Amino Acid Transporters in Plants: Identification and Function

Abstract
:1. Introduction
2. Amino Acid Uptake, Transport, and Distribution
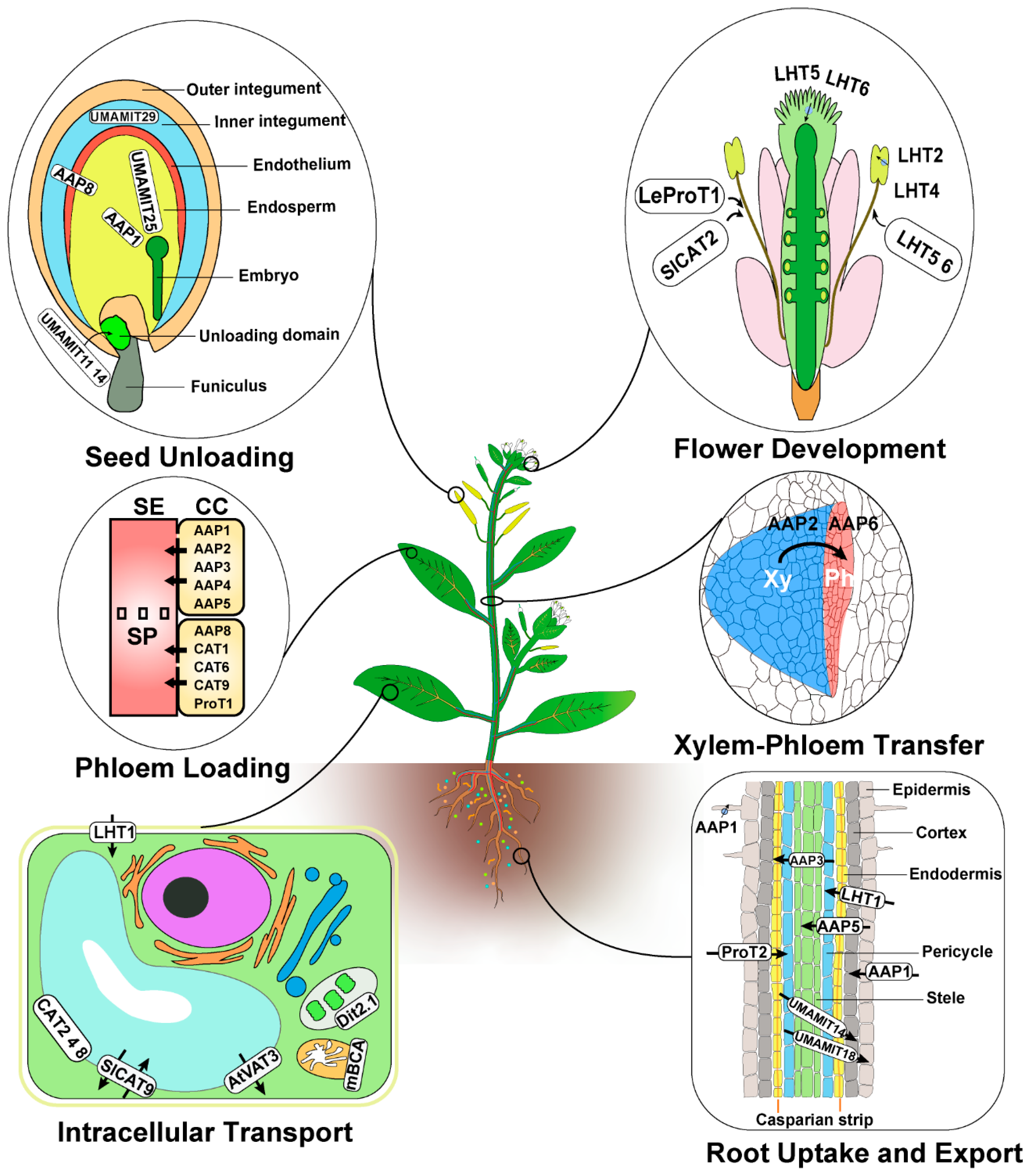
2.1. Transporters Mediating Root Amino Acid Uptake
2.2. Transporters Function in Xylem–Phloem Transfer and Intercellular Transport of Amino Acids
2.3. Transporter Function in Phloem Loading of Amino Acids
2.4. Transporter Function in Phloem Unloading of Amino Acids in Sink Organs
3. The Role of Exporters in Amino Acid Translocation
4. Is the Functional Overlap of Multiple Amino Acid Transporters Redundant or Necessary?
5. Regulation of Amino Acid Transporters in Response to Environmental Stimuli
6. Transport Function in Carbon Metabolism
7. Future Perspectives
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Nsholm, T.; Kielland, K.; Ganeteg, U. Uptake of organic nitrogen by plants. New Phytol. 2009, 182, 31–48. [Google Scholar] [CrossRef] [PubMed]
- Rentsch, D. Uptake and partitioning of amino acids and peptides. Mol. Plant 2010, 3, 997–1011. [Google Scholar]
- Bloom, A.J. The increasing importance of distinguishing among plant nitrogen sources. Curr. Opin. Plant Biol. 2015, 25, 10–16. [Google Scholar] [CrossRef] [PubMed]
- Paungfoo-Lonhienne, C.; Lonhienne, T.G.A.; Rentsch, D.; Robinson, N.; Christie, M.; Webb, R.I.; Gamage, H.K.; Carroll, B.J.; Schenk, P.M.; Schmidt, S. Plants can use protein as a nitrogen source without assistance from other organisms. Proc. Natl. Acad. Sci. USA 2008, 105, 4524–4529. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tegeder, M.; Masclaux-Daubresse, C. Source and sink mechanisms of nitrogen transport and use. New Phytol. 2018, 217, 35–53. [Google Scholar] [CrossRef] [Green Version]
- Rentsch, D.; Schmidt, S.; Tegeder, M. Transporters for uptake and allocation of organic nitrogen compounds in plants. FEBS Lett. 2007, 581, 2281–2289. [Google Scholar] [CrossRef] [Green Version]
- Pratelli, R.; Pilot, G. Regulation of amino acid metabolic enzymes and transporters in plants. J. Exp. Bot. 2014, 65, 5535–5556. [Google Scholar] [CrossRef]
- Besnard, J.; Pratelli, R.; Zhao, C.S.; Sonawala, U.; Collakova, E.; Pilot, G.; Okumoto, S. UMAMIT14 is an amino acid exporter involved in phloem unloading in Arabidopsis roots. J. Exp. Bot. 2016, 67, 6385–6397. [Google Scholar] [CrossRef] [Green Version]
- Lohaus, G.; Burba, M.; Heldt, H.W. Comparison of the contents of sucrose and amino acids in the leaves, phloem sap and taproots of high and low sugar-producing hybrids of sugar beet (Beta vulgaris L.). J. Exp. Bot. 1994, 45, 1097–1101. [Google Scholar] [CrossRef]
- Lam, H.M. Use of Arabidopsis mutants and genes to study amide amino acid biosynthesis. Plant Cell 1995, 7, 887–898. [Google Scholar]
- Masclaux-Daubresse, C.; Daniel-Vedele, F.; Julie, D.; Fabien, C.; Laure, G.; Akira, S. Nitrogen uptake, assimilation and remobilization in plants: Challenges for sustainable and productive agriculture. Ann. Bot. 2010, 105, 1141–1157. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dinkeloo, K.; Boyd, S.; Pilot, G. Update on amino acid transporter functions and on possible amino acid sensing mechanisms in plants. Semin. Cell Dev. Biol. 2018, 74, 105–113. [Google Scholar] [CrossRef] [PubMed]
- Tegeder, M. Transporters for amino acids in plant cells: Some functions and many unknowns. Curr. Opin. Plant Biol. 2012, 15, 315–321. [Google Scholar] [CrossRef] [PubMed]
- Ortiz-Lopez, A.; Chang, H.C.; Bush, D.R. Amino acid transporters in plants. Biochim. Biophys. Acta 2000, 1465, 275–280. [Google Scholar] [CrossRef] [Green Version]
- Wan, Y.F.; King, R.; Mitchell, R.A.C.; Hassani-Pak, K.; Hawkesford, M.J. Spatiotemporal expression patterns of wheat amino acid transporters reveal their putative roles in nitrogen transport and responses to abiotic stress. Sci. Rep. 2017, 7, 5461. [Google Scholar] [CrossRef] [PubMed]
- Okumoto, S.; Pilot, G. Amino acid export in plants: A missing link in nitrogen cycling. Mol. Plant 2011, 4, 453–463. [Google Scholar] [CrossRef] [Green Version]
- Ladwig, F.; Stahl, M.; Ludewig, U.; Hirner, A.A.; Hammes, U.Z.; Stadler, R.; Harter, K.; Koch, W. Siliques are Red1 from Arabidopsis acts as a bidirectional amino acid transporter that is crucial for the amino acid homeostasis of siliques. Plant Physiol. 2012, 158, 1643–1655. [Google Scholar] [CrossRef] [Green Version]
- Ranocha, P.; Dima, O.; Nagy, R.; Felten, J.; Corratgé-Faillie, C.; Novák, O.; Morreel, K.; Lacombe, B.; Martinez, Y.; Pfrunder, S.; et al. Arabidopsis WAT1 is a vacuolar auxin transport facilitator required for auxin homoeostasis. Nat. Commun. 2013, 4, 2625. [Google Scholar] [CrossRef]
- Liu, X.; Bush, D.R. Expression and transcriptional regulation of amino acid transporters in plants. Amino Acids 2006, 30, 113–120. [Google Scholar] [CrossRef]
- Schwacke, R.; Grallath, S.; Breitkreuz, K.E.; Stransky, E.; Stransky, H.; Frommer, W.B.; Rentsch, D. LeProT1, a transporter for proline, glycine betaine, and γ-amino butyric acid in tomato pollen. Plant Cell 1999, 11, 377–391. [Google Scholar]
- Koch, W.; Kwart, M.; Laubner, M.; Heineke, D.; Stransky, H.; Frommer, W.B.; Tegeder, M. Reduced amino acid content in transgenic potato tubers due to antisense inhibition of the leaf H+/amino acid symporter StAAP1. Plant J. 2003, 33, 211–220. [Google Scholar] [CrossRef] [PubMed]
- Miranda, M.; Borisjuk, L.; Tewes, A.; Heim, U.; Sauer, N.; Wobus, U.; Weber, H. Amino acid permeases in developing seeds of Vicia faba L.: Expression precedes storage protein synthesis and is regulated by amino acid supply. Plant J. 2001, 28, 61–71. [Google Scholar] [CrossRef] [PubMed]
- Kohl, S.; Hollmann, J.; Blattner, F.R.; Radchuk, V.; Andersch, F.; Steuernagel, B.; Schmutzer, T.; Scholz, U.; Krupinska, K.; Weber, H.; et al. A putative role for amino acid permeases in sink-source communication of barley tissues uncovered by RNA-seq. BMC Plant Biol. 2012, 12, 154. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pan, X.Y.; Hasan, M.M.; Li, Y.Q.; Liao, C.S.; Zheng, H.Y.; Liu, R.Y.; Li, X.X. Asymmetric transcriptomic signatures between the cob and florets in the maize ear under optimal- and low-nitrogen conditions at silking, and functional characterization of amino acid transporters ZmAAP4 and ZmVAAT3. J. Exp. Bot. 2015. [Google Scholar] [CrossRef] [PubMed]
- Tegeder, M.; Offler, C.E.; Frommer, W.B.; Patrick, J.W. Amino acid transporters are localized to transfer cells of developing pea seeds. Plant Physiol. 2000, 122, 319–325. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, H.M.; Ma, H.L.; Yu, L.; Wang, X.; Zhao, J. Genome-wide survey and expression analysis of amino acid transporter gene family in rice (Oryza sativa L.). PLoS ONE 2012, 7, e49210. [Google Scholar] [CrossRef] [PubMed]
- Tan, Q.; Grennan, A.K.; Pélissier, H.C.; Rentsch, D.; Tegeder, M. Characterization and expression of French bean amino acid transporter PvAAP1. Plant Sci. 2008, 174, 348–356. [Google Scholar] [CrossRef]
- Frommer, W.B.; Hummel, S.; Riesmeier, J.W. Expression cloning in yeast of a cDNA encoding a broad specificity amino acid permease from Arabidopsis thaliana. Proc. Natl. Acad. Sci. USA 1993, 90, 5944–5948. [Google Scholar] [CrossRef] [Green Version]
- Hsu, L.C.; Chiou, T.J.; Chen, L.; Bush, D.R. Cloning a plant amino acid transporter by functional complementation of a yeast amino acid transport mutant. Proc. Natl. Acad. Sci. USA 1993, 90, 7441–7445. [Google Scholar] [CrossRef] [Green Version]
- Cheng, L.; Yuan, H.Y.; Ren, R.; Zhao, S.Q.; Han, Y.P.; Zhou, Q.Y.; Ke, D.X.; Wang, Y.X.; Wang, L. Genome-wide identification, classification, and expression analysis of amino acid transporter gene family in glycine max. Front. Plant Sci. 2016, 7, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Ma, H.L.; Cao, X.L.; Shi, S.D.; Li, S.L.; Gao, J.P.; Ma, Y.L.; Zhao, Q.; Chen, Q. Genome-wide survey and expression analysis of the amino acid transporter superfamily in potato (Solanum tuberosum L.). Plant Physiol. Biochem. 2016, 107, 164–177. [Google Scholar] [CrossRef] [PubMed]
- Wu, M.; Wu, S.N.; Chen, Z.; Dong, Q.; Yan, H.W.; Xiang, Y. Genome-wide survey and expression analysis of the amino acid transporter gene family in poplar. Tree Genet. Genomes 2015, 11, 1–20. [Google Scholar] [CrossRef]
- Wipf, D.; Loqué, D.; Lalonde, S.; Frommer, W.B. Amino acid transporter inventory of the selaginella genome. Front. Plant Sci. 2012, 3. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xie, Y.; Zhao, J.L.; Wang, C.W.; Yu, A.X.; Liu, N.; Chen, L.; Lin, F.; Xu, H.H. Glycinergic–fipronil uptake is mediated by an amino acid carrier system and induces the expression of amino acid transporter genes in ricinus communis seedlings. J. Agric. Food Chem. 2016, 64, 3810–3818. [Google Scholar] [CrossRef]
- Weber, A.P.M. Intracellular metabolite transporters in plants. Mol. Plant 2010, 3, 21–53. [Google Scholar]
- Hoyos, M.E.; Palmieri, L.; Wertin, T.; Arrigoni, R.; Polacco, J.C.; Palmieri, F. Identification of a mitochondrial transporter for basic amino acids in Arabidopsis thaliana by functional reconstitution into liposomes and complementation in yeast. Plant J. 2003, 33, 1027–1035. [Google Scholar] [CrossRef]
- Petra, R.; Dressen, U.; Hebbeker, U.; Hille, D.; Weber, A.P.M. The Arabidopsis mutant dct is deficient in the plastidic glutamate/malate translocator DiT2. Plant J. 2003, 35, 316–331. [Google Scholar]
- Palmieri, L.; Todd, C.D.; Arrigoni, R.; Hoyos, M.E.; Santoro, A.; Polacco, J.C.; Palmieri, F. Arabidopsis mitochondria have two basic amino acid transporters with partially overlapping specificities and differential expression in seedling development. Biochim. Biophys. Acta Bioenerg. 2006, 1757, 1277–1283. [Google Scholar] [CrossRef] [Green Version]
- Dündar, E.; Bush, D.R. BAT1, a bidirectional amino acid transporter in Arabidopsis. Planta 2009, 229, 1047–1056. [Google Scholar] [CrossRef]
- Sekito, T.; Fujiki, Y.; Ohsumi, Y.; Kakinuma, Y. Novel families of vacuolar amino acid transporters. IUBMB Life 2008, 60, 519–525. [Google Scholar] [CrossRef]
- Pudelski, B.; Kraus, S.; Soll, J.; Philippar, K. The plant prat proteins-preprotein and amino acid transport in mitochondria and chloroplasts. Plant Biol. 2010, 12, 42–55. [Google Scholar] [CrossRef] [PubMed]
- Wipf, D.; Ludewig, U.; Tegeder, M.; Rentsch, D.; Koch, W.; Frommer, W.B. Conservation of amino acid transporters in fungi, plants and animals. Trends Biochem. Sci. 2002, 27, 139–147. [Google Scholar] [CrossRef]
- Tegeder, M.; Hammes, U.Z. The way out and in: Phloem loading and unloading of amino acids. Curr. Opin. Plant Biol 2018, 43, 16–21. [Google Scholar] [CrossRef] [PubMed]
- Tegeder, M. Transsporters involved in source to sink partitioning of amino acids and ureides: Opportunities for crop improvement. J. Exp. Bot. 2014, 65, 1865–1878. [Google Scholar] [CrossRef] [PubMed]
- Fischer, W.N.; Loo, D.D.F.; Koch, W.; Ludewig, U.; Frommer, W.B. Low and high affinity amino acid H+-cotransporters for cellular import of neutral and charged amino acids. Plant J. 2002, 29, 717–731. [Google Scholar] [CrossRef]
- Okumoto, S. High affinity amino acid transporters specifically expressed in xylem parenchyma and developing seeds of arabidopsis. J. Biol. Chem. 2002, 277, 45338–45346. [Google Scholar] [CrossRef] [Green Version]
- Grallath, S.; Weimar, T.; Meyer, A.; Gumy, C.; Suter-Grotemeyer, M.; Neuhaus, J.M.; Rentsch, D. The AtProT family: Compatible solute transporters with similar substrate specificity but differential expression patterns. Plant Physiol. 2005, 137, 117–126. [Google Scholar] [CrossRef] [Green Version]
- Lee, Y.H.; Foster, J.; Chen, J.; Voll, L.M.; Weber, A.P.M.; Tegeder, M. AAP1 transports uncharged amino acids into roots of arabidopsis. Plant J. 2007, 50, 305–319. [Google Scholar] [CrossRef]
- Molly, P.; Justin, F.; Mechthild, M. Different and overlapping functions of Arabidopsis LHT6 and AAP1 transporters in root amino acid uptake. J. Exp. Bot. 2014, 65, 5193–5204. [Google Scholar]
- Svennerstam, H.; Ganeteg, U.; Näsholm, T. Root uptake of cationic amino acids by Arabidopsis depends on functional expression of amino acid permease 5. New Phytol. 2008, 180, 620–630. [Google Scholar] [CrossRef]
- Svennerstam, H.; Jmtgrd, S.; Ahmad, I.; Huss-Danell, K.; Ganeteg, N.U. Transporters in Arabidopsis roots mediating uptake of amino acids at naturally occurring concentrations. New Phytol. 2011, 191, 459–467. [Google Scholar] [CrossRef] [PubMed]
- Birnbaum, K.; Shasha, D.E.; Wang, J.Y.; Jung, J.W.; Lambert, G.M.; Galbraith, D.W.; Benfey, P.N. A gene expression map of the Arabidopsis root. Science 2003, 302, 1956–1960. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fischer, W.N.; Andre, B.; Rentsch, D.; Krolkiewicz, S.; Tegeder, M.; Breitkreuz, K.; Frommer, W.B. Amino acid transport in plants. Trends Plant Sci. 1998, 3, 188–195. [Google Scholar] [CrossRef]
- Bush, C.D.R. LHT1, a lysine- and histidine-specific amino acid transporter in Arabidopsis. Plant Physiol. 1997, 115, 1127–1134. [Google Scholar]
- Himer, A.; Ladwig, F.; Stransky, H.; Okumoto, S.; Keinath, M.; Harms, A.; Frommer, W.B.; Koch, W. Arabidopsis LHT1 is a high-affinity transporter for cellular amino acid uptake in both root epidermis and leaf mesophyll. Plant Cell 2006, 18, 1931–1946. [Google Scholar]
- Svennerstam, H.; Ganeteg, U.; Bellini, C.; Näsholm, T. Comprehensive screening of Arabidopsis mutants suggests the lysine histidine transporter 1 to be involved in plant uptake of amino acids. Plant Physiol. 2007, 143, 1853–1860. [Google Scholar] [CrossRef] [Green Version]
- Ganeteg, U.; Ahmad, I.; Jmtgrd, S.; Aguetoni-Cambui, C.; Nsholm, T. Amino acid transporter mutants of Arabidopsis provides evidence that a non-mycorrhizal plant acquires organic nitrogen from agricultural soil. Plant Cell Environ. 2016, 40, 413–423. [Google Scholar] [CrossRef]
- Brady, S.M.; Orlando, D.A.; Lee, J.Y.; Wang, J.Y.; Koch, J.; Dinneny, J.R.; Mace, D.; Ohler, U.; Benfey, P.N. A high-resolution root spatiotemporal map reveals dominant expression patterns. Science 2007, 318, 801–806. [Google Scholar] [CrossRef]
- Guo, N.; Hu, J.Q.; Yan, M.; Qu, H.Y.; Luo, L.; Tegeder, M.; Xu, G.H. Oryza sativa Lysine-Histidine-type Transporter 1 functions in root uptake and root-to-shoot allocation of amino acids in rice. Plant J. 2020. [Google Scholar] [CrossRef]
- Lu, K.; Wu, B.; Wang, J.; Zhu, W.; Nie, H.; Qian, J.; Huang, W.; Fang, Z. Blocking amino acid transporter OsAAP3 improves grain yield by promoting outgrowth buds and increasing tiller number in rice. Plant Biotechnol. J. 2018, 16, 1710–1722. [Google Scholar] [CrossRef] [Green Version]
- Peng, B.; Kong, H.L.; Li, Y.B.; Wang, L.Q.; Zhong, M.; Sun, L.; Gao, G.J.; Zhang, Q.L.; Luo, L.J.; Wang, G.W.; et al. OsAAP6 functions as an important regulator of grain protein content and nutritional quality in rice. Nat. Commun. 2014. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Breitkreuz, K.E.; Shelp, B.J.; Fischer, W.N.; Schwacke, R.; Rentsch, D. Identification and characterization of GABA, proline and quaternary ammonium compound transporters from Arabidopsis thaliana. FEBS Lett. 1999, 450, 280–284. [Google Scholar] [CrossRef] [Green Version]
- Rentsch, D.; Hirner, B.; Schmelzer, E.; Frommer, W.B. Salt stress-induced proline transporters and salt stress-repressed broad specificity amino acid permeases identified by suppression of a yeast amino acid permease-targeting mutant. Plant Cell 1996, 8, 1437–1446. [Google Scholar] [PubMed] [Green Version]
- Fujiwara, T.; Mitsuya, S.; Miyake, H.; Hattori, T.; Takabe, T. Characterization of a novel glycinebetaine/proline transporter gene expressed in the mestome sheath and lateral root cap cells in barley. Planta 2010, 232, 133–143. [Google Scholar] [CrossRef]
- Lehmann, S.; Gumy, C.; Blatter, E.; Boeffel, S.; Fricke, W.; Rentsch, D. In planta function of compatible solute transporters of the AtProT family. J. Exp. Bot. 2011, 62, 787–796. [Google Scholar] [CrossRef] [PubMed]
- Forsum, O.; Svennerstam, H.; Ganeteg, U.; Näsholm, T. Capacities and constraints of amino acid utilization in Arabidopsis. New Phytol. 2008, 179, 1058–1069. [Google Scholar] [PubMed]
- Pate, J.S.; Sharkey, P.J.; Lewis, O.A.M. Xylem to phloem transfer of solutes in fruiting shoots of legumes, studies by a phloem-bleeding technique. Planta 1975, 122, 11–26. [Google Scholar] [CrossRef]
- Atkins, C. Biochemical aspects of assimilate transfers along the phloem path: N-solutes in lupins. Aust. J. Plant Physiol. 2000, 27, 531–537. [Google Scholar] [CrossRef]
- Offler, C.E.; McCurdy, D.W.; Patrick, J.W.; Talbot, M.J. Transfer cells: Cells specialized for a special purpose. Annu. Rev. Plant Biol. 2003, 54, 431–454. [Google Scholar] [CrossRef]
- Hunt, E.; Gattolin, S.; Newbury, H.J.; Bale, J.S.; Tseng, H.M.; Barrett, D.A.; Pritchard, J. A mutation in amino acid permease AAP6 reduces the amino acid content of the Arabidopsis sieve elements but leaves aphid herbivores unaffected. J. Exp. Bot. 2010, 61, 55–64. [Google Scholar] [CrossRef]
- Zhang, L.Z.; Tan, Q.M.; Lee, R.; Trethewy, A.; Lee, Y.H.; Tegeder, M. Altered xylem-phloem transfer of amino acids affects metabolism and leads to increased seed yield and oil content in Arabidopsis. Plant Cell 2010, 22, 3603–3620. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, H.Y.; Krebs, M.; Stierhof, Y.D.; Ludewig, U. Characterization of the putative amino acid transporter genes AtCAT2, 3 & 4: The tonoplast localized AtCAT2 regulates soluble leaf amino acids. J. Plant Physiol. 2014, 171, 594–601. [Google Scholar] [PubMed]
- Su, Y.H.; Frommer, W.B.; Ludewig, U. Molecular and functional characterization of a family of amino acid transporters from Arabidopsis. Plant Physiol. 2004, 136, 3104–3113. [Google Scholar] [CrossRef] [Green Version]
- Yang, H.; Bogner, M.; Stierhof, Y.D.; Ludewig, U. H+-Independent glutamine transport in plant root tips. PLoS ONE 2010, 5, e8917. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Snowden, C.J.; Thomas, B.; Baxter, C.J.; Smith, J.A.C.; Sweetlove, L.J. A tonoplast Glu/Asp/GABA exchanger that affects tomato fruit amino acid composition. Plant J. 2015, 81, 651–660. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, Y.W.; Yang, L.T.; Li, Z.G. Molecular cloning and identification of a putative tomato cationic amino acid transporter-2 gene that is highly expressed in stamens. Plant Cell Tissue Organ Cult. 2012, 112, 55–63. [Google Scholar] [CrossRef]
- Russnak, R.; Konczal, D.; Mcintire, S.L. A family of yeast proteins mediating bidirectional vacuolar amino acid transport. J. Biol. Chem. 2001, 276, 23849–23857. [Google Scholar] [CrossRef] [Green Version]
- Fujiki, Y.; Teshima, H.; Kashiwao, S.; Kawano-Kawada, M.; Ohsumi, Y.; Kakinuma, Y.; Sekito, T. Functional identification of AtAVT3, a family of vacuolar amino acid transporters, in Arabidopsis. FEBS Lett. 2016, 591, 5–15. [Google Scholar] [CrossRef] [Green Version]
- Turgeon, R.; Wolf, S. Phloem transport: Cellular pathways and molecular trafficking. Annu. Rev. Plant Biol. 2009, 60, 207–221. [Google Scholar] [CrossRef] [Green Version]
- Lalonde, S.; Tegeder, M.; Throne-Holst, M.; Frommer, W.B.; Patrick, J.W. Phloem loading and unloading of sugars and amino acids. Plant Cell Environ. 2003, 26, 37–56. [Google Scholar] [CrossRef] [Green Version]
- Tegeder, M.; Ward, J.M. Molecular evolution of plant AAP and LHT amino acid transporters. Front. Plant Sci. 2012, 3. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fischer, W.N.; Kwart, M.; Hummel, S.; Frommer, W.B. Substrate specificity and expression profile of amino acid transporters (AAPs) in Arabidopsis. J. Biol. Chem. 1995, 270, 16315–16320. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Okumoto, S.; Koch, W.; Tegeder, M.; Fischer, W.N.; Biehl, A.; Leister, D.; Stierhof, Y.D.; Frommer, W.B. Root phloem-specific expression of the plasma membrane amino acid proton co-transporter AAP3. J. Exp. Bot. 2004, 55, 2155–2168. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Santiago, J.P.; Tegeder, M. Connecting source with sink: The role of Arabidopsis AAP8 in phloem loading of amino acids. Plant Physiol. 2016, 171, 508–521. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Santiago, J.P.; Tegeder, M. Implications of nitrogen phloem loading for carbon metabolism and transport during Arabidopsis development. J. Integr. Plant Biol. 2017, 59, 409–421. [Google Scholar] [CrossRef] [Green Version]
- Zhang, L.Z.; Garneau, M.G.; Majumdar, R.; Grant, J.; Tegeder, M. Improvement of pea biomass and seed productivity by simultaneous increase of phloem and embryo loading with amino acids. Plant J. 2014, 81, 134–146. [Google Scholar] [CrossRef]
- Hammes, U.Z.; Nielsen, E.; Honaas, L.A.; Taylor, C.G.; Schachtman, D.P. AtCAT6, a sink-tissue-localized transporter for essential amino acids in Arabidopsis. Plant J. 2006, 48, 414–426. [Google Scholar] [CrossRef]
- Hunt, E.J.; Pritchard, J.; Bennett, M.J.; Zhu, X.; Barrett, D.A.; Allen, T.; Bale, J.S.; Newbury, H.J. The Arabidopsis thaliana/Myzus persicae model system demonstrates that a single gene can influence the interaction between a plant and a sap-feeding insect. Mol. Ecol. 2006, 15, 4203–4213. [Google Scholar] [CrossRef]
- Patrick, J.W. Fundamentals of phloem transport physiology. In Phloem: Molecular Cell Biology, Systemic Communication, Biotic Interactions; Thompson, G.A., van Bel, A.J.E., Eds.; Wiley-Blackwell: Hoboken, NJ, USA, 2012; pp. 30–59. [Google Scholar]
- RossElliott, T.J.; Jensen, K.H.; Haaning, K.S.; Wager, B.M.; Knoblauch, J.; Howell, A.H.; Mullendore, D.L.; Monteith, A.G.; Paultre, D.; Yan, D.W.; et al. Phloem unloading in Arabidopsis roots is convective and regulated by the phloem-pole pericycle. Elife 2017, 6, e24125. [Google Scholar] [CrossRef] [Green Version]
- Stadler, R.; Sauer, L.N. Cell-To-cell movement of green fluorescent protein reveals post-phloem transport in the outer integument and identifies symplastic domains in Arabidopsis seeds and embryos. Plant Physiol. 2005, 139, 701–712. [Google Scholar] [CrossRef] [Green Version]
- Tegeder, M.; Ruan, Y.L.; Patrick, J.W. Roles of plasma membrane transporters in phloem functions. Mol. Cell Biol. Syst. Commun. Biot. Interact. 2012, 63–101. [Google Scholar] [CrossRef]
- Patrick, J.W. Sieve element unloading: Cellular pathway, mechanism and control. Physiol. Plant. 1990, 78, 298–308. [Google Scholar] [CrossRef]
- Frommer, W.B.; Hummel, S.; Unseld, M.; Ninnemann, O. Seed and vascular expression of a high-affinity transporter for cationic amino acids in Arabidopsis. Proc. Natl. Acad. Sci. USA 1995, 92, 12036–12040. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Foster, J.; Lee, Y.H.; Tegeder, M. Distinct expression of members of the LHT amino acid transporter family in flowers indicates specific roles in plant reproduction. Sex. Plant Reprod. 2008, 21, 143–152. [Google Scholar] [CrossRef]
- Patrick, J.W.; Stoddard, F.L. Physiology of flowering and grain filling in faba bean. Field Crop. Res. 2010, 115, 234–242. [Google Scholar] [CrossRef]
- Lee, Y.H.; Tegeder, M. Selective expression of a novel high-affinity transport system for acidic and neutral amino acids in the tapetum cells of Arabidopsis flowers. Plant J. 2004, 40, 60–74. [Google Scholar] [CrossRef]
- Werner, D.; Gerlitz, N.; Stadler, R. A dual switch in phloem unloading during ovule development in Arabidopsis. Protoplasma 2011, 248, 225–235. [Google Scholar] [CrossRef]
- Karmann, J.; Müller, B.; Hammes, U.Z. The long and winding road: Transport pathways for amino acids in Arabidopsis seeds. Plant Reprod. 2018, 31, 253–261. [Google Scholar] [CrossRef]
- Müller, B.; Fastner, A.; Karmann, J.; Mansch, V.; Hoffmann, T.; Schwab, W.; Suter-Grotemeyer, M.; Rentsch, D.; Truernit, E.; Ladwig, F.; et al. Amino acid export in developing Arabidopsis seeds depends on umamit facilitators. Curr. Biol. 2015, 25, 3126–3131. [Google Scholar] [CrossRef] [Green Version]
- Besnard, J.; Zhao, C.; Avice, J.C.; Vitha, S.; Hyodo, A.; Pilot, G.; Okumoto, S. Arabidopsis UMAMIT24 and 25 are amino acid exporters involved in seed loading. J. Exp. Bot. 2018, 69, 5221–5232. [Google Scholar] [CrossRef]
- Sanders, A.; Collier, R.; Trethewy, A.; Gould, G.; Sieker, R.; Tegeder, M. AAP1 regulates import of amino acids into developing Arabidopsis embryos. Plant J. 2009, 59, 540–552. [Google Scholar] [CrossRef]
- Schmidt, R.; Stransky, H.; Koch, W. The amino acid permease AAP8 is important for early seed development in Arabidopsis thaliana. Planta 2007, 226, 805–813. [Google Scholar] [CrossRef]
- Canarini, A.; Kaiser, C.; Merchant, A.; Richter, A.; Kaiser, C. Root exudation of primary metabolites: Mechanisms and their roles in plant responses to environmental stimuli. Front. Plant Sci. 2019, 10. [Google Scholar] [CrossRef] [Green Version]
- Pilot, G.; Stransky, H.; Bushey, D.F.; Pratelli, R.; Ludewig, U.; Wingate, V.P.M.; Frommer, W.B. Overexpression of GLUTAMINE DUMPER1 leads to hypersecretion of glutamine from hydathodes of Arabidopsis leaves. Plant Cell 2004, 16, 1827–1840. [Google Scholar] [CrossRef] [Green Version]
- Pratelli, R.; Pilot, G. The plant-specific VIMAG domain of glutamine dumper1 is necessary for the function of the protein in Arabidopsis. FEBS Lett. 2006, 580, 6961–6966. [Google Scholar] [CrossRef] [Green Version]
- Pratelli, R.; Voll, L.M.; Horst, R.J.; Frommer, W.B.; Pilot, G. Stimulation of nonselective amino acid export by glutamine dumper proteins. Plant Physiol. 2010, 152, 762–773. [Google Scholar] [CrossRef] [Green Version]
- Guerra, D.D.; Pratelli, R.; Kraft, E.; Callis, J.; Pilotcet, G. Functional conservation between mammalian MGRN1 and plant LOG2 ubiquitin ligases. FEBS Lett. 2013, 587, 3400–3405. [Google Scholar] [CrossRef] [Green Version]
- Ranocha, P.; Denancé, N.; Vanholme, R.; Freydier, A.; Martinez, Y.; Hoffmann, L.; Köhler, L.; Pouzet, C.; Renou, J.P.; Sundberg, B.; et al. Walls are thin 1 (WAT1), an Arabidopsis homolog of Medicago truncatula NODULIN21, is a tonoplast-localized protein required for secondary wall formation in fibers. Plant J. 2010, 63, 469–483. [Google Scholar] [CrossRef]
- Dündar, E. Multiple GUS expression patterns of a single Arabidopsis gene. Ann. Appl. Biol. 2008, 154, 33–41. [Google Scholar] [CrossRef]
- Chen, L.; Ortiz-Lopez, A.; Jung, A.; Bush, D.R. ANT1, an aromatic and neutral amino acid, transporter in Arabidopsis. Plant Physiol. 2001, 125, 1813–1820. [Google Scholar] [CrossRef] [Green Version]
- Choi, J.; Eom, S.; Shin, K.; Lee, R.A.; Choi, S.; Lee, J.H.; Lee, S.; Soh, M.S. Identification of lysine histidine transporter 2 as an 1-aminocyclopropane carboxylic acid transporter in Arabidopsis thaliana by transgenic complementation approach. Front. Plant Sci. 2019, 10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Delrot, S.; Atanassova, R.; Maurousset, L. Regulation of sugar, amino acid and peptide plant membrane transporters. Biochim. Biophys. Acta 2000, 1465, 281–306. [Google Scholar] [CrossRef] [Green Version]
- Lu, Y.G.; Song, Z.Y.; Lü, K.; Lian, X.M.; Cai, H.M. Molecular characterization, expression and functional analysis of the amino acid transporter gene family (OsAATs) in rice. Acta Physiol. Plant. 2012, 34, 1943–1962. [Google Scholar] [CrossRef]
- Guo, M.G. Molecular and Genomic Analysis of Nitrogen Regulation of Amino Acid Permease I (AAP1) in Arabidopsis. Ph.D. Thesis, University of Illinois at Urbana-Champaign, Urbana, IL, USA, 2004. [Google Scholar]
- Liu, G.S.; Ji, Y.Y.; Bhuiyan, N.H.; Pilot, G.; Selvaraj, G.; Zou, J.T.; Wei, Y.D. Amino Acid homeostasis modulates salicylic acid-associated redox status and defense responses in Arabidopsis. Plant Cell 2010, 22, 3845–3863. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guether, M.; Volpe, V.; Balestrini, R.; Requena, N.; Wipf, D.; Bonfante, P. LjLHT1.2—A mycorrhiza-inducible plant amino acid transporter from lotus japonicas. Biol. Fertil. Soils 2011, 47, 925–936. [Google Scholar] [CrossRef] [Green Version]
- Zhang, R.; Zhu, J.; Cao, H.Z.; Xie, X.L.; Huang, J.J.; Chen, X.H.; Luo, Z.Y. Isolation and characterization of LHT-type plant amino acid transporter gene from panax ginseng meyer. J. Ginseng Res. 2013, 37, 361–370. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Iqbal, A.; Dong, Q.; Wang, X.R.; Gui, H.P.; Zhang, H.H.; Zhang, X.L.; Song, M.Z. Transcriptome analysis reveals differences in key genes and pathways regulating carbon and nitrogen metabolism in cotton genotypes under n starvation and resupply. Int. J. Mol. Sci. 2020, 21, 1500. [Google Scholar] [CrossRef] [Green Version]
- Perchlik, M.; Tegeder, M. Leaf amino acid supply affects photosynthetic and plant nitrogen use efficiency under nitrogen stress. Plant Physiol. 2018, 178, 174–188. [Google Scholar] [CrossRef] [Green Version]
- Foyer, C.H.; Parry, M.; Noctor, G. Markers and signals associated with nitrogen assimilation in higher plants. J. Exp. Bot. 2003, 54, 585–593. [Google Scholar] [CrossRef]
- Bolchi, A.; Petrucco, S.; Tenca, P.L.; Foroni, C.; Ottonello, S. Coordinate modulation of maize sulfate permease and ATP sulfurylase mRNAs in response to variations in sulfur nutritional status: Stereospecific down-regulation by L-cysteine. Plant Mol. Biol. 1999, 39, 527–537. [Google Scholar] [CrossRef]


{kind=link}
| Locus | Gene Name | Substrate (Expression in Yeast or/and Xenopus Oocytes) | Tissue Localization | Subcellular Localization | Phenotype | References |
|---|---|---|---|---|---|---|
| At2g40900 | UMAMIT11 | Glu and Gln | roots, leaves, flowers, throughout seed development | plasma membrane | umamit11: Levels of free amino acids in fruits (Asp, Thr, Glu, Ala)↑; Seed volume↓ | [100] |
| At2g39510 | UMAMIT14 | broad range of amino acids | roots, leaves, flowers, throughout seed development | plasma membrane | umamit14: Levels of free amino acids in fruits (Ser, Asp, Thr, Gln, Glu, Asn, Pro, Ala, GABA)↑; Seed volume;↓ amino acid transferred to the roots and secreted by the roots↓ | [8,100] |
| Atlg44800 | UMAMIT18 (SIAR1) | Gln, Asp, Ala, Asn, Thr, Val, His, Leu | pericycle, stamen, developing seeds, roots | plasma membrane | siarl mutants: Early stages of silique development (amino acid content↓; anthocyanins↑); amino acid transferred to the roots and secreted by the roots↓ | [8,17] |
| At1g25270 | UMAMIT24 | broad range of amino acids | developing seeds (seed coat) | tonoplast | umamit24 knockout: Amino acid contents in seeds↓ | [101] |
| At1g09380 | UMAMIT25 | broad range of amino acids | developing seeds (endosperm cells) | plasma membrane | umamit25 knockout: Amino acid contents in seeds↓ | [101] |
| At1g01070 | UMAMIT28 | Glu and Gln | roots, leaves, flowers, later in seed development | plasma membrane | umamit28: Levels of free amino acids in fruits (Asn, Pro, Ala, GABA)↑; Seed volume↓ | [100] |
| At4g01430 | UMAMIT29 | Glu and Gln | roots, leaves, flowers, before late torpedo stage | plasma membrane | umamit29: Levels of free amino acids in fruits (Ser, Thr, Gln, Glu, Asn, Pro, Ala, GABA)↑; Seed volume↓ | [100] |
| At1g75500 | UMAMIT5 (WAT1) | IAA | developing xylem vessels and fibers | tonoplast | wat1 mutants: A defect in cell elongation; no secondary cell walls in fibers | [18,109] |
| At2g01170 | BAT1 | Ala, Arg, Glu, Lys | vascular tissues | plasma membrane | — | [39,110] |
| At3g11900 | ANT1 | aromatic and neutral amino acid, Arg | flowers and cauline leaves | ER in the perinuclear region | ant1 mutants: Essential amino acids within the SEs↑ | [78,88,111] |
| At5g65990 | AVT3A | neutral amino acids | various tissues of whole plants | tonoplast | — | [78] |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yao, X.; Nie, J.; Bai, R.; Sui, X. Amino Acid Transporters in Plants: Identification and Function. Plants 2020, 9, 972. https://doi.org/10.3390/plants9080972
Yao X, Nie J, Bai R, Sui X. Amino Acid Transporters in Plants: Identification and Function. Plants. 2020; 9(8):972. https://doi.org/10.3390/plants9080972
Chicago/Turabian StyleYao, Xuehui, Jing Nie, Ruoxue Bai, and Xiaolei Sui. 2020. "Amino Acid Transporters in Plants: Identification and Function" Plants 9, no. 8: 972. https://doi.org/10.3390/plants9080972
APA StyleYao, X., Nie, J., Bai, R., & Sui, X. (2020). Amino Acid Transporters in Plants: Identification and Function. Plants, 9(8), 972. https://doi.org/10.3390/plants9080972