heterogeneous cell population to start with.
6. Mobilization and Differentiation of Epicardial Cells in The Injured Heart—Studies in Zebrafish and Mice
While it was long thought that the epicardium of the adult heart is terminally differentiated, it has now emerged that it can regain its embryonic character as a source of cells and signals for the underlying myocardium under conditions of injury. The first insight into this unexpected behavior was obtained from work in the zebrafish, a vertebrate that is able to fully regenerate its heart after loss of up to 20% of its cell mass [48]. Ventricular amputation or cryoinjury (simulating ischemic conditions after myocardial infarction in mammals) was found to activate an embryonic gene program in the epicardium of adult zebrafish [49,50]. Transcription factors like Tbx18, Wt1 and Aldh1a2 that are expressed in the embryonic but not in the adult epicardium are (re-)activated in the whole epicardium and become restricted to the epicardium covering the injury site at later stages [49,51]. Injury-activated epicardial cells start to proliferate, secrete angiogenic factors, and undergo a mesenchymal transition [52,53]. Lineage-tracing after heart apex amputation using an inducible tcf21cre fish line (Tg(tcf21:CreER)pd42) has revealed that epicardial cells contribute to vascular supporting cells but not to cardiomyocytes in the regenerating zebrafish heart [38]. A second study used the wt1b:EGFP line (Tg(wt1b:EGFP)li1) to follow Wt1-expressing cells after cryoinjury. It was observed that these Wt1-positive (epicardium-derived) cells mostly contributed to interstitial cells, “myofibroblasts” and vascular supporting cells but not to endothelial cells or cardiomyocytes in the injured heart [51]. To avoid the limitations of lineage labeling, González-Rosa and colleagues studied the fate of genetically labeled cryoinjured ventricular grafts in a transplantation model. Epicardial cells mostly contributed to the fibroblast lineage but not to cardiomyocytes after a long-term follow up (60 days post-injury) [51] further strengthening the assumption that injury-activated epicardial cells as their embryonic relatives are bipotent with respect to differentiation into fibroblasts and smooth muscle cells.
Unlike zebrafish, mammals cannot replace lost cardiomyocytes but form extensive scars after cardiac injury leading to severely impaired cardiac function. Surprisingly however, epicardial reactivation and epithelial-mesenchymal transition is a feature that also occurs in mice after myocardial infarction [53]. The lineage commitment of these reactivated epicardial cells has remained controversial.
Lineage tracing studies using Wt1tm2(cre/ERT2)Wtp2 [53] and Tg(GATA5-cre)1Krc [54] lines identified interstitial fibroblasts, “myofibroblasts” and smooth muscle cells but not cardiomyocytes and endothelial cells amongst the scar invading epicardium-derived cells after myocardial infarction. In contrast, epicardial fate mapping using a BAC derived Wt1tm1(EGFP/cre)Wtp driver found contribution of epicardial cells to the fibroblast population, “myofibroblasts” and coronary endothelium and later also to the cardiomyocyte population in an infarct model [55]. To complicate the situation even further, it was suggested from studies with the inducible Wt1tm2(cre/ERT2)Wtp2 and the constitutively Wt1tm1(EGFP/cre)Wtp driver lines that treating mice with thymosin beta4 before induction of myocardial ischema accelerates and enhances the mobilization of epicardium-derived cells and induces their contribution to the cardiomyocyte lineage [56]. In a subsequent study, Zhou and co-workers used the same Wt1tm2(cre/ERT2)Wtp2 line for lineage tracing and confirmed the beneficial effects of thymosin beta4 on revascularization and scar reduction, but did not find any evidence that thymosin beta4 treatment after myocardial infarction reprograms epicardial cells into cardiomyocytes [57]. As already mentioned for lineage-tracing studies during embryonic development, Wt1 is expressed in coronary endothelial cells (and possibly in cardiomyocytes) and might be reactivated by the ischemic conditions after myocardial infarction [30,31,57,58] putting a question mark behind the usefulness of Wt1-based lineage tracing systems for epicardial fate mapping under injury conditions just as in the embryonic situation.
Nonetheless, we can safely state that the epicardium is reactivated after cardiac injury to participate in the wound healing response by providing (mostly) fibroblasts and smooth muscle cells to the injury site—a process highly similar in zebrafish and mice (for a summary see Table 2). A contribution to cardiomyocytes and endothelial cells cannot be excluded at this point in the mouse but is likely to be a minor one. More intriguingly, epicardial cells provide proangiogenic factors to the heart [51,53] that may improve cardiac function after injury. While the epicardial injury program is likely to be conserved in lower and higher vertebrates, the myocardial response seems fundamentally different as mammalian cardiomyocytes unlike the zebrafish do not dedifferentiate and re-enter the cell cycle. It remains to be seen whether this reflects an intrinsic difference within the cardiomyocyte or reflects the difference in a growth promoting environment in which the generation of the epicardium may also play a role. It may be noted that that the neonatal murine heart still has the ability to regenerate after ventricular resection [59].
| Injury | Method | iC | CF | pSC | CM | EC |
|---|---|---|---|---|---|---|
| Zebrafish | ||||||
| Cryoinjury | Transplantation [51] | X | X | X | - | - |
| (infarct model) | Lineage-tracing | X | X | X | - | - |
| Ventricular resection | Lineage-tracing | X | - | - | ||
| Mouse | ||||||
| Myocardial infarction | Lineage-tracing | X | X | X | X | X /- |
| Lineage-tracing | X | X | X | - | - | |
| Lineage-tracing | X | X | - | |||
Abbreviations: CM: cardiomyocyte; EC: endothelial cell; iC: interstitial cell; CF: cardiac fibroblast; pSC: perivascular support cell (zebrafish)/coronary smooth muscle cells (mammals).
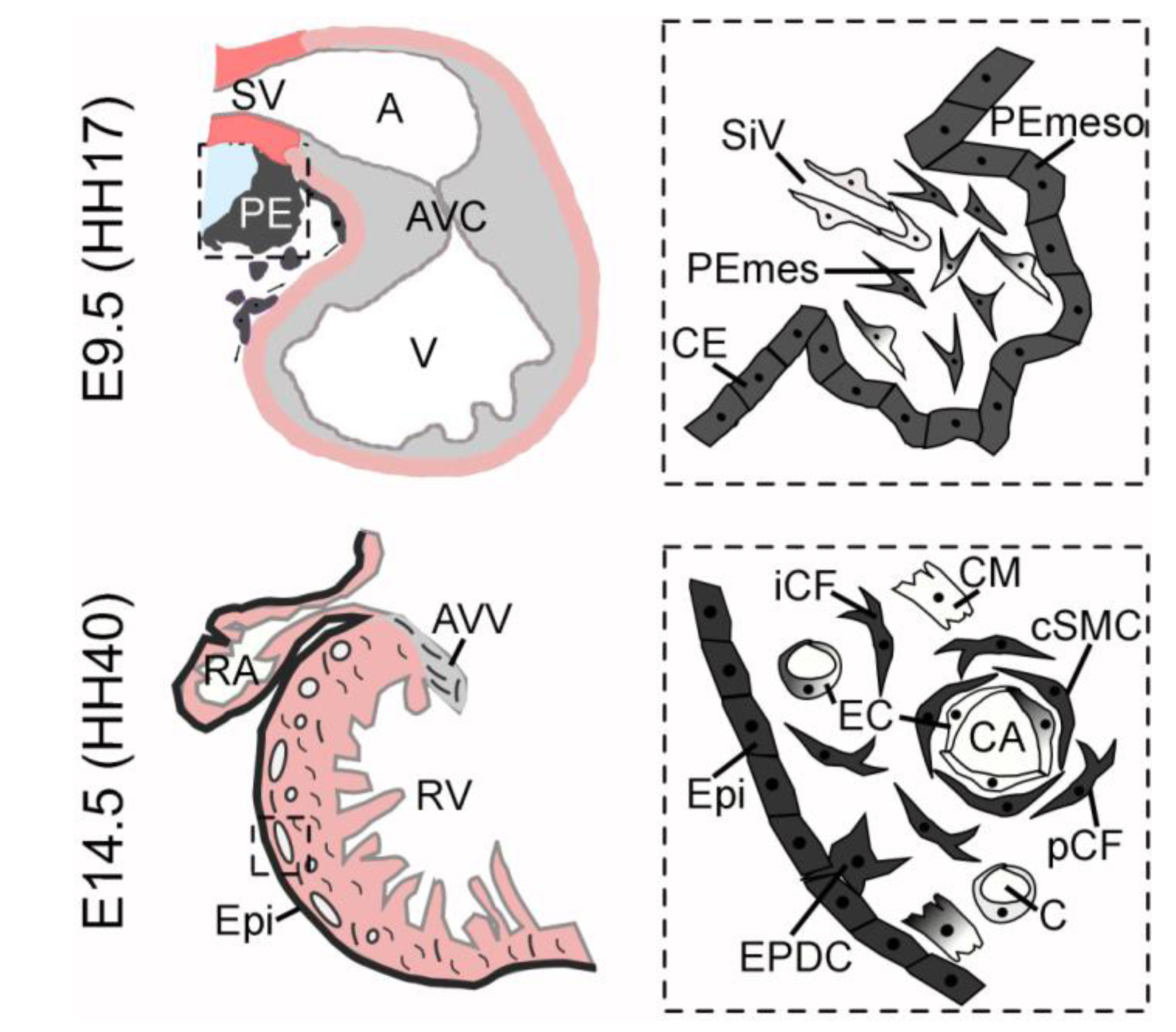
Figure 1.
Scheme of (pro-)epicardial lineages. The upper row shows a schematic view of an E9.5 sagittally sectioned murine heart (similar to HH17 avian heart) on the left side. The proepicardium (PE) is indicated in dark grey, the chamber myocardium in light pink, the sinus venosus mesenchyme (SV) in red and the cardiac jelly and cushion mesenchyme in light grey. Note the close proximity of proepicardium, sinus venosus mesenchyme, septum transversum and liver, respectively (light blue). The right scheme represents a magnification of the proepicardium (boxed area). Coelomic epithelium (CE) and the proepicardial mesothelium (PEmeso) are indicated in dark grey and share the expression of Tbx18 and Wt1. Proepicardial mesenchymal cells (PEmes) are highly diverse. Wt1-, Tbx18, Scx-, Sema3D- and Tcf21-expressing cells are found within the mesenchyme with varying overlapping expression patterns. Their individual descent and contribution to the heart have remained largely elusive. Sinusoid-like vessels (SiV) reach into the proepicardium and may provide endothelial cells. The lower row shows a scheme of the murine heart at E14.5 (resembling an avian heart at HH40). Epicardium-derived cells (EPDCs) are indicated in dark grey and invade the ventricular myocardium as well as the cushion tissue of the atrio-ventricular valves (AVV). A higher magnification of the boxed area within the right ventricle (RV) is given to indicate the fate of these cells. They mainly contribute to coronary smooth muscle cells (cSMC) and interstitial (iCF) as well as perivascular fibroblasts (pCF). An epicardial origin of endothelial cells (EC) and cardiomyocytes (CM) is unlikely (indicated by a gray-white gradient within these cells).
Figure 1.
Scheme of (pro-)epicardial lineages. The upper row shows a schematic view of an E9.5 sagittally sectioned murine heart (similar to HH17 avian heart) on the left side. The proepicardium (PE) is indicated in dark grey, the chamber myocardium in light pink, the sinus venosus mesenchyme (SV) in red and the cardiac jelly and cushion mesenchyme in light grey. Note the close proximity of proepicardium, sinus venosus mesenchyme, septum transversum and liver, respectively (light blue). The right scheme represents a magnification of the proepicardium (boxed area). Coelomic epithelium (CE) and the proepicardial mesothelium (PEmeso) are indicated in dark grey and share the expression of Tbx18 and Wt1. Proepicardial mesenchymal cells (PEmes) are highly diverse. Wt1-, Tbx18, Scx-, Sema3D- and Tcf21-expressing cells are found within the mesenchyme with varying overlapping expression patterns. Their individual descent and contribution to the heart have remained largely elusive. Sinusoid-like vessels (SiV) reach into the proepicardium and may provide endothelial cells. The lower row shows a scheme of the murine heart at E14.5 (resembling an avian heart at HH40). Epicardium-derived cells (EPDCs) are indicated in dark grey and invade the ventricular myocardium as well as the cushion tissue of the atrio-ventricular valves (AVV). A higher magnification of the boxed area within the right ventricle (RV) is given to indicate the fate of these cells. They mainly contribute to coronary smooth muscle cells (cSMC) and interstitial (iCF) as well as perivascular fibroblasts (pCF). An epicardial origin of endothelial cells (EC) and cardiomyocytes (CM) is unlikely (indicated by a gray-white gradient within these cells).

7. Summary and Perspectives
The last 20 years have seen a large number of studies that collectively suggest that the epicardium is a crucial source of cells for the developing and regenerating vertebrate heart. Smooth muscle cells and fibroblasts are the two main cell types that differentiate from epicardium-derived mesenchymal cells, making the epicardium an essential tissue for the generation of the cardiac fibroskeleton and the coronary vasculature (Figure 1). It remains an open question as to how individual epicardial cells are mobilized and when they become committed to one or another fate. Although a number of reports have suggested an endothelial and myocardial differentiation of epicardial cells, this contribution—if not just a mere technical artefact—is definitely a very minor one both in development and regeneration. Nonetheless, efforts to find suitable cocktails of factors that can reprogram epicardial cells to one of these fates may be worthwhile to pursue in the future.
While the epicardium derives from the proepicardium, the latter may present a more complex pool of cells that comprises epicardial precursors as well as endothelial and myocardial cells. However, we do need a more elaborate panel of markers that distinguish the proepicardium from surrounding tissue and/or describe its lineage segregation to better state its role as a cell source in development.
Acknowledgments
This work was supported by a grant from the Hannover Biomedical Research School (HBRS) to F.G. and by grants from the German Research Foundation (DFG) for the Cluster of Excellence REBIRTH (From Regenerative Biology to Reconstructive Therapy) and for the Clinical Research Group KFO136 at Hannover Medical School to A.K.
References and Notes
- Vincent, S.D.; Buckingham, M.E. How to make a heart: the origin and regulation of cardiac progenitor cells. Curr. Top. Dev. Biol. 2010, 90, 1–41. [Google Scholar] [CrossRef]
- Saga, Y.; Kitajima, S.; Miyagawa-Tomita, S. Mesp1 expression is the earliest sign of cardiovascular development. Trends Cardiovasc. Med. 2000, 10, 345–352. [Google Scholar] [CrossRef]
- Serluca, F.C. Development of the proepicardial organ in the zebrafish. Dev. Biol. 2008, 315, 18–27. [Google Scholar] [CrossRef]
- Rodgers, L.S.; Lalani, S.; Runyan, R.B.; Camenisch, T.D. Differential growth and multicellular villi direct proepicardial translocation to the developing mouse heart. Dev. Dyn. 2008, 237, 145–152. [Google Scholar] [CrossRef]
- Komiyama, M.; Ito, K.; Shimada, Y. Origin and development of the epicardium in the mouse embryo. Anat. Embryol. (Berl) 1987, 176, 183–189. [Google Scholar] [CrossRef]
- Schulte, I.; Schlueter, J.; Abu-Issa, R.; Brand, T.; Manner, J. Morphological and molecular left-right asymmetries in the development of the proepicardium: a comparative analysis on mouse and chick embryos. Dev. Dyn. 2007, 236, 684–695. [Google Scholar] [CrossRef]
- Perez-Pomares, J.M.; Macias, D.; Garcia-Garrido, L.; Munoz-Chapuli, R. The origin of the subepicardial mesenchyme in the avian embryo: an immunohistochemical and quail-chick chimera study. Dev. Biol. 1998, 200, 57–68. [Google Scholar]
- Cossette, S.; Misra, R. The identification of different endothelial cell populations within the mouse proepicardium. Dev. Dyn. 2011, 240, 2344–2353. [Google Scholar]
- Viragh, S.; Gittenberger-de Groot, A.C.; Poelmann, R.E.; Kalman, F. Early development of quail heart epicardium and associated vascular and glandular structures. Anat. Embryol. (Berl) 1993, 188, 381–393. [Google Scholar]
- Wilting, J.; Buttler, K.; Schulte, I.; Papoutsi, M.; Schweigerer, L.; Manner, J. The proepicardium delivers hemangioblasts but not lymphangioblasts to the developing heart. Dev. Biol. 2007, 305, 451–459. [Google Scholar]
- Manner, J. The development of pericardial villi in the chick embryo. Anat. Embryol. (Berl) 1992, 186, 379–385. [Google Scholar] [CrossRef]
- Poelmann, R.E.; Gittenberger-de Groot, A.C.; Mentink, M.M.; Bokenkamp, R.; Hogers, B. Development of the cardiac coronary vascular endothelium, studied with antiendothelial antibodies, in chicken-quail chimeras. Circ. Res. 1993, 73, 559–568. [Google Scholar] [CrossRef]
- Olivey, H.E.; Svensson, E.C. Epicardial-myocardial signaling directing coronary vasculogenesis. Circ. Res. 2010, 106, 818–832. [Google Scholar] [CrossRef]
- Perez-Pomares, J.M.; Macias, D.; Garcia-Garrido, L.; Munoz-Chapuli, R. Contribution of the primitive epicardium to the subepicardial mesenchyme in hamster and chick embryos. Dev. Dyn. 1997, 210, 96–105. [Google Scholar] [CrossRef]
- Gittenberger-de Groot, A.C.; Vrancken Peeters, M.P.; Mentink, M.M.; Gourdie, R.G.; Poelmann, R.E. Epicardium-derived cells contribute a novel population to the myocardial wall and the atrioventricular cushions. Circ. Res. 1998, 82, 1043–1052. [Google Scholar] [CrossRef]
- Mikawa, T.; Gourdie, R.G. Pericardial mesoderm generates a population of coronary smooth muscle cells migrating into the heart along with ingrowth of the epicardial organ. Dev. Biol. 1996, 174, 221–232. [Google Scholar] [CrossRef]
- Dettman, R.W.; Denetclaw, W., Jr.; Ordahl, C.P.; Bristow, J. Common epicardial origin of coronary vascular smooth muscle, perivascular fibroblasts, and intermyocardial fibroblasts in the avian heart. Dev. Biol. 1998, 193, 169–181. [Google Scholar] [CrossRef]
- Manner, J. Does the subepicardial mesenchyme contribute myocardioblasts to the myocardium of the chick embryo heart? A quail-chick chimera study tracing the fate of the epicardial primordium. Anat. Rec. 1999, 255, 212–226. [Google Scholar]
- Perez-Pomares, J.M.; Carmona, R.; Gonzalez-Iriarte, M.; Atencia, G.; Wessels, A.; Munoz-Chapuli, R. Origin of coronary endothelial cells from epicardial mesothelium in avian embryos. Int. J. Dev. Biol. 2002, 46, 1005–1013. [Google Scholar]
- Guadix, J.A.; Carmona, R.; Munoz-Chapuli, R.; Perez-Pomares, J.M. In vivo and in vitro analysis of the vasculogenic potential of avian proepicardial and epicardial cells. Dev Dyn. 2006, 235, 1014–1026. [Google Scholar] [CrossRef]
- Pluck, A. Conditional mutagenesis in mice: The Cre/loxP recombination system. Int. J. Exp. Pathol. 1996, 77, 269–278. [Google Scholar]
- Kretzschmar, K.; Watt, F.M. Lineage tracing. Cell 2012, 148, 33–45. [Google Scholar] [CrossRef]
- Wilm, B.; Ipenberg, A.; Hastie, N.D.; Burch, J.B.; Bader, D.M. The serosal mesothelium is a major source of smooth muscle cells of the gut vasculature. Development 2005, 132, 5317–5328. [Google Scholar] [CrossRef]
- Moore, A.W.; McInnes, L.; Kreidberg, J.; Hastie, N.D.; Schedl, A. YAC complementation shows a requirement for Wt1 in the development of epicardium, adrenal gland and throughout nephrogenesis. Development 1999, 126, 1845–1857. [Google Scholar]
- Merki, E.; Zamora, M.; Raya, A.; Kawakami, Y.; Wang, J.; Zhang, X.; Burch, J.; Kubalak, S.W.; Kaliman, P.; Izpisua Belmonte, J.C.; Chien, K.R.; Ruiz-Lozano, P. Epicardial retinoid X receptor alpha is required for myocardial growth and coronary artery formation. Proc. Natl. Acad. Sci. USA 2005, 102, 18455–18460. [Google Scholar] [CrossRef]
- Cai, C.L.; Martin, J.C.; Sun, Y.; Cui, L.; Wang, L.; Ouyang, K.; Yang, L.; Bu, L.; Liang, X.; Zhang, X.; Stallcup, W.B.; Denton, C.P.; McCulloch, A.; Chen, J.; Evans, S.M. A myocardial lineage derives from Tbx18 epicardial cells. Nature 2008, 454, 104–108. [Google Scholar] [CrossRef]
- Zhou, B.; Ma, Q.; Rajagopal, S.; Wu, S.M.; Domian, I.; Rivera-Feliciano, J.; Jiang, D.; von Gise, A.; Ikeda, S.; Chien, K.R.; Pu, W.T. Epicardial progenitors contribute to the cardiomyocyte lineage in the developing heart. Nature 2008, 454, 109–113. [Google Scholar] [CrossRef]
- Christoffels, V.M.; Grieskamp, T.; Norden, J.; Mommersteeg, M.T.; Rudat, C.; Kispert, A. Tbx18 and the fate of epicardial progenitors. Nature 2009, 458, E8–E9; Discussion E9–E10. [Google Scholar] [CrossRef]
- Grieskamp, T.; Rudat, C.; Ludtke, T.H.; Norden, J.; Kispert, A. Notch signaling regulates smooth muscle differentiation of epicardium-derived cells. Circ. Res. 2011, 108, 813–823. [Google Scholar] [CrossRef]
- Rudat, C.; Kispert, A. Wt1 and epicardial fate mapping. Circ. Res. 2012, 111, 165–169. [Google Scholar] [CrossRef]
- Zhou, B.; Pu, W.T. Genetic Cre-loxP assessment of epicardial cell fate using Wt1-driven Cre alleles. Circ. Res. 2012, 111, e276–e280. [Google Scholar] [CrossRef]
- Wessels, A.; van den Hoff, M.J.; Adamo, R.F.; Phelps, A.L.; Lockhart, M.M.; Sauls, K.; Briggs, L.E.; Norris, R.A.; van Wijk, B.; Perez-Pomares, J.M.; Dettman, R.W.; Burch, J.B. Epicardially derived fibroblasts preferentially contribute to the parietal leaflets of the atrioventricular valves in the murine heart. Dev. Biol. 2012, 366, 111–124. [Google Scholar] [CrossRef]
- Zhou, B.; von Gise, A.; Ma, Q.; Hu, Y.W.; Pu, W.T. Genetic fate mapping demonstrates contribution of epicardium-derived cells to the annulus fibrosis of the mammalian heart. Dev. Biol. 2010, 338, 251–261. [Google Scholar] [CrossRef]
- Acharya, A.; Baek, S.T.; Huang, G.; Eskiocak, B.; Goetsch, S.; Sung, C.Y.; Banfi, S.; Sauer, M.F.; Olsen, G.S.; Duffield, J.S.; Olson, E.N.; Tallquist, M.D. The bHLH transcription factor Tcf21 is required for lineage-specific EMT of cardiac fibroblast progenitors. Development 2012, 139, 2139–2149. [Google Scholar] [CrossRef]
- Rinkevich, Y.; Mori, T.; Sahoo, D.; Xu, P.X.; Bermingham, J.R., Jr.; Weissman, I.L. Identification and prospective isolation of a mesothelial precursor lineage giving rise to smooth muscle cells and fibroblasts for mammalian internal organs, and their vasculature. Nat. Cell. Biol. 2012, 14, 1251–1260. [Google Scholar] [CrossRef]
- Red-Horse, K.; Ueno, H.; Weissman, I.L.; Krasnow, M.A. Coronary arteries form by developmental reprogramming of venous cells. Nature 2010, 464, 549–553. [Google Scholar] [CrossRef]
- Katz, T.C.; Singh, M.K.; Degenhardt, K.; Rivera-Feliciano, J.; Johnson, R.L.; Epstein, J.A.; Tabin, C.J. Distinct compartments of the proepicardial organ give rise to coronary vascular endothelial cells. Dev. Cell. 2012, 22, 639–650. [Google Scholar] [CrossRef]
- Kikuchi, K.; Gupta, V.; Wang, J.; Holdway, J.E.; Wills, A.A.; Fang, Y.; Poss, K.D. tcf21+ epicardial cells adopt non-myocardial fates during zebrafish heart development and regeneration. Development 2011, 138, 2895–2902. [Google Scholar] [CrossRef]
- Ruiz-Villalba, A.; Ziogas, A.; Ehrbar, M.; Perez-Pomares, J.M. Characterization of epicardial-derived cardiac interstitial cells: Differentiation and mobilization of heart fibroblast progenitors. PLoS One 2013, 8, e53694. [Google Scholar]
- Kruithof, B.P.; van Wijk, B.; Somi, S.; Kruithof-de Julio, M.; Perez Pomares, J.M.; Weesie, F.; Wessels, A.; Moorman, A.F.; van den Hoff, M.J. BMP and FGF regulate the differentiation of multipotential pericardial mesoderm into the myocardial or epicardial lineage. Dev. Biol. 2006, 295, 507–522. [Google Scholar] [CrossRef]
- Lu, J.; Landerholm, T.E.; Wei, J.S.; Dong, X.R.; Wu, S.P.; Liu, X.; Nagata, K.; Inagaki, M.; Majesky, M.W. Coronary smooth muscle differentiation from proepicardial cells requires rhoA-mediated actin reorganization and p160 rho-kinase activity. Dev. Biol. 2001, 240, 404–418. [Google Scholar] [CrossRef]
- Schlueter, J.; Manner, J.; Brand, T. BMP is an important regulator of proepicardial identity in the chick embryo. Dev. Biol. 2006, 295, 546–558. [Google Scholar]
- Compton, L.A.; Potash, D.A.; Mundell, N.A.; Barnett, J.V. Transforming growth factor-beta induces loss of epithelial character and smooth muscle cell differentiation in epicardial cells. Dev. Dyn. 2006, 235, 82–93. [Google Scholar]
- Vega-Hernandez, M.; Kovacs, A.; De Langhe, S.; Ornitz, D.M. FGF10/FGFR2b signaling is essential for cardiac fibroblast development and growth of the myocardium. Development 2011, 138, 3331–3340. [Google Scholar]
- del Monte, G.; Casanova, J.C.; Guadix, J.A.; MacGrogan, D.; Burch, J.B.; Perez-Pomares, J.M.; de la Pompa, J.L. Differential Notch signaling in the epicardium is required for cardiac inflow development and coronary vessel morphogenesis. Circ. Res. 2011, 108, 824–836. [Google Scholar]
- Austin, A.F.; Compton, L.A.; Love, J.D.; Brown, C.B.; Barnett, J.V. Primary and immortalized mouse epicardial cells undergo differentiation in response to TGFbeta. Dev. Dyn. 2008, 237, 366–376. [Google Scholar] [CrossRef]
- Zamora, M.; Manner, J.; Ruiz-Lozano, P. Epicardium-derived progenitor cells require beta-catenin for coronary artery formation. Proc. Natl. Acad. Sci. USA 2007, 104, 18109–18114. [Google Scholar] [CrossRef]
- Poss, K.D.; Wilson, L.G.; Keating, M.T. Heart regeneration in zebrafish. Science 2002, 298, 2188–2190. [Google Scholar]
- Lepilina, A.; Coon, A.N.; Kikuchi, K.; Holdway, J.E.; Roberts, R.W.; Burns, C.G.; Poss, K.D. A dynamic epicardial injury response supports progenitor cell activity during zebrafish heart regeneration. Cell 2006, 127, 607–619. [Google Scholar] [CrossRef]
- Schnabel, K.; Wu, C.C.; Kurth, T.; Weidinger, G. Regeneration of cryoinjury induced necrotic heart lesions in zebrafish is associated with epicardial activation and cardiomyocyte proliferation. PLoS One 2011, 6, e18503. [Google Scholar]
- Gonzalez-Rosa, J.M.; Peralta, M.; Mercader, N. Pan-epicardial lineage tracing reveals that epicardium derived cells give rise to myofibroblasts and perivascular cells during zebrafish heart regeneration. Dev. Biol. 2012, 370, 173–186. [Google Scholar] [CrossRef]
- Kim, J.; Wu, Q.; Zhang, Y.; Wiens, K.M.; Huang, Y.; Rubin, N.; Shimada, H.; Handin, R.I.; Chao, M.Y.; Tuan, T.L.; Starnes, V.A.; Lien, C.L. PDGF signaling is required for epicardial function and blood vessel formation in regenerating zebrafish hearts. Proc. Natl. Acad. Sci. USA 2010, 107, 17206–17210. [Google Scholar] [CrossRef]
- Zhou, B.; Honor, L.B.; He, H.; Ma, Q.; Oh, J.H.; Butterfield, C.; Lin, R.Z.; Melero-Martin, J.M.; Dolmatova, E.; Duffy, H.S.; Gise, A.; Zhou, P.; Hu, Y.W.; Wang, G.; Zhang, B.; Wang, L.; Hall, J.L.; Moses, M.A.; McGowan, F.X.; Pu, W.T. Adult mouse epicardium modulates myocardial injury by secreting paracrine factors. J. Clin. Invest. 2011, 121, 1894–1904. [Google Scholar] [CrossRef]
- Smart, N.; Risebro, C.A.; Clark, J.E.; Ehler, E.; Miquerol, L.; Rossdeutsch, A.; Marber, M.S.; Riley, P.R. Thymosin beta4 facilitates epicardial neovascularization of the injured adult heart. Ann. N. Y. Acad. Sci. 2010, 1194, 97–104. [Google Scholar]
- van Wijk, B.; Gunst, Q.D.; Moorman, A.F.; van den Hoff, M.J. Cardiac regeneration from activated epicardium. PLoS One 2012, 7, e44692. [Google Scholar]
- Smart, N.; Bollini, S.; Dube, K.N.; Vieira, J.M.; Zhou, B.; Davidson, S.; Yellon, D.; Riegler, J.; Price, A.N.; Lythgoe, M.F.; Pu, W.T.; Riley, P.R. De novo cardiomyocytes from within the activated adult heart after injury. Nature 2011, 474, 640–644. [Google Scholar] [CrossRef]
- Zhou, B.; Honor, L.B.; Ma, Q.; Oh, J.H.; Lin, R.Z.; Melero-Martin, J.M.; von Gise, A.; Zhou, P.; Hu, T.; He, L.; Wu, K.H.; Zhang, H.; Zhang, Y.; Pu, W.T. Thymosin beta 4 treatment after myocardial infarction does not reprogram epicardial cells into cardiomyocytes. J. Mol. Cell. Cardiol. 2011, 52, 43–47. [Google Scholar]
- Wagner, K.D.; Wagner, N.; Bondke, A.; Nafz, B.; Flemming, B.; Theres, H.; Scholz, H. The Wilms' tumor suppressor Wt1 is expressed in the coronary vasculature after myocardial infarction. FASEB J. 2002, 16, 1117–1119. [Google Scholar]
- Porrello, E.R.; Mahmoud, A.I.; Simpson, E.; Hill, J.A.; Richardson, J.A.; Olson, E.N.; Sadek, H.A. Transient regenerative potential of the neonatal mouse heart. Science 2011, 331, 1078–1080. [Google Scholar] [CrossRef]
- Jopling, C.; Sleep, E.; Raya, M.; Marti, M.; Raya, A.; Izpisua Belmonte, J.C. Zebrafish heart regeneration occurs by cardiomyocyte dedifferentiation and proliferation. Nature 2010, 464, 606–609. [Google Scholar]
- Smart, N.; Bollini, S.; Dube, K.N.; Vieira, J.M.; Zhou, B.; Riegler, J.; Price, A.N.; Lythgoe, M.F.; Davidson, S.; Yellon, D.; Pu, W.T.; Riley, P.R. Myocardial regeneration: expanding the repertoire of thymosin beta4 in the ischemic heart. Ann. N. Y. Acad. Sci. 2012, 1269, 92–101. [Google Scholar]
© 2013 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
MDPI and ACS Style
Greulich, F.; Kispert, A. Epicardial Lineages. J. Dev. Biol. 2013, 1, 32-46. https://doi.org/10.3390/jdb1010032
AMA Style
Greulich F, Kispert A. Epicardial Lineages. Journal of Developmental Biology. 2013; 1(1):32-46. https://doi.org/10.3390/jdb1010032
Chicago/Turabian StyleGreulich, Franziska, and Andreas Kispert. 2013. "Epicardial Lineages" Journal of Developmental Biology 1, no. 1: 32-46. https://doi.org/10.3390/jdb1010032