
The Combination of Both Heat and Water Stresses May Worsen Botryosphaeria Dieback Symptoms in Grapevine

 , ,
, ,  ,
,  ,
, 
Abstract
:1. Introduction
2. Results and Discussion
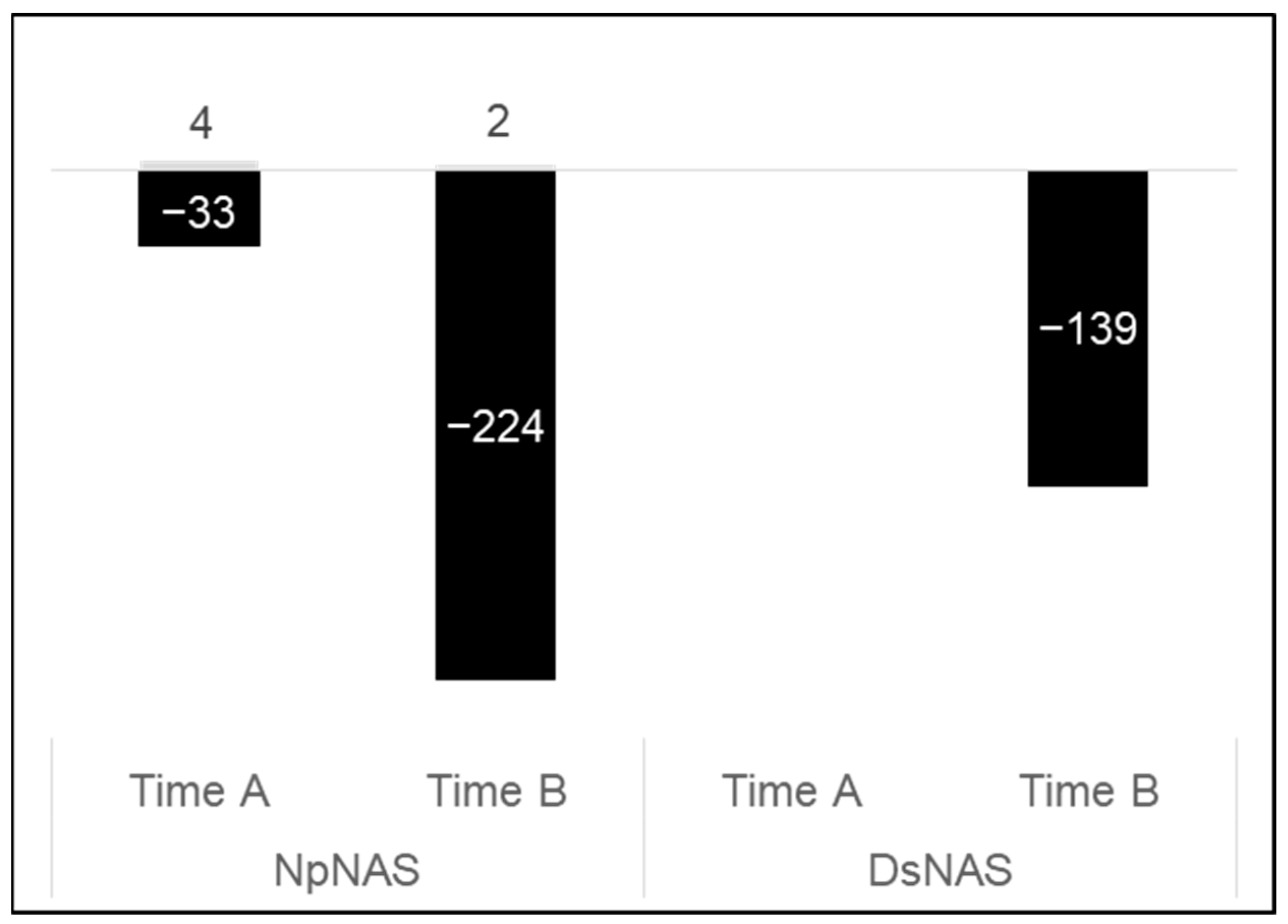
2.1. Measurement of Ds and Np Necrosis under Different Stress Combinations
2.2. Transcriptomic Analysis of Single Stress Responses
2.2.1. Heat Stress Impact on Leaf Transcriptome
2.2.2. Water Stress Impact on Leaf Transcriptome
2.2.3. Fungal Infection Impact on Leaf Transcriptome
2.3. Transcriptomic Analysis of Combined Stress Responses
2.4. Metabolomic Analysis of Single Stress Responses
2.4.1. Water Stress Impact on Leaf Metabolome
2.4.2. Heat Stress Impact on Leaf Metabolome
2.4.3. Fungal Infection Impact on Leaf Metabolome
2.5. Metabolomic Analysis of Combined Stress Responses
2.6. Molecular and Metabolic Response when Combining Infection an Abiotic Stresses
2.6.1. Analysis of Leaf Transcriptome following Combined Biotic and Abiotic Interaction
2.6.2. Analysis of Leaf Metabolome following Biotic and Abiotic Interactions
2.7. Conclusions
3. Materials and Methods
3.1. Experimental Design
3.1.1. Plant Material and Growth Conditions
3.1.2. Artificial Fungal Inoculation
3.1.3. Abiotic Stress Treatments
3.2. Sample Collection
3.3. Phytopathogen Re-Isolation and Necrosis Measurement
3.4. Transcriptomic Analysis
3.4.1. RNA Extraction, cDNA Library Construction, and Illumina RNA Sequencing
3.4.2. Read Mapping, Assembly, and Differential-Expression Analysis
3.4.3. Gene Ontology Analysis
3.4.4. Illumina RNA-Seq Results Validation by qRT-PCR
3.5. Metabolomic Analysis
3.5.1. FT-ICR-MS Analysis
3.5.2. Data Analysis
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Guerin-Dubrana, L.; Fontaine, F.; Mugnai, L. Grapevine Trunk Disease in European and Mediterranean Vineyards: Occurrence, Distribution and Associated Disease-Affecting Cultural Factors. Phytopathol. Mediterr. 2019, 58, 49–71. [Google Scholar] [CrossRef]
- Fontaine, F.; Gramaje, D.; Armengol, J.; Smart, R.; Nagy, Z.A.; Borgo, M.; Rego, C.; Corio-Costet, M.F. Grapevine Trunk Diseases. A Review; OIV publications: Paris, France, 2016; pp. 1–24. [Google Scholar]
- Dubos, B.; Cere, L.; Larignon, P.; Fulchic, R. Observation on Black Dead Arm in French Vineyards. Phytopathol. Mediterr. 2001, 40, 336–342. [Google Scholar] [CrossRef]
- Bertsch, C.; Ramírez-Suero, M.; Magnin-Robert, M.; Larignon, P.; Chong, J.; Abou-Mansour, E.; Spagnolo, A.; Clément, C.; Fontaine, F. Grapevine Trunk Diseases: Complex and Still Poorly Understood. Plant Pathol. 2013, 62, 243–265. [Google Scholar] [CrossRef]
- Larignon, P.; Spagnolo, A.; Bertsch, C.; Fontaine, F. First Report of Young Grapevine Decline Caused by Neofusicoccum parvum in France. Plant Dis. 2015, 99, 1859. [Google Scholar] [CrossRef]
- van Niekerk, J.M.; Crous, P.W.; Groenewald, J.Z.E.; Fourie, P.H.; Halleen, F. DNA Phylogeny, Morphology and Pathogenicity of Botryosphaeria Species on Grapevines. Mycologia 2004, 96, 781–798. [Google Scholar] [CrossRef] [PubMed]
- Spagnolo, A.; Magnin-Robert, M.; Alayi, T.D.; Cilindre, C.; Schaeffer-Reiss, C.; Van Dorsselaer, A.; Clément, C.; Larignon, P.; Ramirez-Suero, M.; Chong, J.; et al. Differential Responses of Three Grapevine Cultivars to Botryosphaeria Dieback. Phytopathology 2014, 104, 1021–1035. [Google Scholar] [CrossRef]
- Úrbez-Torres, J.R. The Status of Botryosphaeriaceae Species Infecting Grapevines. Phytopathol. Mediterr. 2011, 50, 5–45. [Google Scholar] [CrossRef]
- Mugnai, L.; Graniti, A.; Surico, G. Esca (Black Measles) and Brown Wood-Streaking: Two Old and Elusive Diseases of Grapevines. Plant Dis. 1999, 83, 404–418. [Google Scholar] [CrossRef]
- Bruez, E.; Vallance, J.; Gerbore, J.; Lecomte, P.; Da Costa, J.P.; Guerin-Dubrana, L.; Rey, P. Analyses of the Temporal Dynamics of Fungal Communities Colonizing the Healthy Wood Tissues of Esca Leaf-Symptomatic and Asymptomatic Vines. PLoS ONE 2014, 9, e95928. [Google Scholar] [CrossRef]
- Bruez, E.; Baumgartner, K.; Bastien, S.; Travadon, R.; Guérin-Dubrana, L.; Rey, P. Various Fungal Communities Colonise the Functional Wood Tissues of Old Grapevines Externally Free from Grapevine Trunk Disease Symptoms. Aust. J. Grape Wine Res. 2016, 22, 288–295. [Google Scholar] [CrossRef]
- Ferreira, J.H.S.; van Wyk, P.S.; Calitz, F.J. Slow Dieback of Grapevine in South Africa: Stress Related Predisposition of Young Vines for Infection Be Phaeoacremonium chlamydosporum. S. Afr. J. Enol. Vitic. 1999, 20, 43–46. [Google Scholar]
- Sosnowski, M.R.; Shtienberg, D.; Creaser, M.L.; Wicks, T.J.; Lardner, R.; Scott, E.S. The Influence of Climate on Foliar Symptoms of Eutypa Dieback in Grapevines. Phytopathology 2007, 97, 1284–1289. [Google Scholar] [CrossRef] [PubMed]
- Fischer, M.; Peighami-Ashnaei, S. Grapevine, Esca Complex, and Environment: The Disease Triangle. Phytopathol. Mediterr. 2019, 58, 17–37. [Google Scholar] [CrossRef]
- Songy, A.; Fernandez, O.; Clément, C.; Larignon, P.; Fontaine, F. Grapevine Trunk Diseases under Thermal and Water Stresses. Planta 2019, 249, 1655–1679. [Google Scholar] [CrossRef]
- Larignon, P.; Fontaine, F.; Farine, S.; Clément, C.; Bertsch, C. Esca et Black Dead Arm: Deux Acteurs Majeurs Des Maladies Du Bois Chez La Vigne. Comptes Rendus Biol. 2009, 332, 765–783. [Google Scholar] [CrossRef]
- van Niekerk, J.; Strever, A.E.; Toit, G.P.D.; Halleen, F.; Fourie, P.H. Influence of Water Stress on Botryosphaeriaceae Disease Expression in Grapevines. Phytopathol. Mediterr. 2011, 50, 151–165. [Google Scholar] [CrossRef]
- Qiu, Y.; Steel, C.C.; Ash, G.J.; Savocchia, S. Effects of Temperature and Water Stress on the Virulence of Botryosphaeriaceae spp. Causing Dieback of Grapevines and Their Predicted Distribution Using CLIMEX in Australia. Acta Hortic. 2016, 1115, 171–182. [Google Scholar] [CrossRef]
- Paolinelli-Alfonso, M.; Villalobos-Escobedo, J.M.; Rolshausen, P.; Herrera-Estrella, A.; Galindo-Sánchez, C.; López-Hernández, J.F.; Hernandez-Martinez, R. Global Transcriptional Analysis Suggests Lasiodiplodia theobromae Pathogenicity Factors Involved in Modulation of Grapevine Defensive Response. BMC Genom. 2016, 17, 615. [Google Scholar] [CrossRef]
- Atkinson, N.J.; Urwin, P.E. The Interaction of Plant Biotic and Abiotic Stresses: From Genes to the Field. J. Exp. Bot. 2012, 63, 3523–3543. [Google Scholar] [CrossRef]
- Suzuki, N.; Rivero, R.M.; Shulaev, V.; Blumwald, E.; Mittler, R. Abiotic and Biotic Stress Combinations. New Phytol. 2014, 203, 32–43. [Google Scholar] [CrossRef]
- Hannah, L.; Roehrdanz, P.R.; Ikegami, M.; Shepard, A.V.; Shaw, M.R.; Tabor, G.; Zhi, L.; Marquet, P.A.; Hijmans, R.J. Climate Change, Wine, and Conservation. Proc. Natl. Acad. Sci. USA 2013, 110, 6907–6912. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ollat, N.; Zito, S.; Richard, Y.; Aigrain, P.; Brugière, F.; Duchêne, E.; Cortazar-Atauri, I.G.D.; Gautier, J.; Giraud-Héraud, E.; Hannin, H.; et al. La diversité des vignobles français face au changement climatique: Simulations climatiques et prospective participative. Climatologie 2021, 18, 3. [Google Scholar] [CrossRef]
- Chaves, M.M.; Zarrouk, O.; Francisco, R.; Costa, J.M.; Santos, T.; Regalado, A.P.; Rodrigues, M.L.; Lopes, C.M. Grapevine under Deficit Irrigation: Hints from Physiological and Molecular Data. Ann. Bot. 2010, 105, 661–676. [Google Scholar] [CrossRef] [PubMed]
- Edwards, E.J.; Smithson, L.; Graham, D.C.; Clingeleffer, P.R. Grapevine Canopy Response to a High-Temperature Event during Deficit Irrigation. Aust. J. Grape Wine Res. 2011, 17, 153–161. [Google Scholar] [CrossRef]
- Lovisolo, C.; Perrone, I.; Carra, A.; Ferrandino, A.; Flexas, J.; Medrano, H.; Schubert, A.; Lovisolo, C.; Perrone, I.; Carra, A.; et al. Drought-Induced Changes in Development and Function of Grapevine (Vitis spp.) Organs and in Their Hydraulic and Non-Hydraulic Interactions at the Whole-Plant Level: A Physiological and Molecular Update. Funct. Plant Biol. 2010, 37, 98–116. [Google Scholar] [CrossRef]
- Carvalho, L.C.; Gonçalves, E.F.; Marques da Silva, J.; Costa, J.M. Potential Phenotyping Methodologies to Assess Inter- and Intravarietal Variability and to Select Grapevine Genotypes Tolerant to Abiotic Stress. Front. Plant Sci. 2021, 12, 718202. [Google Scholar] [CrossRef]
- Haider, M.S.; Kurjogi, M.M.; Khalil-Ur-Rehman, M.; Fiaz, M.; Pervaiz, T.; Jiu, S.; Haifeng, J.; Chen, W.; Fang, J. Grapevine Immune Signaling Network in Response to Drought Stress as Revealed by Transcriptomic Analysis. Plant Physiol. Biochem. 2017, 121, 187–195. [Google Scholar] [CrossRef]
- Lecourieux, F.; Kappel, C.; Pieri, P.; Charon, J.; Pillet, J.; Hilbert, G.; Renaud, C.; Gomès, E.; Delrot, S.; Lecourieux, D. Dissecting the Biochemical and Transcriptomic Effects of a Locally Applied Heat Treatment on Developing Cabernet Sauvignon Grape Berries. Front Plant Sci. 2017, 8, 53. [Google Scholar] [CrossRef]
- Rienth, M.; Torregrosa, L.; Luchaire, N.; Chatbanyong, R.; Lecourieux, D.; Kelly, M.T.; Romieu, C. Day and Night Heat Stress Trigger Different Transcriptomic Responses in Green and Ripening Grapevine (Vitis vinifera) Fruit. BMC Plant Biol. 2014, 14, 108. [Google Scholar] [CrossRef]
- Rocheta, M.; Becker, J.D.; Coito, J.L.; Carvalho, L.; Amâncio, S. Heat and Water Stress Induce Unique Transcriptional Signatures of Heat-Shock Proteins and Transcription Factors in Grapevine. Funct. Integr. Genom. 2014, 14, 135–148. [Google Scholar] [CrossRef]
- Savoi, S.; Wong, D.C.J.; Arapitsas, P.; Miculan, M.; Bucchetti, B.; Peterlunger, E.; Fait, A.; Mattivi, F.; Castellarin, S.D. Transcriptome and Metabolite Profiling Reveals That Prolonged Drought Modulates the Phenylpropanoid and Terpenoid Pathway in White Grapes (Vitis vinifera, L.). BMC Plant Biol. 2016, 16, 67. [Google Scholar] [CrossRef] [Green Version]
- Camps, C.; Kappel, C.; Lecomte, P.; Léon, C.; Gomès, E.; Coutos-Thévenot, P.; Delrot, S. A Transcriptomic Study of Grapevine (Vitis vinifera cv. Cabernet-Sauvignon) Interaction with the Vascular Ascomycete Fungus Eutypa lata. J. Exp. Bot. 2010, 61, 1719–1737. [Google Scholar] [CrossRef]
- Lemaitre-Guillier, C.; Fontaine, F.; Roullier-Gall, C.; Harir, M.; Magnin-Robert, M.; Clément, C.; Trouvelot, S.; Gougeon, R.D.; Schmitt-Kopplin, P.; Adrian, M. Cultivar- and Wood Area-Dependent Metabolomic Fingerprints of Grapevine Infected by Botryosphaeria Dieback. Phytopathology 2020, 110, 1821–1837. [Google Scholar] [CrossRef] [PubMed]
- Magnin-Robert, M.; Spagnolo, A.; Boulanger, A.; Joyeux, C.; Clément, C.; Abou-Mansour, E.; Fontaine, F. Changes in Plant Metabolism and Accumulation of Fungal Metabolites in Response to Esca Proper and Apoplexy Expression in the Whole Grapevine. Phytopathology 2016, 106, 541–553. [Google Scholar] [CrossRef] [PubMed]
- Magnin-Robert, M.; Adrian, M.; Trouvelot, S.; Spagnolo, A.; Jacquens, L.; Letousey, P.; Rabenoelina, F.; Harir, M.; Roullier-Gall, C.; Clément, C.; et al. Alterations in Grapevine Leaf Metabolism Occur Prior to Esca Apoplexy Appearance. Mol. Plant-Microbe Interact. 2017, 30, 946–959. [Google Scholar] [CrossRef] [PubMed]
- Spagnolo, A.; Magnin-Robert, M.; Alayi, T.D.; Cilindre, C.; Mercier, L.; Schaeffer-Reiss, C.; Van Dorsselaer, A.; Clément, C.; Fontaine, F. Physiological Changes in Green Stems of Vitis vinifera, L. cv. Chardonnay in Response to Esca Proper and Apoplexy Revealed by Proteomic and Transcriptomic Analyses. J. Proteome Res. 2012, 11, 461–475. [Google Scholar] [CrossRef] [PubMed]
- Czemmel, S.; Galarneau, E.R.; Travadon, R.; McElrone, A.J.; Cramer, G.R.; Baumgartner, K. Genes Expressed in Grapevine Leaves Reveal Latent Wood Infection by the Fungal Pathogen Neofusicoccum parvum. PLoS ONE 2015, 10, e0121828. [Google Scholar] [CrossRef]
- Massonnet, M.; Figueroa-Balderas, R.; Galarneau, E.R.A.; Miki, S.; Lawrence, D.P.; Sun, Q.; Wallis, C.M.; Baumgartner, K.; Cantu, D. Neofusicoccum parvum Colonization of the Grapevine Woody Stem Triggers Asynchronous Host Responses at the Site of Infection and in the Leaves. Front Plant Sci. 2017, 8, 1117. [Google Scholar] [CrossRef]
- Koussevitzky, S.; Suzuki, N.; Huntington, S.; Armijo, L.; Sha, W.; Cortes, D.; Shulaev, V.; Mittler, R. Ascorbate Peroxidase 1 Plays a Key Role in the Response of Arabidopsis thaliana to Stress Combination. J. Biol. Chem. 2008, 283, 34197–34203. [Google Scholar] [CrossRef]
- Rizhsky, L.; Liang, H.; Shuman, J.; Shulaev, V.; Davletova, S.; Mittler, R. When Defense Pathways Collide. The Response of Arabidopsis to a Combination of Drought and Heat Stress. Plant Physiol. 2004, 134, 1683–1696. [Google Scholar] [CrossRef]
- Zandalinas, S.I.; Mittler, R.; Balfagón, D.; Arbona, V.; Gómez-Cadenas, A. Plant Adaptations to the Combination of Drought and High Temperatures. Physiol. Plant. 2018, 162, 2–12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Galarneau, E.R.A.; Lawrence, D.P.; Travadon, R.; Baumgartner, K. Drought Exacerbates Botryosphaeria Dieback Symptoms in Grapevines and Confounds Host-Based Molecular Markers of Infection by Neofusicoccum parvum. Plant Dis. 2019, 103, 1738–1745. [Google Scholar] [CrossRef] [PubMed]
- Lima, M.R.M.; Felgueiras, M.L.; Cunha, A.; Chicau, G.; Ferreres, F.; Dias, A.C.P. Differential Phenolic Production in Leaves of Vitis vinifera cv. Alvarinho Affected with Esca Disease. Plant Physiol. Biochem. 2017, 112, 45–52. [Google Scholar] [CrossRef] [PubMed]
- Reis, P.; Pierron, R.; Larignon, P.; Lecomte, P.; Abou-Mansour, E.; Farine, S.; Bertsch, C.; Jacques, A.; Trotel-Aziz, P.; Rego, C.; et al. Vitis Methods to Understand and Develop Strategies for Diagnosis and Sustainable Control of Grapevine Trunk Diseases. Phytopathology 2019, 109, 916–931. [Google Scholar] [CrossRef] [PubMed]
- Reis, P.; Magnin-Robert, M.; Nascimento, T.; Spagnolo, A.; Abou-Mansour, E.; Fioretti, C.; Clément, C.; Rego, C.; Fontaine, F. Reproducing Botryosphaeria Dieback Foliar Symptoms in a Simple Model System. Plant Dis. 2016, 100, 1071–1079. [Google Scholar] [CrossRef]
- Liu, M.; Ju, Y.; Min, Z.; Fang, Y.; Meng, J. Transcriptome Analysis of Grape Leaves Reveals Insights into Response to Heat Acclimation. Sci. Hortic. 2020, 272, 109554. [Google Scholar] [CrossRef]
- Zha, Q.; Xi, X.; He, Y.; Jiang, A. Transcriptomic Analysis of the Leaves of Two Grapevine Cultivars under High-Temperature Stress. Sci. Hortic. 2020, 265, 109265. [Google Scholar] [CrossRef]
- Toffolatti, S.L.; De Lorenzis, G.; Brilli, M.; Moser, M.; Shariati, V.; Tavakol, E.; Maddalena, G.; Passera, A.; Casati, P.; Pindo, M.; et al. Novel Aspects on The Interaction Between Grapevine and Plasmopara viticola: Dual-RNA-Seq Analysis Highlights Gene Expression Dynamics in The Pathogen and The Plant During the Battle for Infection. Genes 2020, 11, 261. [Google Scholar] [CrossRef]
- Kopecky, D.; Matušíková, I.; Sziderics, A.H.; Trognitz, F.; Spieß, N.; Stierschneider, M.; Fluch, S. In Silico Search for Drought-Responsive Genes in Plants on the Basis of Scientific Data: Case Study on Poplar Roots. Acta Physiol. Plant. 2013, 35, 1955–1966. [Google Scholar] [CrossRef]
- Hao, L.; Liu, X.; Zhang, X.; Sun, B.; Liu, C.; Zhang, D.; Tang, H.; Li, C.; Li, Y.; Shi, Y.; et al. Genome-Wide Identification and Comparative Analysis of Drought Related Genes in Roots of Two Maize Inbred Lines with Contrasting Drought Tolerance by RNA Sequencing. J. Integr. Agric. 2020, 19, 449–464. [Google Scholar] [CrossRef]
- Santibanez, C. Comparative Genetic and Metabolic Characterization between Two Table Grape Varieties with Contrasted Color Berry Skin: Red Globe and Chimenti Globe. Ph.D. Thesis, Université de Bordeaux & La Pontificia Universidad de Chile, Bordeaux, France, 2017. [Google Scholar]
- Díaz-Riquelme, J.; Zhurov, V.; Rioja, C.; Pérez-Moreno, I.; Torres-Pérez, R.; Grimplet, J.; Carbonell-Bejerano, P.; Bajda, S.; Van Leeuwen, T.; Martínez-Zapater, J.M.; et al. Comparative Genome-Wide Transcriptome Analysis of Vitis vinifera Responses to Adapted and Non-Adapted Strains of Two-Spotted Spider Mite, Tetranychus urticae. BMC Genom. 2016, 17, 74. [Google Scholar] [CrossRef] [Green Version]
- He, L.; Xu, X.-Q.; Wang, Y.; Chen, W.-K.; Sun, R.-Z.; Cheng, G.; Liu, B.; Chen, W.; Duan, C.-Q.; Wang, J.; et al. Modulation of Volatile Compound Metabolome and Transcriptome in Grape Berries Exposed to Sunlight under Dry-Hot Climate. BMC Plant Biol. 2020, 20, 59. [Google Scholar] [CrossRef]
- Wang, M.; Sun, X.; Wang, C.; Cui, L.; Chen, L.; Zhang, C.; Shangguan, L.; Fang, J. Characterization of MiR061 and Its Target Genes in Grapevine Responding to Exogenous Gibberellic Acid. Funct. Integr. Genom. 2017, 17, 537–549. [Google Scholar] [CrossRef]
- Ju, Y.; Min, Z.; Zhang, Y.; Zhang, K.; Liu, M.; Fang, Y. Transcriptome Profiling Provide New Insights into the Molecular Mechanism of Grapevine Response to Heat, Drought, and Combined Stress. Sci. Hortic. 2021, 286, 110076. [Google Scholar] [CrossRef]
- Berens, M.L.; Berry, H.M.; Mine, A.; Argueso, C.T.; Tsuda, K. Evolution of Hormone Signaling Networks in Plant Defense. Annu. Rev. Phytopathol. 2017, 55, 401–425. [Google Scholar] [CrossRef]
- Bezrutczyk, M.; Yang, J.; Eom, J.-S.; Prior, M.; Sosso, D.; Hartwig, T.; Szurek, B.; Oliva, R.; Vera-Cruz, C.; White, F.F.; et al. Sugar Flux and Signaling in Plant-Microbe Interactions. Plant J. 2018, 93, 675–685. [Google Scholar] [CrossRef]
- Abou-Mansour, E.; Débieux, J.-L.; Ramírez-Suero, M.; Bénard-Gellon, M.; Magnin-Robert, M.; Spagnolo, A.; Chong, J.; Farine, S.; Bertsch, C.; L’Haridon, F.; et al. Phytotoxic Metabolites from Neofusicoccum parvum, a Pathogen of Botryosphaeria Dieback of Grapevine. Phytochemistry 2015, 115, 207–215. [Google Scholar] [CrossRef]
- Labois, C.; Wilhelm, K.; Laloue, H.; Tarnus, C.; Bertsch, C.; Goddard, M.-L.; Chong, J. Wood Metabolomic Responses of Wild and Cultivated Grapevine to Infection with Neofusicoccum parvum, a Trunk Disease Pathogen. Metabolites 2020, 10, 232. [Google Scholar] [CrossRef]
- Griesser, M.; Weingart, G.; Schoedl-Hummel, K.; Neumann, N.; Becker, M.; Varmuza, K.; Liebner, F.; Schuhmacher, R.; Forneck, A. Severe Drought Stress Is Affecting Selected Primary Metabolites, Polyphenols, and Volatile Metabolites in Grapevine Leaves (Vitis vinifera cv. Pinot Noir). Plant Physiol. Biochem. 2015, 88, 17–26. [Google Scholar] [CrossRef]
- Hochberg, U.; Degu, A.; Toubiana, D.; Gendler, T.; Nikoloski, Z.; Rachmilevitch, S.; Fait, A. Metabolite Profiling and Network Analysis Reveal Coordinated Changes in Grapevine Water Stress Response. BMC Plant Biol. 2013, 13, 184. [Google Scholar] [CrossRef]
- Cramer, G.R.; Ergül, A.; Grimplet, J.; Tillett, R.L.; Tattersall, E.A.R.; Bohlman, M.C.; Vincent, D.; Sonderegger, J.; Evans, J.; Osborne, C.; et al. Water and Salinity Stress in Grapevines: Early and Late Changes in Transcript and Metabolite Profiles. Funct. Integr. Genom. 2007, 7, 111–134. [Google Scholar] [CrossRef]
- Bettenfeld, P.; Cadena i Canals, J.; Jacquens, L.; Fernandez, O.; Fontaine, F.; van Schaik, E.; Courty, P.-E.; Trouvelot, S. The Microbiota of the Grapevine Holobiont: A Key Component of Plant Health. J. Adv. Res. 2021, 40, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Leal, C.; Richet, N.; Guise, J.-F.; Gramaje, D.; Armengol, J.; Fontaine, F.; Trotel-Aziz, P. Cultivar Contributes to the Beneficial Effects of Bacillus subtilis PTA-271 and Trichoderma atroviride SC1 to Protect Grapevine Against Neofusicoccum parvum. Front Microbiol. 2021, 12, 726132. [Google Scholar] [PubMed]
- Trotel-Aziz, P.; Abou-Mansour, E.; Courteaux, B.; Rabenoelina, F.; Clément, C.; Fontaine, F.; Aziz, A. Bacillus SubtilisPTA-271 Counteracts Botryosphaeria Dieback in Grapevine, Triggering Immune Responses and Detoxification of Fungal Phytotoxins. Front. Plant Sci. 2019, 10, 25. [Google Scholar] [CrossRef] [PubMed]
- Saijo, Y.; Loo, E.P. Plant Immunity in Signal Integration between Biotic and Abiotic Stress Responses. New Phytol. 2020, 225, 87–104. [Google Scholar] [CrossRef]
- Chong, J.; Poutaraud, A.; Hugueney, P. Metabolism and Roles of Stilbenes in Plants. Plant Sci. 2009, 177, 143–155. [Google Scholar] [CrossRef]
- Jeandet, P.; Clément, C.; Courot, E. Resveratrol Production at Large Scale Using Plant Cell Suspensions. Eng. Life Sci. 2014, 14, 622–632. [Google Scholar] [CrossRef]
- Ku, Y.-S.; Sintaha, M.; Cheung, M.-Y.; Lam, H.-M. Plant Hormone Signaling Crosstalks between Biotic and Abiotic Stress Responses. Int. J. Mol. Sci. 2018, 19, 3206. [Google Scholar] [CrossRef]
- Rizhsky, L.; Liang, H.; Mittler, R. The Combined Effect of Drought Stress and Heat Shock on Gene Expression in Tobacco. Plant Physiol. 2002, 130, 1143–1151. [Google Scholar] [CrossRef]
- Calvo-Garrido, C.; Songy, A.; Marmol, A.; Roda, R.; Clément, C.; Fontaine, F. Description of the Relationship between Trunk Disease Expression and Meteorological Conditions, Irrigation and Physiological Response in Chardonnay Grapevines. OENO One 2021, 55, 97–113. [Google Scholar] [CrossRef]
- Lebon, E.; Duchêne, E.; Brun, O.; Clément, C. Phenology of Flowering and Starch Accumulation in Grape (Vitis vinifera, L.) Cuttings and Vines. Ann. Bot. 2005, 95, 943–948. [Google Scholar] [CrossRef] [PubMed]
- Spagnolo, A.; Mondello, V.; Larignon, P.; Villaume, S.; Rabenoelina, F.; Clément, C.; Fontaine, F.; Spagnolo, A.; Mondello, V.; Larignon, P.; et al. Defense Responses in Grapevine (cv. Mourvèdre) after Inoculation with the Botryosphaeria Dieback Pathogens Neofusicoccum parvum and Diplodia seriata and Their Relationship with Flowering. Int. J. Mol. Sci. 2017, 18, 393. [Google Scholar] [CrossRef] [PubMed]
- Larignon, P.; Dubos, B. Fungi Associated with Esca Disease in Grapevine. Eur. J. Plant Pathol. 1997, 103, 147–157. [Google Scholar] [CrossRef]
- Andrews, S. FastQC: A Quality Control Tool for High Throughput Sequence Data. Available online: https://www.bioinformatics.babraham.ac.uk/projects/fastqc (accessed on 1 March 2018).
- Martin, M. Cutadapt Removes Adapter Sequences from High-Throughput Sequencing Reads. EMBnet. J. 2011, 17, 10–12. [Google Scholar] [CrossRef]
- Jaillon, O.; Aury, J.-M.; Noel, B.; Policriti, A.; Clepet, C.; Casagrande, A.; Choisne, N.; Aubourg, S.; Vitulo, N.; Jubin, C.; et al. The Grapevine Genome Sequence Suggests Ancestral Hexaploidization in Major Angiosperm Phyla. Nature 2007, 449, 463–467. [Google Scholar] [CrossRef] [PubMed]
- Vitulo, N.; Forcato, C.; Carpinelli, E.C.; Telatin, A.; Campagna, D.; D’Angelo, M.; Zimbello, R.; Corso, M.; Vannozzi, A.; Bonghi, C.; et al. A Deep Survey of Alternative Splicing in Grape Reveals Changes in the Splicing Machinery Related to Tissue, Stress Condition and Genotype. BMC Plant Biol. 2014, 14, 99. [Google Scholar] [CrossRef]
- Patro, R.; Duggal, G.; Love, M.I.; Irizarry, R.A.; Kingsford, C. Salmon Provides Fast and Bias-Aware Quantification of Transcript Expression. Nat. Methods 2017, 14, 417–419. [Google Scholar] [CrossRef]
- Soneson, C.; Love, M.I.; Robinson, M.D. Differential Analyses for RNA-Seq: Transcript-Level Estimates Improve Gene-Level Inferences. F1000Research 2015, 4, 1521. [Google Scholar] [CrossRef]
- Robinson, M.D.; McCarthy, D.J.; Smyth, G.K. EdgeR: A Bioconductor Package for Differential Expression Analysis of Digital Gene Expression Data. Bioinformatics 2010, 26, 139–140. [Google Scholar] [CrossRef]
- Haas, B.J.; Papanicolaou, A.; Yassour, M.; Grabherr, M.; Blood, P.D.; Bowden, J.; Couger, M.B.; Eccles, D.; Li, B.; Lieber, M.; et al. De Novo Transcript Sequence Reconstruction from RNA-Seq Using the Trinity Platform for Reference Generation and Analysis. Nat. Protoc. 2013, 8, 1494–1512. [Google Scholar] [CrossRef]
- Benjamini, Y.; Hochberg, Y. Controlling the False Discovery Rate: A Practical and Powerful Approach to Multiple Testing. J. R. Stat. Soc. Ser. B Methodol. 1995, 57, 289–300. [Google Scholar] [CrossRef]
- Tian, T.; Liu, Y.; Yan, H.; You, Q.; Yi, X.; Du, Z.; Xu, W.; Su, Z. AgriGO v2.0: A GO Analysis Toolkit for the Agricultural Community, 2017 Update. Nucleic Acids Res. 2017, 45, W122–W129. [Google Scholar] [CrossRef] [PubMed]
- Bézier, A.; Lambert, B.; Baillieul, F. Study of Defense-Related Gene Expression in Grapevine Leaves and Berries Infected with Botrytis cinerea. Eur. J. Plant Pathol. 2002, 108, 111–120. [Google Scholar] [CrossRef]
- Hellemans, J.; Mortier, G.; De Paepe, A.; Speleman, F.; Vandesompele, J. QBase Relative Quantification Framework and Software for Management and Automated Analysis of Real-Time Quantitative PCR Data. Genome Biol. 2007, 8, R19. [Google Scholar] [CrossRef] [PubMed]
- Adrian, M.; Lucio, M.; Roullier-Gall, C.; Héloir, M.-C.; Trouvelot, S.; Daire, X.; Kanawati, B.; Lemaître-Guillier, C.; Poinssot, B.; Gougeon, R.; et al. Metabolic Fingerprint of PS3-Induced Resistance of Grapevine Leaves against Plasmopara viticola Revealed Differences in Elicitor-Triggered Defenses. Front. Plant Sci. 2017, 8, 101. [Google Scholar] [CrossRef]
- Gougeon, R.D.; Lucio, M.; Frommberger, M.; Peyron, D.; Chassagne, D.; Alexandre, H.; Feuillat, F.; Voilley, A.; Cayot, P.; Gebefügi, I.; et al. The Chemodiversity of Wines Can Reveal a Metabologeography Expression of Cooperage Oak Wood. Proc. Natl. Acad. Sci. USA 2009, 106, 9174–9179. [Google Scholar] [CrossRef]
- Tziotis, D.; Hertkorn, N.; Schmitt-Kopplin, P. Kendrick-Analogous Network Visualisation of Ion Cyclotron Resonance Fourier Transform Mass Spectra: Improved Options for the Assignment of Elemental Compositions and the Classification of Organic Molecular Complexity. Eur. J. Mass Spectrom. 2011, 17, 415–421. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| A—All Necrosis Length Data Combined | Degree of Freedom | Sum of Square | Square Mean | F-Value | p-Value | Significance |
| Factor 1: infection | 2 | 0.07289 | 0.03644 | 47.843 | 4.23 × 10−9 | *** |
| Faction 2: stress | 3 | 0.00359 | 0.0012 | 1.572 | 0.222 | |
| Interaction (infection × stress) | 6 | 0.01235 | 0.00206 | 2.701 | 0.0379 | * |
| Residuals | 24 | 0.01828 | 0.00076 | |||
| Coefficients: | Estimate | Std. Error | t Value | p-Value | Significance | |
| (Intercept) | 0.153963 | 0.015935 | 9.662 | 9.58 × 10−10 | *** | |
| stressWS | −0.040229 | 0.022535 | −1.785 | 0.0869 | ||
| stressHS | 0.014863 | 0.022535 | 0.66 | 0.5158 | ||
| stressWSHS | 0.016368 | 0.022535 | 0.726 | 0.4747 | ||
| infectionDs | −0.023705 | 0.022535 | −1.052 | 0.3033 | ||
| infectionNp | −0.105748 | 0.022535 | −4.693 | 9.07 × 10−5 | *** | |
| stressWS:infectionDs | 0.005667 | 0.031869 | 0.178 | 0.8604 | ||
| stressHS:infectionDs | −0.060717 | 0.031869 | −1.905 | 0.0688 | ||
| stressWSHS:infectionDs | −0.088933 | 0.031869 | −2.791 | 0.0101 | * | |
| stressWS:infectionNp | 0.04796 | 0.031869 | 1.505 | 0.1454 | ||
| stressHS:infectionNp | −0.027043 | 0.031869 | −0.849 | 0.4045 | ||
| stressWSHS:infectionNp | −0.038271 | 0.031869 | −1.201 | 0.2415 | ||
| B—Only Np Infected vs. Control Values | Degree of Freedom | Sum of Square | Square Mean | F-Value | p-Value | Significance |
| Factor 1: infection | 1 | 0.07271 | 0.07271 | 91.219 | 5.19 × 10−8 | *** |
| Faction 2: stress | 3 | 0.00117 | 0.00039 | 0.49 | 0.694 | |
| Interaction (infection × stress) | 3 | 0.00663 | 0.00221 | 2.773 | 0.0753 | |
| Residuals | 16 | 0.01275 | 0.0008 | |||
| C—Only Np infected vs. Control Values | Degree of Freedom | Sum of Square | Square Mean | F-Value | p-Value | Significance |
| Factor 1: infection | 1 | 0.021385 | 0.021385 | 24.586 | 0.000142 | *** |
| Faction 2: stress | 3 | 0.004729 | 0.001576 | 1.812 | 0.185496 | |
| Interaction (infection × stress) | 3 | 0.009667 | 0.003222 | 3.705 | 0.033785 | * |
| Residuals | 16 | 0.013917 | 0.00087 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fernandez, O.; Lemaître-Guillier, C.; Songy, A.; Robert-Siegwald, G.; Lebrun, M.-H.; Schmitt-Kopplin, P.; Larignon, P.; Adrian, M.; Fontaine, F. The Combination of Both Heat and Water Stresses May Worsen Botryosphaeria Dieback Symptoms in Grapevine. Plants 2023, 12, 753. https://doi.org/10.3390/plants12040753
Fernandez O, Lemaître-Guillier C, Songy A, Robert-Siegwald G, Lebrun M-H, Schmitt-Kopplin P, Larignon P, Adrian M, Fontaine F. The Combination of Both Heat and Water Stresses May Worsen Botryosphaeria Dieback Symptoms in Grapevine. Plants. 2023; 12(4):753. https://doi.org/10.3390/plants12040753
Chicago/Turabian StyleFernandez, Olivier, Christelle Lemaître-Guillier, Aurélie Songy, Guillaume Robert-Siegwald, Marc-Henri Lebrun, Philippe Schmitt-Kopplin, Philippe Larignon, Marielle Adrian, and Florence Fontaine. 2023. "The Combination of Both Heat and Water Stresses May Worsen Botryosphaeria Dieback Symptoms in Grapevine" Plants 12, no. 4: 753. https://doi.org/10.3390/plants12040753