
Ecogeographic Study of Ipomoea Species in Mauritius, Indian Ocean

 ,
,  , and
, and
Abstract
:1. Introduction
2. Results
2.1. Inventory of Ipomoea Species
2.2. Field Study
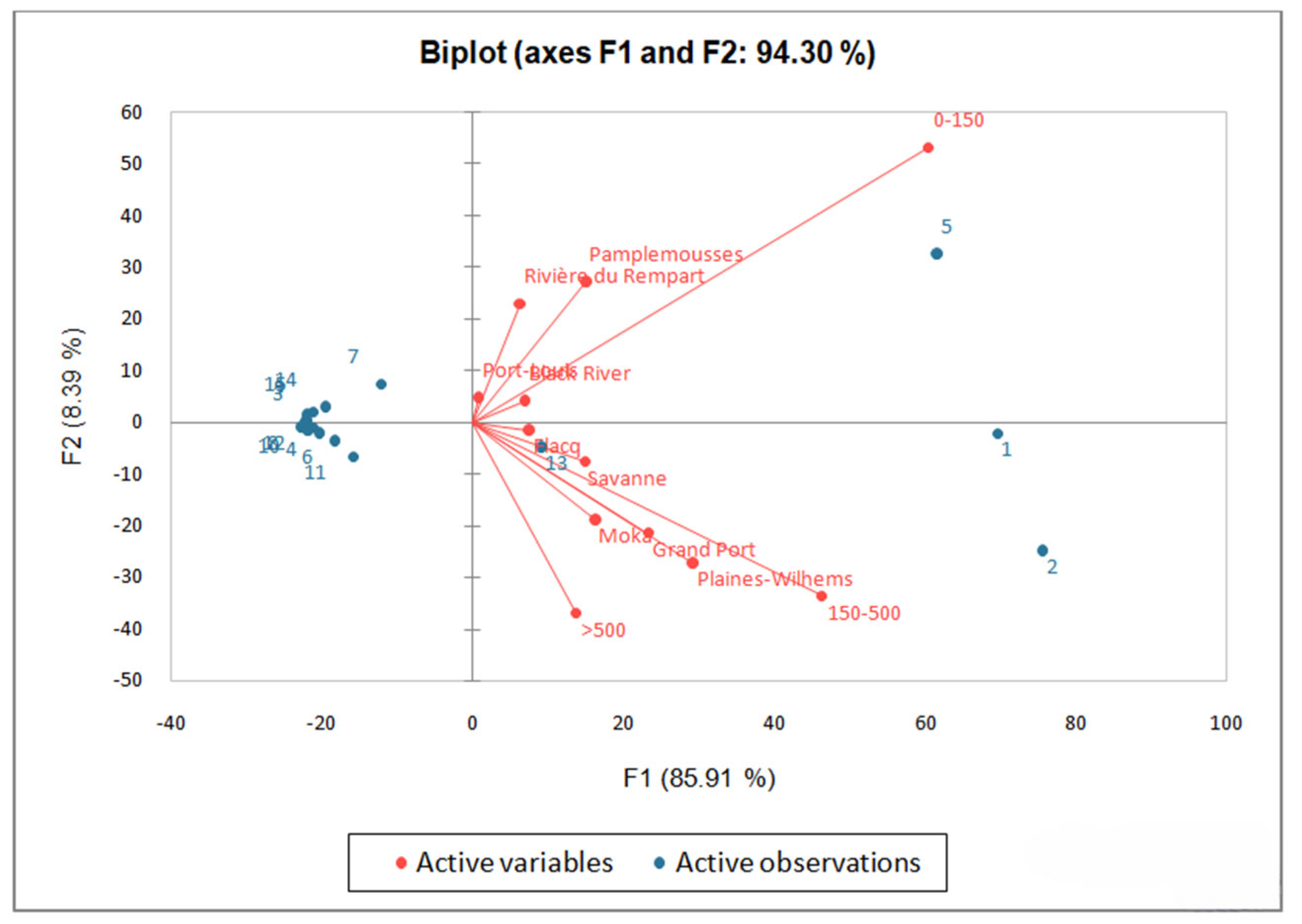
2.3. Distribution Patterns of Ipomoea Species
2.4. Relationship between Species Distribution and Geographical Area
2.5. Regional Richness
2.6. Ecogeographic Conspectus
2.6.1. Ipomoea Alba
2.6.2. Ipomoea Aquatica
2.6.3. Ipomoea Batatas
2.6.4. Ipomoea Cairica
2.6.5. Ipomoea Carnea
2.6.6. Ipomoea Herderifolia
2.6.7. Ipomoea Indica
2.6.8. Ipomoea Littoralis
2.6.9. Ipomoea Nil
2.6.10. Ipomoea Obscura
2.6.11. Ipomoea Ochracea
2.6.12. Ipomoea Pes-Caprae
2.6.13. Ipomoea Purpurea
2.6.14. Ipomoea Quamoclit
2.6.15. Ipomoea Tiliacea
2.6.16. Ipomoea Triloba
3. Materials and Methods
3.1. Study Area
3.2. Ecogeography Study Methodology
3.2.1. Identification of Taxon
3.2.2. Fieldwork
3.2.3. The Ecogeographic Database
3.3. Statistical Analysis
4. Discussion
4.1. Distribution Pattern
4.2. Management and Control
4.3. Conservation
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- IPBES. Global Assessment Report of the Intergovernmental Science-Policy Platform on Biodiversity and Ecosystem Services; Brondízio, E.S., Settele, J., Díaz, S., Ngo, H.T., Eds.; IPBES Secretariat: Bonn, Germany, 2019. [Google Scholar]
- FAO. The State of the World’s Biodiversity for Food and Agriculture; Bélanger, J., Pilling, D., Eds.; FAO Commission on Genetic Resources for Food and Agriculture Assessments: Rome, Italy, 2019. [Google Scholar]
- Dulloo, M.E.; Bissessur, P.; Rana, J. Monitoring plant genetic diversity for food and agriculture. In Plant Genetic Resources: A Review of Current Research and Future Needs; Dulloo, M.E., Ed.; Burleigh Dodds Science Publishing: Cambridge, UK, 2021; ISBN 978-1-78676-451-5. [Google Scholar]
- Salgotra, R.K.; Chauhan, B.S. Genetic Diversity, Conservation, and Utilization of Plant Genetic Resources. Genes 2023, 14, 174. [Google Scholar] [CrossRef] [PubMed]
- Ebert, A.W.; Engels, J.M.M. Plant Biodiversity and Genetic Resources Matter. Plants 2020, 9, 1706. [Google Scholar] [CrossRef]
- Dulloo, M.E.; Thormann, I.; Drucker, A.G. What do we have to lose? Monitoring crop genetic diversity. In Enhancing Crop Genepool Use: Capturing Wild Relative and Landrace Diversity for Crop Improvement; Maxted, N., Dulloo, M.E., Ford-Lloyd, B.V., Eds.; CABI: Oxfordshire, UK, 2016; pp. 421–435. ISBN 978-1-78064-613-8. [Google Scholar]
- Ramírez-Villegas, J.; Khoury, C.K.; Jarvis, A.; Debouck, D.; Guarino, L. A Gap Analysis Methodology for Collecting Crop Genepools: A Case Study with Phaseolus Beans. PLoS ONE 2010, 5, e13497. [Google Scholar] [CrossRef] [PubMed]
- Jarvis, A.; Lane, A.; Hijmans, R.J. The effect of climate change on crop wild relatives. Agric. Ecosyst. Environ. 2008, 126, 13–23. [Google Scholar] [CrossRef]
- Vaslet, S. Project Introduction: Germination. Biodiversité & Agriculture Océan Indien; CIRAD, Pôle de protection des plantes: St Pierre, Réunion, 2021. [Google Scholar]
- Farooq, S.; El-Azam, F. Co-Existence of salt and drought tolerance in Triticeae. Hereditas 2001, 135, 205–210. [Google Scholar] [CrossRef]
- De Hann, S. Community based conservation of crop genetic resources. In Plant Genetic Resources: A Review of Current Research and Future Needs, 1st ed.; Dulloo, M.E., Ed.; Burleigh Dodds Science Publishing: Cambridge, UK, 2021; ISBN 978-1-78676-451-5. [Google Scholar]
- Guarino, L.; Maxted, N.; Sawkins, M. Analysis of geo-referenced data and the conservation and use of plant genetic resources. In Proceedings of the Linking Genetics and Geography: Emerging Strategies for Managing Crop Biodiversity ASA/CSSA/SSSA Annual Meeting, Anaheim, CA, USA, 26–31 October 1997. [Google Scholar]
- Dulloo, M.E.; Bullock, D.J.; North, S.G. Report on the Expedition to Round Island and Gunner’s Quoin, Mauritius July/August 1996; Mauritian Wildlife Foundation: Port Louis, Mauritius, 1999; p. 62. [Google Scholar]
- Maxted, N. Ecogeography and genetic conservation. In Genetic Resources of Mediterranean Pasture and Forage Legumes. Current Plant Science and Biotechnology in Agriculture; Bennett, S.J., Cocks, P.S., Eds.; Springer: Dordrecht, The Netherlands, 1999; Volume 33, pp. 53–66. [Google Scholar] [CrossRef]
- Álvarez, N.C.; Vincent, H.A.; Kell, S.P.; Eastwood, R.J.; Maxted, N. Ecogeographic surveys. In Collecting Plant Genetic Diversity: Technical Guidelines; Guarino, L., Rao, V.R., Goldberg, E., Eds.; Bioversity International: Rome, Italy, 2011; Available online: http://cropgenebank.sgrp.cgiar.org/index.php?option=com_content&view=article&id=652&Itemid=864&lang=English (accessed on 13 November 2023).
- Tohme, J.; Jones, P.; Beebe, S.; Iwanaga, M. The combined use of agroecological and characterisation data to establish the CIAT Phaseolus vulgaris core collection. In Core Collections of Plant Genetic Resources; Hodgkin, T., Brown, A.H.D., van Hintum, T.J.L., Morales, E.A.V., Eds.; Core Collections of Plant Genetic Resources, IBPGR; John Wiley and Sons: Chichester, UK, 1995; pp. 95–107. [Google Scholar]
- Parra-Quijano, M.; Iriondo, J.M.; Torres, E. Applications of ecogeography and geographic information systems in conservation and utilization of plant genetic resources. Span. J. Agric. Res. 2012, 10, 419–429. [Google Scholar] [CrossRef]
- Zuriaga, E.; Blanca, J.M.; Cordero, L.; Sifres, A.; Blas-Cerdán, W.G.; Morales, R.; Nuez, F. Genetic and bioclimatic variation in Solanum pimpinellifolium. Genet. Resour. Crop Evol. 2009, 56, 39–51. [Google Scholar] [CrossRef]
- Ehrman, T.; Cocks, P.S. Ecogeographic survey of wild annual legumes in Syria: Distribution patterns. J. Appl. Ecol. 1990, 27, 578–591. [Google Scholar] [CrossRef]
- Bothmer, R.V.; Jacobsen, N.; Baden, C.; Bagger Jørgensen, R.; Linde-Laursen, I. An Ecogeographical Study of the Genus Hordeum; Systematic and Ecogeographic Studies on Crop Genepools, 7; IBPGR: Rome, Italy, 1991; p. 127. [Google Scholar]
- Maxted, N.; van Slageren, M.W.; Rihan, J.R. Ecogeographic surveys. In Collecting Plant Genetic Diversity, Technical Guidelines; Guarino, L., Ramanatha Rao, V., Reid, R., Eds.; CAB International: Wallingford, UK, 1995; pp. 255–285. [Google Scholar]
- Maxted, N.; Dulloo, E.; Ford-Lloyd, B.V.; Iriondo, J.M.; Jarvis, A. Gap analysis: A tool for complementary genetic conservation assessment. Divers. Distrib. 2008, 14, 1018–1030. [Google Scholar] [CrossRef]
- Batchu, A.K.; Zimmermann, D.; Schulze-Lefert, P.; Koprek, T. Correlation between hordatine accumulation, environmental factors and genetic diversity in wild barley (Hordeum spontaneum C. Koch) accessions from the Near East Fertile Crescent. Genetica 2006, 127, 87–99. [Google Scholar] [CrossRef]
- Bhullar, N.K.; Street, K.; Mackay, M.; Yahiaoui, N.; Keller, B. Unlocking wheat genetic resources for the molecular identification of previously undescribed functional alleles at the Pm3 resistance locus. Proc. Natl. Acad. Sci. USA 2009, 106, 9519–9524. [Google Scholar] [CrossRef] [PubMed]
- Lecours, V. Habitat mapping. In Encyclopedia of Ecology, 2nd ed.; Brian, D.F., Ed.; Elsevier Inc: Amsterdam, The Netherlands, 2019; pp. 212–222. ISBN 9780444641304. [Google Scholar] [CrossRef]
- Steiner, J.J.; Greene, S.L. Proposed ecological descriptors and their utility for plant germplasm collections. Crop Sci. 1996, 36, 439–451. [Google Scholar] [CrossRef]
- Dulloo, M.; Maxted, N.; Guarino, L.; Florens, D.; Bewbury, H.; Lloyd, B.F. Ecogeographic survey of the genus Coffea in the Mascarene Islands. Bot. J. Linn. 1999, 131, 263–284. [Google Scholar] [CrossRef]
- Al Beyrouthy, J.; Karam, N.; Al-Zein, M.S.; Yazbek, M. Ecogeographic survey and gap analysis for Medicago L.: Recommendations for in situ and ex situ conservation of Lebanese species. Genet. Resour. Crop Evol. 2019, 66, 1009–1026. [Google Scholar] [CrossRef]
- Wood, J.R.I.; Muñoz-Rodríguez, P.; Williams, B.R.M.; Scotland, R.W. A foundation monograph of Ipomoea (Convolvulaceae) in the New World. PhytoKeys 2020, 143, 1–823. [Google Scholar]
- Khoury, C.K.; Heider, B.; Castañeda-Álvarez, N.P.; Achicanoy, H.A.; Sosa, C.C.; Miller, R.E.; Scotland, R.W.; Wood, J.R.I.; Rossel, G.; Eserman, L.A.; et al. Distributions, ex situ conservation priorities, and genetic resource potential of crop wild relatives of sweetpotato [Ipomoea batatas (L.) Lam., I. series Batatas]. Front. Plant Sci. 2015, 6, 251. [Google Scholar] [CrossRef]
- Umekar, M.; Hemke, A.; Yadav, S. Convolvulaceae: A morning glory plant. Int. J. Pharm. Sci. Rev. Res. 2021, 51, 103–117. [Google Scholar]
- Bosser, J.; Heine, H. Convolvulaceae. In Flore des Mascareignes; Bosser, J., Ferguson, I.K., Soopramanien, C., Eds.; The Sugar Industry Research Institute, Mauritius, L’Institutde Recherche pour le Developpement (IRD): Paris, France; Royal Botanic Garden Kew: London, UK; Le Soutiende la Commissionde l’Océan Indien et de l’Union Europeenne: Port Louis, Mauritius, 2000; pp. 1–63. [Google Scholar]
- National Parks & Conservation Service; Ministry of Agro Industry and Food Security. Application for Extension and Renomination of Macchabee-Bel Ombre Biosphere Reserve. In Nomination Dossier. 2019. Available online: https://npcs.govmu.org/Documents/BLACK%20RIVER%20GORGES%20-%20BEL%20OMBRE%20NOMINATION%20DOSSIER%20-.pdf (accessed on 10 August 2024).
- Nimmakayala, P.; Vajja, G.; Reddy, U.K. Wild Crop Relatives: Genomic and Breeding Resources: Industrial Crops; Springer: Berlin/Heidelberg, Germany, 2011; pp. 123–132. [Google Scholar] [CrossRef]
- Barroso, A.A.M.; Ferreira, P.S.H.; Martins, D. Growth and Development of Ipomoea Weeds. Planta Daninha 2019, 37, e019186421. [Google Scholar] [CrossRef]
- Wood, J.; Carine, M.; Harris, D.; Wilkin, P.; Williams, B.; Scotland, R. Ipomoea (Convolvulaceae) in Bolivia. Kew Bull. 2015, 70, 31. [Google Scholar] [CrossRef]
- Srivastava, D.; Rauniyar, N. Medicinal Plants of Genus Ipomoea; LAP Lambert Academic Publisher: Beau Bassin, Mauritius, 2020. [Google Scholar]
- Gregory, P. (Ed.) Austin, The Taxonomy, Evolution and Genetic Diversity of Sweet Potatoes and Related Wild Species. In Exploration, Maintenance and Utilization of Sweet Potato Genetic Resources; CIP: Lima, Peru, 1987; pp. 27–59. [Google Scholar]
- Austin, D.F.; Huaman, Z. A synopsis of Ipomoea (Convolvulaceae) in the Americas. Taxon 1996, 45, 3–38. [Google Scholar] [CrossRef]
- Roullier, C.; Duputié, A.; Wennekes, P.; Benoit, L.; Bringas, V.M.F.; Rossel, G.; Tay, D.; McKey, D.; Lebot, V. Disentangling the origins of cultivated sweet potato (Ipomoea batatas (L.) Lam.). PLoS ONE 2013, 8, e62707. [Google Scholar] [CrossRef]
- Aguoru, C.U.; Uhia, P.; Olasan, J.O. Varietal Characterisation and Taxonomic Evaluation of Sweet Potato (Ipomoea batatas) Using Macro- and Micromorphological Evidence. Open Access Libr. J. OALIB 2015, 2, 1–7. [Google Scholar] [CrossRef]
- Stathers, T.; Low, J.; Mwanga, R.; Carey, T.; McEwan, M.; David, S.; Gibson, R.; Namanda, S.; McEwan, M.; Malinga, J.; et al. Everything You Ever Wanted to Know about Sweetpotato: Reaching Agents of Change ToT Manual; International Potato Center: Lima, Peru, 2018; Volume 12, p. 664. Available online: https://hdl.handle.net/10568/98334 (accessed on 15 November 2023).
- Woolfe, J.A. Sweet Potato: An Untapped Food Resource; Cambridge University Press: New York, NY, USA; Cambridge, UK, 1992; p. 643. [Google Scholar]
- Ministry of Agriculture and Natural Resources, Mauritius. Mauritius: Country Report to the FAO International Technical Conference on Plant Genetic Resources. 1996. Available online: https://www.fao.org/fileadmin/templates/agphome/documents/PGR/SoW1/asia/MAURITIU.PDF (accessed on 10 August 2024).
- CIP. Global Strategy for Ex-Situ Conservation of Sweetpotato Genetic Resources. Sweetpotato Genetic Resources Conservation–CIP Genebank November 2007. Available online: https://www.genebanks.org/resources/publications/sweetpotato-strategy-2007/ (accessed on 6 May 2024).
- Rouillard, G.; Guého, J. Convolvulacées. In Les Plantes et leur Histoire à l’Ile Maurice; MSM Press: Port Louis, Mauritius, 1999; pp. 363–368. [Google Scholar]
- Farooq, S.; Onen, H.; Ozaslan, C. Erken Tanı, Takip ve Bilgi Sistemi (Early Detection, Monitoring and Information System for Invasive Alien Plants). In Türkiye Istilacı itkiler Kataloğu, 1st ed.; Onen, G., Ed.; Gıda, Tarim ve Hayvancılık Bakanlığı Tarımsal Araştırmalar ve Politikalar Genel Müdürlüğü Bitki Sağlığı Araştırmaları Daire Başkanlığı: Istanbul, Turkey, 2015. [Google Scholar]
- Onen, H.; Farooq, S. Current Status and Future Prospects of Invasive Plants in Turkey. In Invasive Species in the Mediterranean; Watch Letter 33; CIHEAM: Paris, France, 2015; pp. 1–4. [Google Scholar]
- Onen, H.; Farooq, S.; Muñoz-Rodríguez, P.; Ali, A.S.; Alfarraj, S. Ipomoea tricolor (Convolvulaceae) in Turkey: New occurrence record and potential spread areas under current climatic conditions. J. King Saud Univ. Sci. 2023, 35, 102543. [Google Scholar] [CrossRef]
- Daruty de Grandpré, C. Plantes Medicinales de l’Ile Maurice et des Pays Intertropicaux; General steam Print. Co.: Port Louis, Mauritius, 1886. [Google Scholar]
- Gurib-Fakim, A.; Sewraj, M.; Gueho, J.; Dulloo, E. Medicalethnobotany of some weeds of Mauritius and Rodrigues. J. Ethnopharmacol. 1993, 39, 175–185. [Google Scholar] [CrossRef] [PubMed]
- Maimoni-Rodella, R.C.S.; Yanagizawa, Y.A.N.P. Floral biology and breeding system of three Ipomoea weeds. Planta Daninha 2007, 25, 35–42. [Google Scholar] [CrossRef]
- Hançerli, L.; Ayata, M.; Cakan, H.; Uygur, N.; Uygur, S. Türkiye Florası için Yeni Yabancı Ot Türü Kaydı Ipomoea hederifolia L. (Convolvulaceae) (A New Weed Species Record for the Flora of Turkey Ipomoea hederifolia L. (Convolvulaceae). Turk. J. Weed Sci. 2018, 21, 36–38. [Google Scholar]
- Onen, H.; Ozaslan, C.; Farooq, S.; Jones, C.A. Ipomoea coccinea L. (Convolvulaceae): A new introduced alien plant species in Turkey. EPPO Bull. 2021, 51, 207–212. [Google Scholar] [CrossRef]
- Truong, V.D.; Avula, R.Y.; Pecota, K.V.; Yencho, G.C. Sweetpotato production, processing, and nutritional quality. In Handbook of Vegetables and Vegetable Processing, 2nd ed.; Siddiq, M., Uebersax, M.A., Eds.; John Wiley & Sons, Ltd.: Hoboken, NJ, USA, 2018; Volume 2, pp. 811–837. [Google Scholar]
- Soto-Caro, A.; Luo, T.; Wu, F.; Guan, Z. The U.S. Sweet Potato Market: Price Response and Impact of Supply Shocks. Horticulturae 2022, 8, 856. [Google Scholar] [CrossRef]
- FAO. FAOSTAT, Food and Agriculture Data-Production Indices. 2022. Available online: https://www.fao.org/faostat/en/#data/QI (accessed on 11 May 2024).
- Zhang, P.; Fan, W.; Wang, H.; Wu, Y.; Zhou, W.; Yang, J. Developing new sweet potato varieties with improved performance. In Achieving Sustainable Cultivation of Potatoes; Wang-Pruski, G., Ed.; Burleigh Dodds Science Publishing: Cambridge, UK, 2018; p. 1. [Google Scholar] [CrossRef]
- Sivakumar, P.S. Sweet potato: A crops for food and nutritional security in Orissa. In Orissa Review; Odisha State Archives: Odisha, India, 2003; pp. 61–64. [Google Scholar]
- Reddy, R.; Soibam, H.; Ayam, V.S.; Panja, P.; Mitra, S. Morphological characterization of sweet potato cultivars during growth, development and harvesting. Indian J. Agric. Res. 2018, 52, 46–50. [Google Scholar] [CrossRef]
- Alam, M.K. A comprehensive review of sweet potato (Ipomoea batatas [L.] Lam): Revisiting the associated health benefits. Trends Food Sci. Technol. 2021, 115, 512–529. [Google Scholar] [CrossRef]
- AgMRC. 2021. Available online: https://www.agmrc.org/commodities-products/vegetables/sweet-potatoes (accessed on 11 May 2024).
- Statistics Mauritius. 2024. Available online: https://statsmauritius.govmu.org/Pages/Statistics/By_Subject/Agriculture/Arch_Agriculture.aspx (accessed on 11 May2024).
- Austin, D.F. The Ipomoea batatas complex-I. Taxonomy. Bull. Torrey Bot. Club 1978, 105, 114–129. [Google Scholar] [CrossRef]
- McDonald, J.A.; Austin, D.F. Changes and additions in ipomoea section batatas (Convolvulaceae). Brittonia 1990, 42, 116–120. [Google Scholar] [CrossRef]
- IUCN Red List of Threatened Species. 2024. Available online: https://www.iucnredlist.org/search?query=ipomoea&searchType=species (accessed on 20 May 2024).
- Maxted, N. Systematic and Ecogeographic Studies on Crop Genepools. 8. In An Ecogeographical Study of Vicia Subgenus Vicia; International Plant Genetic Resources Institute: Rome, Italy, 1995; Available online: https://pdf.usaid.gov/pdf_docs/PNABU773.pdf (accessed on 10 August 2022).
- Maxted, N.; Ford-Lloyd, B.V.; Hawkes, J.G. Complementary Conservation Strategies. In Plant Genetic Conservation: The In Situ Approach; Maxted, N., Ford-Lloyd, B.V., Hawkes, J.G., Eds.; Chapman and Hall: London, UK, 1997; pp. 15–41. [Google Scholar]
- Gurib-Fakim, A.J.; Gueho; Sewraj-Bissoondoyal, M. The Medicinal Plants of Mauritius—Part 1. Int. J. Pharmacogn. 1997, 35, 237–254. [Google Scholar] [CrossRef]
- Mauremootoo, J.R.; Cheke, A.S.; Watt, I. Mauritius & Rodrigues Historical Context; Unpublished Book Chapter produced for Conservation International’s State of the Hotspots publication: Madagascar & Indian Ocean Islands; 2003. Available online: https://www.researchgate.net/publication/282643787_Mauritius_Rodrigues_Historical_Context (accessed on 16 August 2023).
- Google Earth Pro. 2024. Available online: https://www.google.com/earth/about/versions/ (accessed on 24 August 2022).
- IBM Corp. IBM SPSS Statistics 21; IBM Corporation: Armonk, NY, USA, 2012; Available online: https://www.ibm.com/support/pages/spss-statistics-210-available-download (accessed on 3 July 2024).
- Mistura, C.C.; Barbieri, R.L.; Marques Castro, C.; Padulosi, S.; Alercia, A.; Bioversity International; Empresa Brasileira de Pesquisa Agropecuaria (EMBRAPA). Descriptors for Butia [Butia odorata (Barb. Rodr.) Noblick]; Bioversity International: Rome, Italy; Brazilian Agriculture Research Corporation, Embrapa Temperate Agriculture: Pelotas, RS, Brazil, 2015; p. 51. ISBN 978-92-9255-021-9. [Google Scholar]
- Alercia, A.; López, F.; Marsella, M.; Cerutti, A.L. Descriptors for Crop Wild Relatives Conserved In Situ (CWRI v.1.1); Revised Version; FAO on behalf of the International Treaty on Plant Genetic Resources for Food and Agriculture: Rome, Italy, 2022. [Google Scholar] [CrossRef]
- Fischer, G.; Hizsnyik, E.; van Velthuizen, H.; Wiberg, D.; Hermann, S. Agro-ecological Assessment. Methodology and Results. In Climate, Land, Energy & Water Strategies. A Case Study of Mauritius; IIASA: Laxenburg, Austria, 2013; Available online: https://pure.iiasa.ac.at/id/eprint/10657/1/XO-13-010.pdf (accessed on 10 July 2024).
- Dhurmea, K.R.; Boojhawon, R.; Rughooputh, S.D.D.V. A drought climatology for Mauritius using the standardized precipitation index. Hydrol. Sci. J. 2019, 64, 227–240. [Google Scholar] [CrossRef]
- Halais, P.; Davy, E.J. Notes on the 1:100,000 Agro-Climatic Map of Mauritius; Mauritius Sugar Industry Research Institute (MSIRI): Port Louis, Mauritius, 1969; Volume 23, pp. 4–15. [Google Scholar]
- BRIO-Building Resilience in the Indian Ocean. Available online: http://climat.coi-ioc.org/ (accessed on 26 March 2024).
- XLSTAT. XLSTAT Statistical Analysis Software. 2023. Available online: https://www.xlstat.com (accessed on 16 June 2024).
- Microsoft Corporation. Microsoft Excel 2007. 2024. Available online: https://support.microsoft.com/en-us/office/install-office-2007-88a8e329-3335-4f82-abb2-ecea3e319657 (accessed on 12 July 2024).
- Omni Calculator. 2024. Available online: https://www.omnicalculator.com/ (accessed on 10 July 2024).
- Wood, J.R.I.; Scotland, R.W. Notes on Ipomoea L. (Convolvulaceae) in Cuba and neighbouring islands with a checklist of species found in Cuba. Kew Bull. 2017, 72, 45. [Google Scholar] [CrossRef] [PubMed]
- Niklas, K.J. Climbing plants: Attachment and the ascent for light. Curr. Biol. 2011, 21, R199–R201. [Google Scholar] [CrossRef]
- Boodhoo, S.Y. The Changing Climate of Mauritius; Mauritius Meteorological Services: Vacoas-Phoenix, Mauritius, 2008. [Google Scholar]
- Nel, W.; Mongwa, T.; Sumner, P.D.; Anderson, R.L.; Dhurmea, K.R.; Boodhoo, Y.; Boojhawon, R.; Rughooputh, S.D.D.V. The Nature of Erosive Rainfall on a Tropical Volcanic Island with an Elevated Interior. Phys. Geogr. 2012, 33, 269–284. [Google Scholar] [CrossRef]
- Marsala, V.; Galli, A.; Paglia, G.; Miccadei, E. Landslide Susceptibility Assessment of Mauritius Island (Indian Ocean). Geosciences 2019, 9, 493. [Google Scholar] [CrossRef]
- Smith, T.M.; Shugart, H.H.; Bonan, G.B.; Smith, J.B. Modeling the potential response of vegetation to global climate change. Adv. Ecol. Res. 1992, 22, 93–116. [Google Scholar] [CrossRef]
- Yu, L.; Cao, M.; Li, K. Climate-induced changes in the vegetation pattern of China in the 21st century. Ecol. Restor. 2006, 21, 912–919. [Google Scholar] [CrossRef]
- Scheiter, S.; Higgins, S.I. Impacts of climate change on the vegetation of Africa: An adaptive dynamic vegetation modelling approach. Glob. Change Biol. 2009, 15, 2224–2246. [Google Scholar] [CrossRef]
- Kletter, A.; Von Hardenberg, J.; Meron, E.; Provenzale, A. Patterned vegetation and rainfall intermittency. J. Theor. Biol. 2009, 256, 574–583. [Google Scholar] [CrossRef] [PubMed]
- Meron, E. Pattern formation-a missing link in the study of ecosystem response to environmental changes. Math. Biosci. 2016, 271, 1–18. [Google Scholar] [CrossRef] [PubMed]
- Sun, G.; Li, L.; Li, J.; Liu, C.; Wu, Y.; Gao, S.; Wang, Z.; Feng, G. Impacts of climate change on vegetation pattern: Mathematical modeling and data analysis. Phys. Life Rev. 2022, 43, 239–270. [Google Scholar] [CrossRef] [PubMed]
- Bhardwaj, S.; Singh, S.; Ganguly, I.; Bhatia, A.K.; Bharti, V.K.; Dixit, S.P. Genome-wide diversity analysis for signatures of selection of Bos indicus adaptability under extreme agro-climatic conditions of temperate and tropical ecosystems. Anim. Gene 2021, 20, 200115. [Google Scholar] [CrossRef]
- Kumar, J.; Choudhary, A.K.; Gupta, D.S.; Kumar, S. Towards Exploitation of Adaptive Traits for Climate-Resilient Smart Pulses. Int. J. Mol. Sci. 2019, 20, 2971. [Google Scholar] [CrossRef]
- Klausmeier, C.A. Regular and irregular patterns in semiarid vegetation. Science 1999, 284, 1826–1828. [Google Scholar] [CrossRef]
- McGrath, G.S.; Paik, K.; Hinz, C. Microtopography alters self-organized vegetation patterns in water-limited ecosystems. J. Geophys. Res. Biogeosci. 2012, 117. [Google Scholar] [CrossRef]
- Sherratt, J.A. Pattern solutions of the Klausmeier model for banded vegetation in semiarid environments iv: Slowly moving patterns and their stability. SIAM J. Appl. Math. 2013, 73, 330–350. [Google Scholar] [CrossRef]
- Baartman, J.E.; Temme, A.J.; Saco, P.M. The effect of landform variation on vegetation patterning and related sediment dynamics. Earth Surf. Process. Landf. 2018, 43, 2121–2135. [Google Scholar] [CrossRef]
- Gandhi, P.; Werner, L.; Iams, S.; Gowda, K.; Silber, M. A topographic mechanism for arcing of dryland vegetation bands. J. R. Soc. Interface 2018, 15, 20180508. [Google Scholar] [CrossRef] [PubMed]
- Samkol, P.; Preston, T.R.; Ly, J. Effect of increasing offer level of water spinach (Ipomoea aquatica) on intake, growth and digestibility coefficients of rabbits. Livest. Res. Rural. Dev. 2006, 18, 81–92. Available online: http://www.lrrd.org/lrrd18/2/samk18025.htm (accessed on 25 May 2024).
- Brown, S.; Frank, M. Railroad Vine (Ipomoea pes-caprae): Identification and Uses. Edis 2020, ENH1323, 1–7. [Google Scholar] [CrossRef]
- Devall, M.S. The Biological Flora of Coastal Dunes and Wetlands. 2. Ipomoea pes-caprae (L.) Roth. J. Coast. Res. 1992, 8, 442–456. [Google Scholar]
- Akinniyi, G.; Lee, J.; Kim, H.; Lee, J.-G.; Yang, I. A Medicinal Halophyte Ipomoea pes-caprae (Linn.) R. Br.: A Review of Its Botany, Traditional Uses, Phytochemistry, and Bioactivity. Mar. Drugs 2022, 20, 329. [Google Scholar] [CrossRef]
- Guarino, L.; Maxted, N.; Chiwona, E.A. (Eds.) A Methodological Model for Ecogeographic Surveys of Crops; IPGRI Technical Bulletin: Rome, Italy, 2005; Volume 9, p. 55. ISBN 978-92-9043-690-4/92-9043-690-5. [Google Scholar]
- Shrestha, S.; Rajbhandary, S. Ipomoea indica and Ipomoea triloba (Convolvulaceae)–New Records for Flora of Nepal. J. Jpn. Bot. 2014, 89, 181–185. [Google Scholar]
- Austin, D.F. Nomenclature of the Ipomoea nil complex (Convolvulaceae). Taxon 1986, 35, 355–358. [Google Scholar] [CrossRef]
- The National Invasive Alien Species Strategy for the Republic of Mauritius 2008–2017. Available online: https://npcs.govmu.org/Documents/Reports/National%20Invasive%20Alien%20Species%20Strategy%20for%20the%20Republic%20of%20Mauritius.pdf (accessed on 27 June 2024).
- Kaudeer, S.; Pandoo, S. Controlling Invasive Alien Species (IAS) to Protect Biodiversity, Economy and Health in Mauritius. 2021. Available online: https://www.undp.org/mauritius-seychelles/blog/controlling-invasive-alien-species-ias-protect-biodiversity-economy-and-health-mauritius (accessed on 27 June 2024).
- CABI Compendium. CABI International, Ipomoea alba (White Moonflower). 2022. Available online: https://www.cabidigitallibrary.org/doi/10.1079/cabicompendium.119823 (accessed on 16 February 2024).
- CABI Compendium. CABI International, Ipomoea indica (Ocean Blue Morning-Glory). 2022. Available online: https://www.cabidigitallibrary.org/doi/10.1079/cabicompendium.107833 (accessed on 16 February 2024).
- PIER (Pacific Island Ecosystems at Risk), US Forest Service. In Pacific Islands Ecosystems at Risk; HEAR, University of Hawaii: Honolulu, HI, USA, 2017; Available online: http://www.hear.org/pier/ (accessed on 3 May 2024).
- Weeds of New Zealand, Weed Busters. 2017. Available online: https://www.marlborough.govt.nz/repository/libraries/id:1w1mps0ir17q9sgxanf9/hierarchy/Documents/Environment/Biosecurity/BlueMorningGlory.pdf (accessed on 3 May 2024).
- Queensland Government. Weeds of Australia, Biosecurity Queensland Edition. In Weeds of Australia, Biosecurity Queensland Edition; Queensland Government: Australia, 2017; Available online: http://keyserver.lucidcentral.org/weeds/data/media/Html/search.html (accessed on 3 May 2024).
- Neto, A.; Cunha, S.; Voigt, E. Vegetative Propagation of the Dune Building Plant for Use in Dune Rehabilitation Projects Ipomoea Pes-Caprae. J. Coast. Res. 2006, 39, 1251–1254. [Google Scholar]
- Sucre, B.; Suárez, N. Effect of Salinity and PEG-Induced Water Stress and Water Status, Gas Exchange, Solute Accumulation and Leaf Growth in Ipomoea Pes-Caprae. Environ. Exp. Bot. 2011, 70, 192–203. [Google Scholar] [CrossRef]
- Maxted, N.; Magos Brehm, J.; Kell, S. Resource Book for Preparation of National Conservation Plans for Crop Wild Relatives and Landraces; FAO: Rome, Italy, 2013; Volume 463. [Google Scholar]
- IUCN. Environmental Synopsis, Mauritius. 1993. Available online: https://portals.iucn.org/library/sites/library/files/documents/Co-MU-Env-005.pdf (accessed on 19 August 2023).
- Mauremootoo, J.; Pandoo, S.; Bachraz, V.; Buldawoo, I.; Cole, N. Invasive species management in Mauritius: From the reactive to the proactive—The National Invasive Species Management Strategy and its implementation. In Island Invasives: Scaling up to Meet the Challenge; Veitch, C.R., Clout, M.N., Martin, A.R., Russell, J.C., West, C.J., Eds.; IUCN Occasional Paper SSC: Gland, Switzerland, 2019; Volume 62, pp. 503–509. [Google Scholar]
- Convention on Biological Diversity (CBD). Mauritius-Country Profile. 2024. Available online: https://www.cbd.int/countries/profile?country=mu (accessed on 23 March 2024).
- Heywood, V.H.; Brummitt, R.K.; Culham, A.; Seberg, O. Flowering Plant Families of the World; Firefly Books: Richmond Hill, ON, Canada; Kutztown, PA, USA, 2007; 424p. [Google Scholar]
- Lionakis Meyer, D.J.; Price, R.A. Important Seed Characters for Certain Species in the Morning-glory Family (Convolvulaceae). 2020. Available online: https://analyzeseeds.com/wp-content/uploads/2020/11/Convolvulaceae-2020-complete.pdf (accessed on 10 March 2024).
- Molinar, R.H. Indigenous Asian speciality vegetables in the Central Valley of California. HortScience 2012, 47, 835–838. [Google Scholar] [CrossRef]
- Ministry of Environment and Sustainable Development. Fourth National Report on the Convention on Biological Diversity, Republic of Mauritius. 2010. Available online: https://www.cbd.int/doc/world/mu/mu-nr-04-en.pdf (accessed on 20 July 2023).
- United Nations Food Systems Summit (UNFSS 2021) Action Track 1: Ensure Access to Safe and Nutritious Food for All. Available online: https://www.un.org/en/food-systems-summit/action-tracks (accessed on 20 March 2024).
- Ministry of Agro Industry and Food Security. The Forestry Service. 2024. Available online: https://forestry.govmu.org/Pages/Forests-in-Mauritius.aspx#:~:text=%E2%80%8B%E2%80%8B%E2%80%8B%20Forests%20in%20Mauritius%E2%80%8B&text=Approximately%20147%2023%20ha%20of,with%20good%20quality%20native%20forests (accessed on 11 August 2024).
- NSAP. The National Strategic Action Plan for the Conservation and Sustainable Use of Crop Wild Relatives in The Republic of Mauritius; University of Mauritius and Ministry of AgroIndustry and Food Security: Port Louis, Mauritius, 2016. [Google Scholar]
- Pressey, R.L.; Cowling, R.M.; Rouget, M. Formulating conservation targets for biodiversity pattern and process in the Cape Floristic Region, South Africa. Biol. Conserv. 2003, 112, 99–127. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | No of Site | Percentage Occurrence |
|---|---|---|
| I. indica | 137 | 29.0 |
| I. alba | 123 | 26.0 |
| I. obscura | 107 | 22.6 |
| I. cairica | 44 | 9.3 |
| I. pes-caprae | 14 | 3.0 |
| I. tilliacea | 12 | 2.5 |
| I. ochracea | 7 | 1.5 |
| I. batatas | 6 | 1.3 |
| I. carnea | 5 | 1.1 |
| I. nil | 5 | 1.1 |
| I. hederifolia | 4 | 0.6 |
| I. triloba | 3 | 0.6 |
| I. quamoclit | 2 | 0.4 |
| I. littoralis | 2 | 0.4 |
| I. aquatica | 2 | 0.4 |
| I. purpurea | 1 | 0.2 |
| Summary Statistics: | ||||
|---|---|---|---|---|
| Variable | Richness | Total Number of Individuals | Shannon Diversity Index | Pielou Evenness Index |
| Moka | 9 | 91 | 1.85 | 0.84 |
| Pamplemousses | 9 | 63 | 1.52 | 0.69 |
| Black River | 8 | 30 | 1.73 | 0.83 |
| Grand Port | 8 | 75 | 1.33 | 0.64 |
| Plaines-Wilhems | 7 | 93 | 1.32 | 0.68 |
| Savanne | 7 | 58 | 1.62 | 0.83 |
| Flacq | 7 | 28 | 1.51 | 0.77 |
| Rivière du Rempart | 7 | 26 | 1.32 | 0.68 |
| Port-Louis | 1 | 3 | 0 | - |
| Areas Delimitated within the Districts of Mauritius | |||
|---|---|---|---|
| Pamplemousses | Riviere du Rempart | Flacq | Grand Port |
| Terre Rouge | Grand Baie | St Julien d’Hotman | Grand sable |
| Tombeau Bay | Vale | Bon Accueil | BambousVirieux |
| Balaclava | Pereybere | Lalmatie | Bois des Amourettes |
| Pointe aux Piments | Petit Raffray | Queen Victoria | Vieux Grand Port |
| Pointe aux Biches | Goodlands | Central Flacq | Ferney |
| Troux aux Biches | Roche noire | Quatre Cocos | Riviere des Creoles |
| Pamplemousses | Mare d’australia | Belle mare | Mahebourg |
| Triolet | Rivière du Rempart | Palmar | Plaine Magnien |
| Ville Bague | Cottage | Trou d’Eau Douce | Trois Boutiques |
| Mont Goût | Cap Malheureux | Beau champ | Le bouchon |
| Poudre d’Or | Grand River South East | Mon Desert | |
| Calodyne | Quatre Soeurs | L’escalier | |
| Sebastopol | Mare D’albert | ||
| Belle Rive | Rose Belle | ||
| Belle Air | Banane/clunny | ||
| Savanne | Black River | Plaine Wilhems | Moka |
| Grand Bois | Chamarel | Le Pétrin | Réduit |
| Bois Cheri | Le Morne | Midlands | Verdun |
| Grand Bassin | La Gaulette | La vigie | Quartier-Militaire |
| Black River Gorges | Case Noyale | Eau Coulée | Belle Rive |
| Bassin blanc | Grande Riviere Noire | Camp Fouquereaux | Providence |
| Chamouny | La Preneuse | Curepipe | Melrose |
| Chemin Grenier | Tamarin | Trou aux Cerfs | Montagne Blanche |
| St Felix | Bamboo | Valentina | Alma |
| Riambel | Gros cailloux | Vacoas-Phoenix | |
| Pomponette beach | Albion | Mon Désir | |
| Souillac | Carreau laliane | Port Louis | |
| Gris gris | Solferino | Long Mountain | |
| St Aubin | Quatre Bornes | Caroline | |
| Rivière des Anguilles | Ebène | Roche Bois | |
| Britannia | |||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Boyjnath, Y.; Dulloo, M.E.; Bhoyroo, V.; Ranghoo-Sanmukhiya, V.M. Ecogeographic Study of Ipomoea Species in Mauritius, Indian Ocean. Plants 2024, 13, 2706. https://doi.org/10.3390/plants13192706
Boyjnath Y, Dulloo ME, Bhoyroo V, Ranghoo-Sanmukhiya VM. Ecogeographic Study of Ipomoea Species in Mauritius, Indian Ocean. Plants. 2024; 13(19):2706. https://doi.org/10.3390/plants13192706
Chicago/Turabian StyleBoyjnath, Yakshini, Mohammad Ehsan Dulloo, Vishwakalyan Bhoyroo, and Vijayanti Mala Ranghoo-Sanmukhiya. 2024. "Ecogeographic Study of Ipomoea Species in Mauritius, Indian Ocean" Plants 13, no. 19: 2706. https://doi.org/10.3390/plants13192706