
Inhibition of CK2 Reduces NG2 Expression in Juvenile Angiofibroma

 , , and
, , and 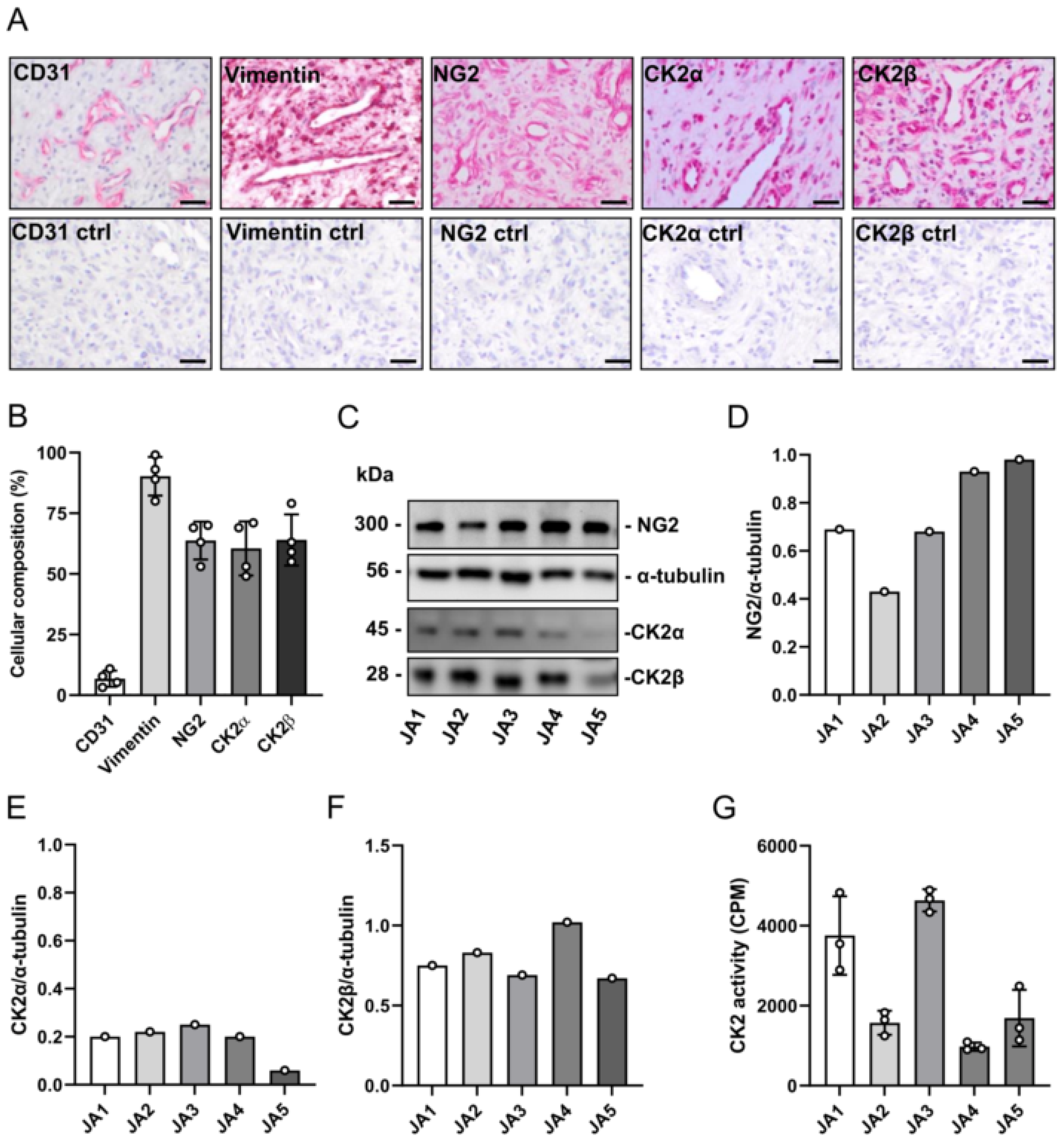{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Antibodies
2.2. Patient-Derived JA Tissue Samples
2.3. Cell Culture
2.4. Immunohistology
2.5. Western Blot Analysis
2.6. CK2 Phosphorylation Assay
2.7. Flow Cytometry
2.8. Water-Soluble Tetrazolium (WST)-1 Assay
2.9. Quantitative Real-Time PCR (qRT-PCR)
2.10. Spheroid Sprouting Assay
2.11. Statistical Analysis
3. Results
3.1. Expression of CK2 and NG2 in Patient-Derived JA Tissue Samples
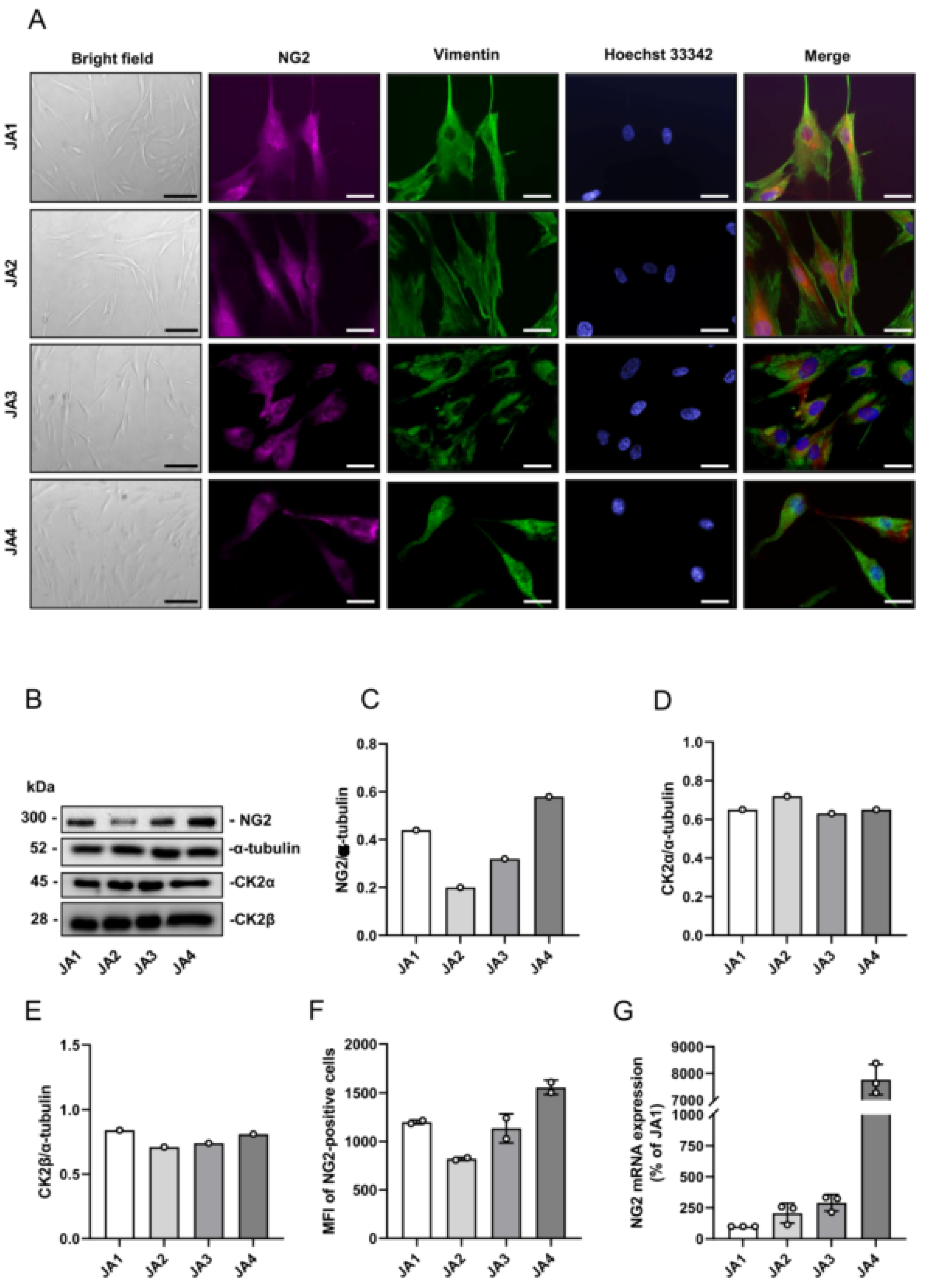
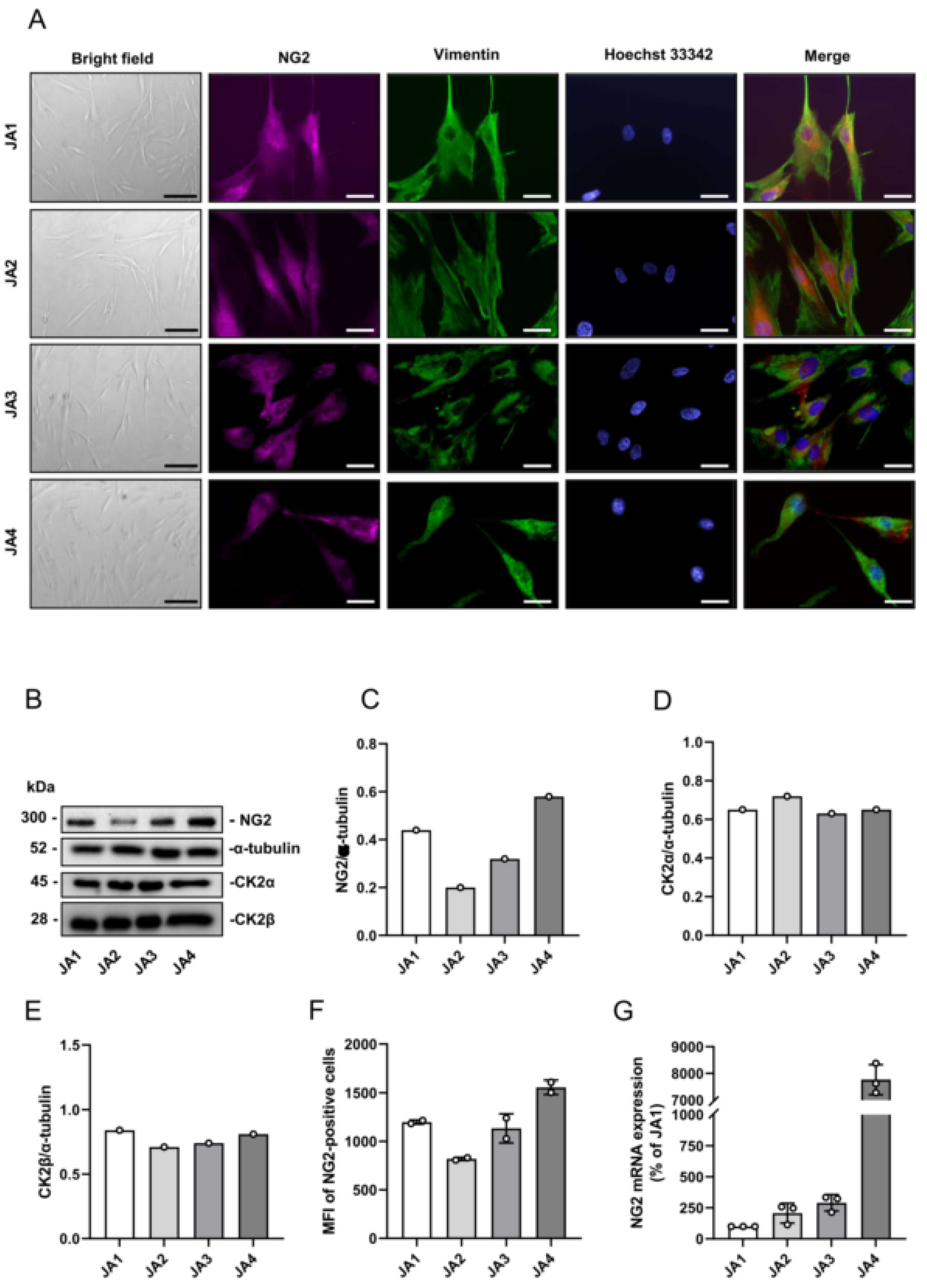
3.2. Expression of CK2 and NG2 in Patient-Derived JA Cells
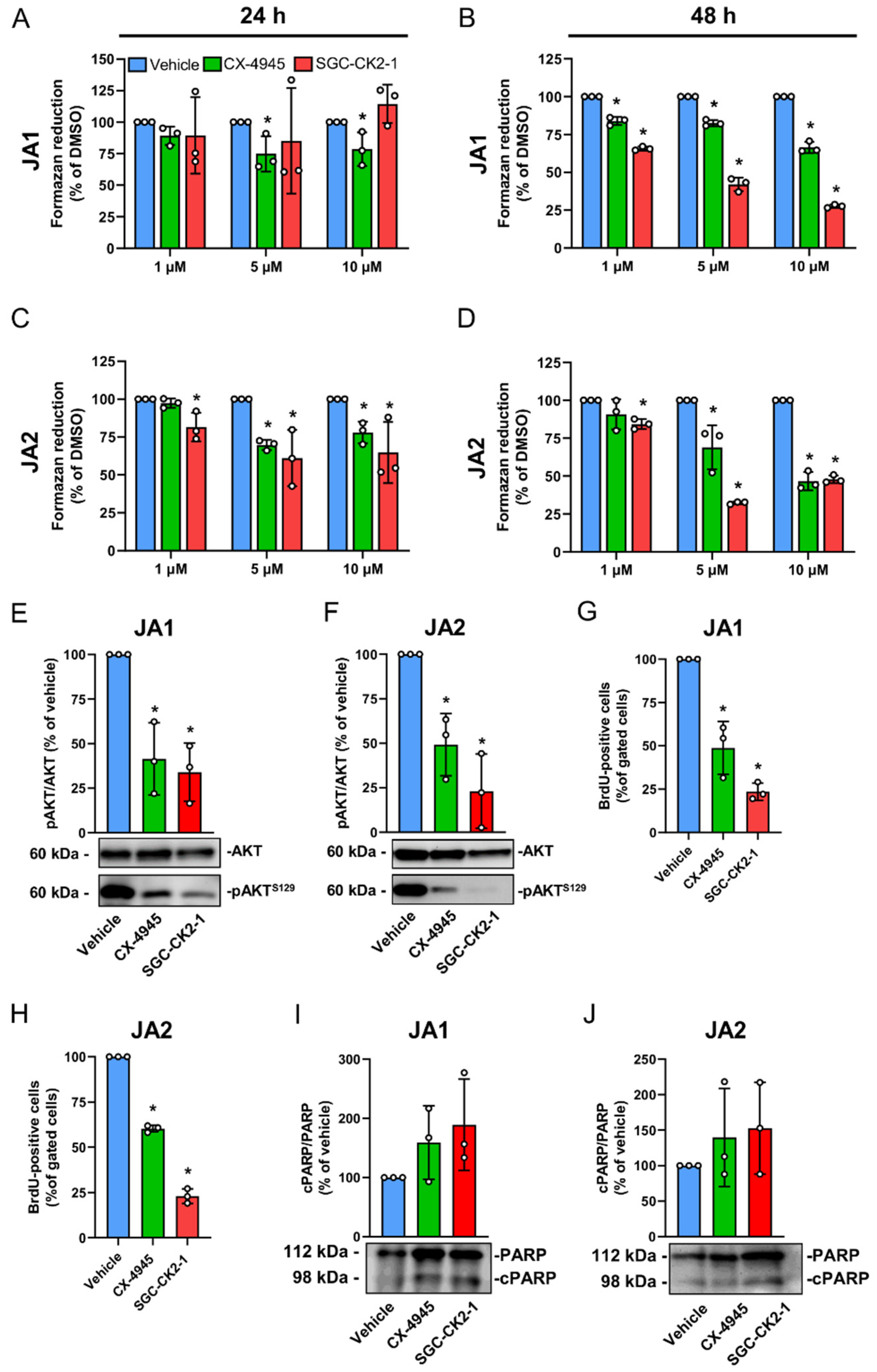
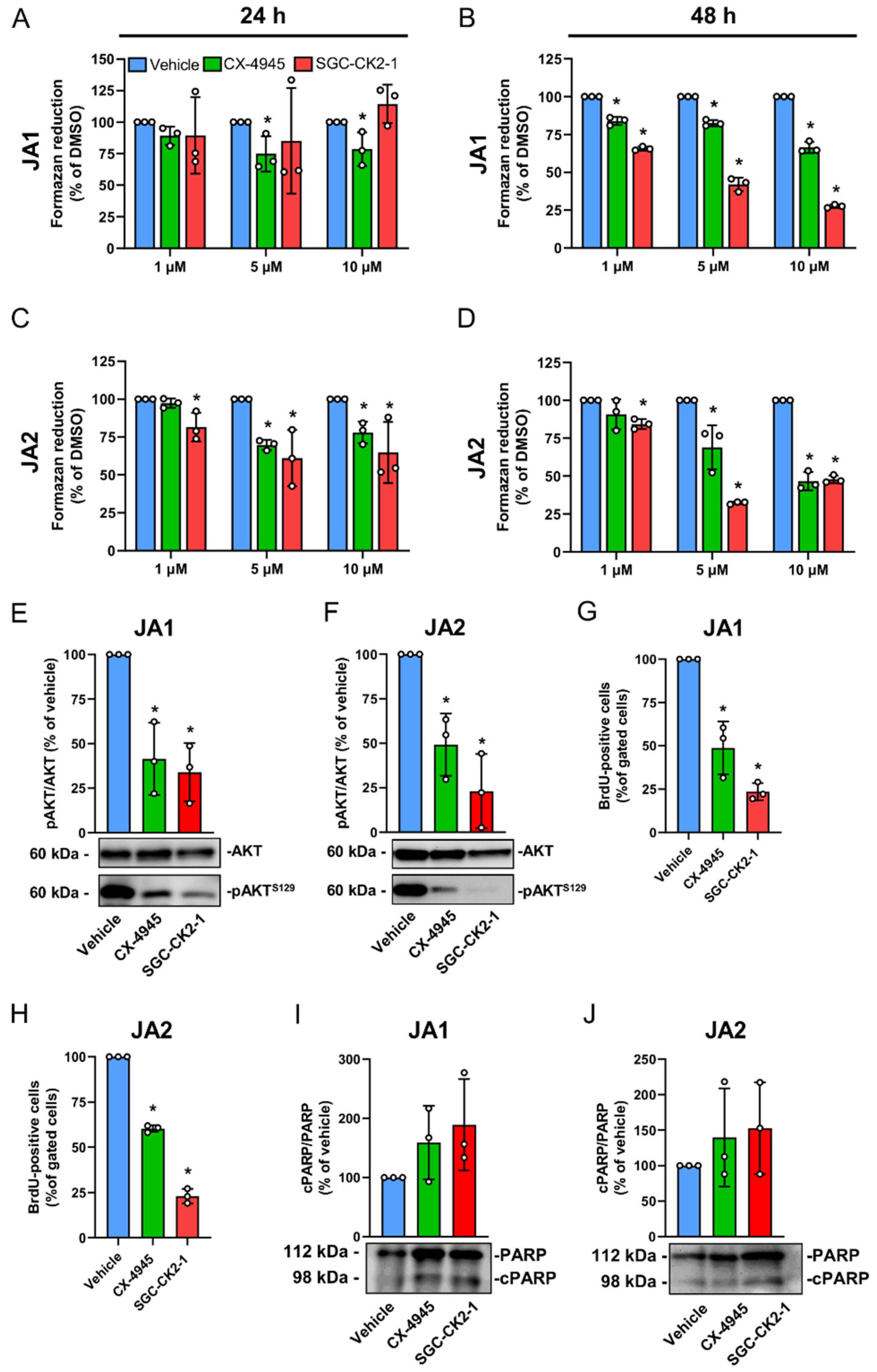
3.3. CK2 Inhibition Suppresses the Proliferation of Patient-Derived JA Cells
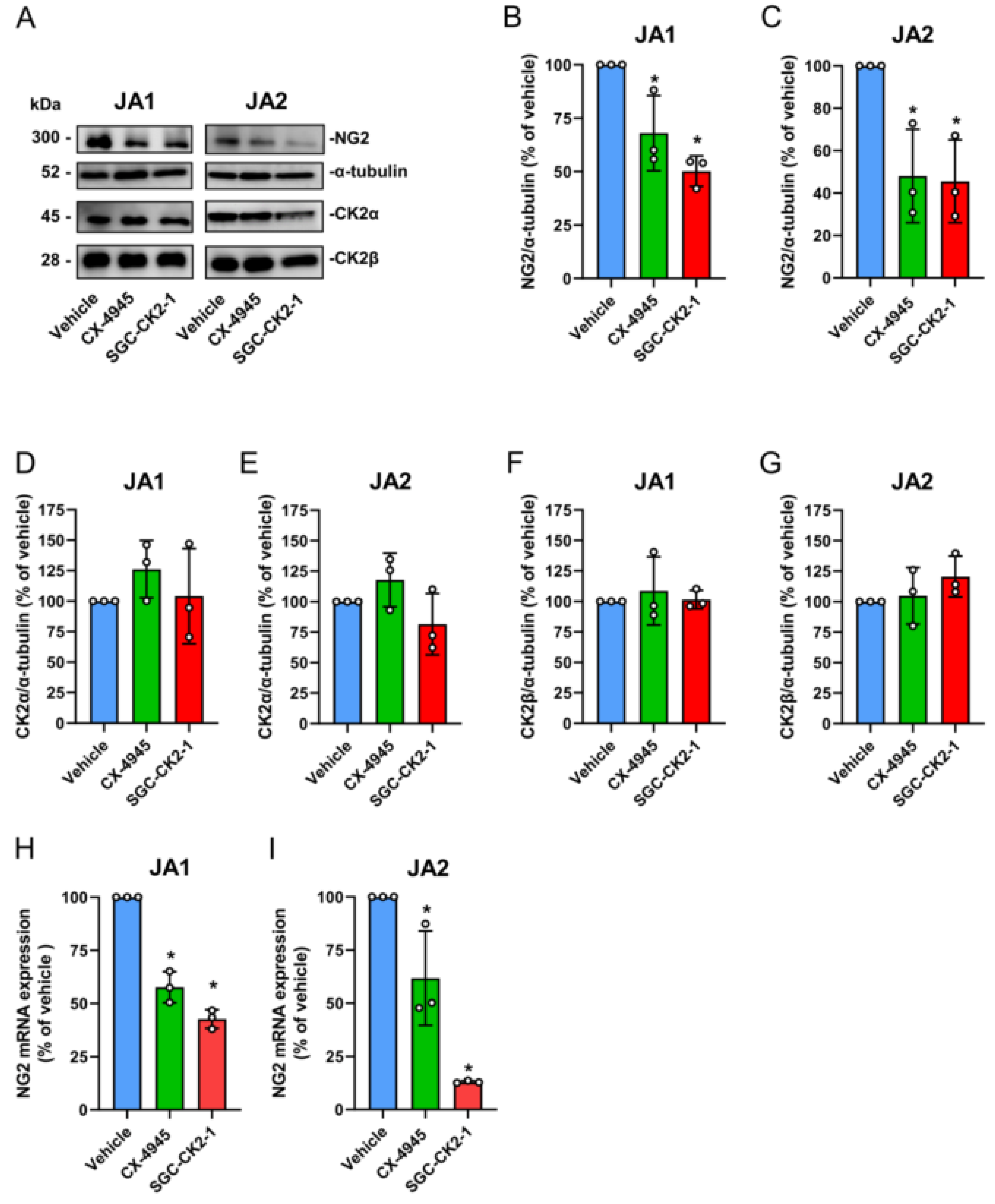
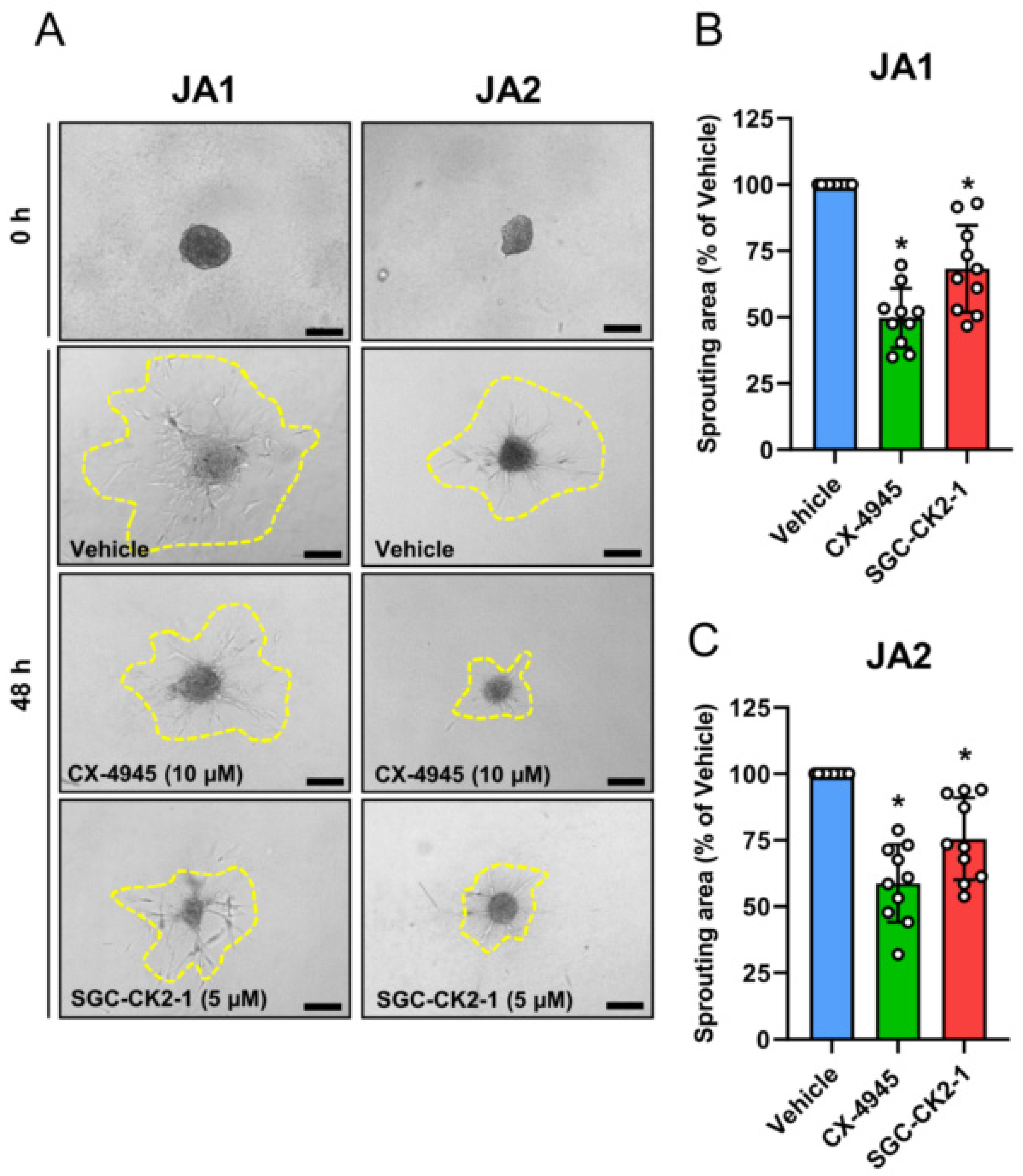
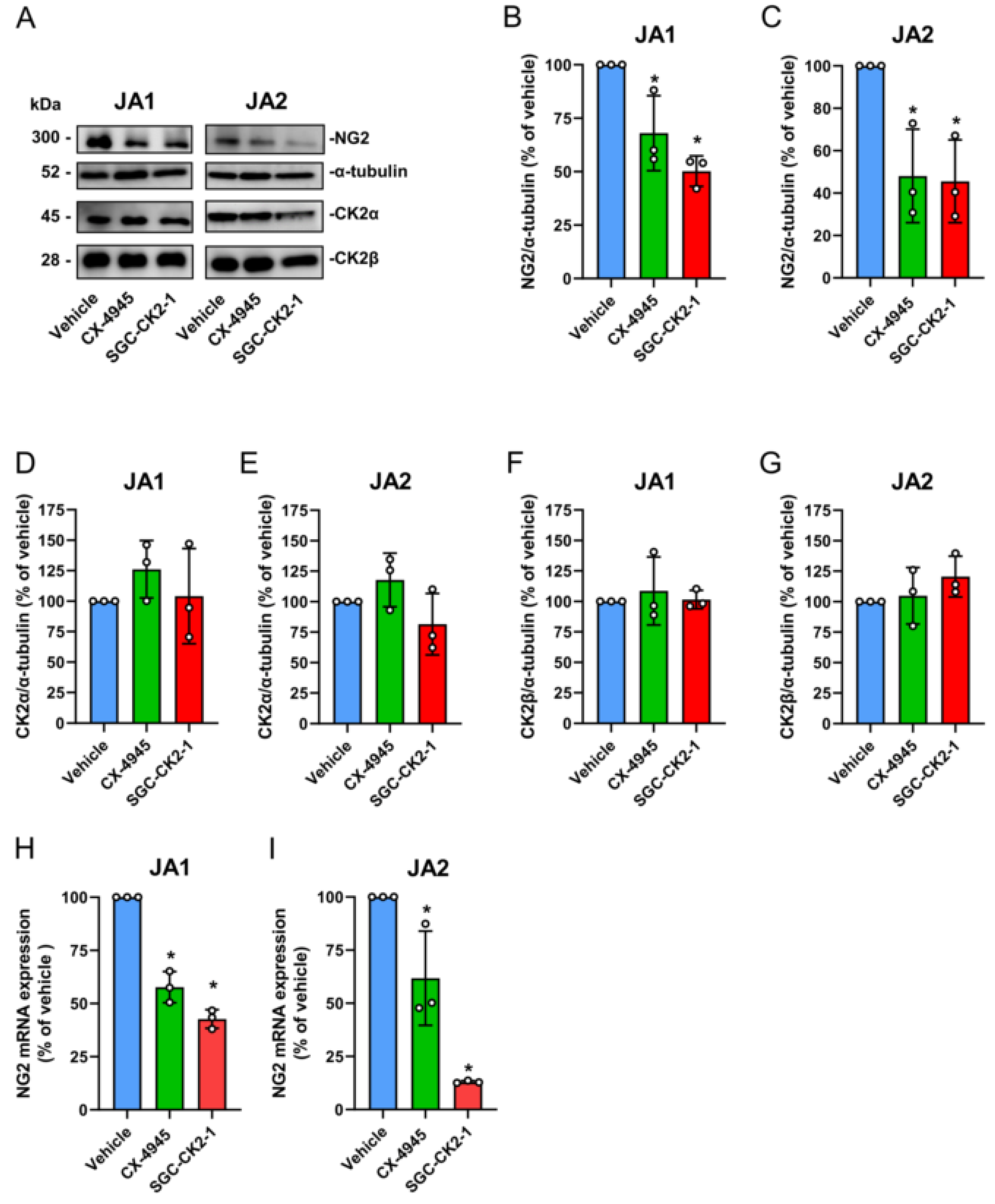
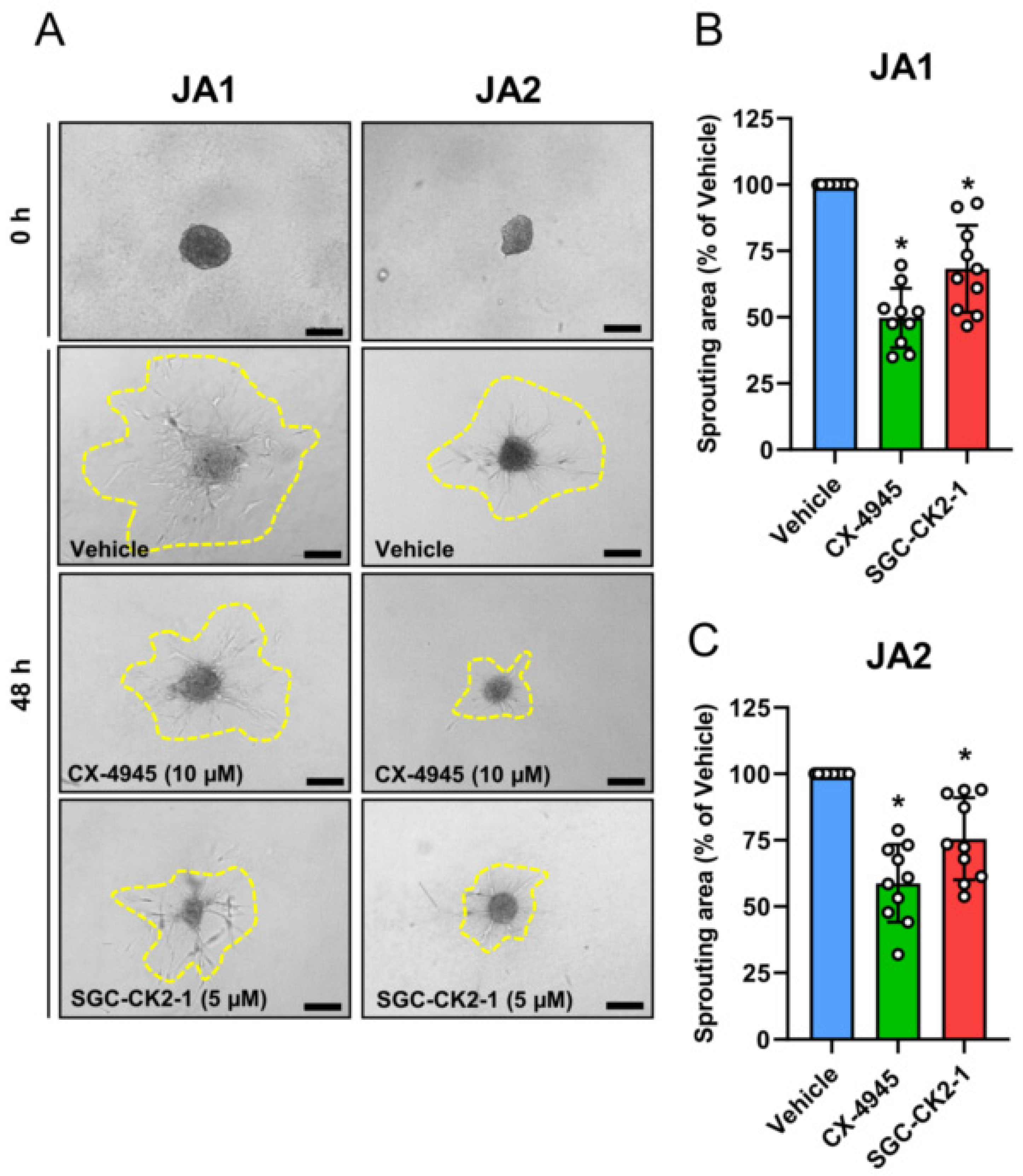
3.4. CK2 Inhibition Reduces NG2 Expression and the Migratory Capacity of Patient-Derived JA Cells
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Li, W.; Ni, Y.; Lu, H.; Hu, L.; Wang, D. Current perspectives on the origin theory of juvenile nasopharyngeal angiofibroma. Discov. Med. 2019, 27, 245–254. [Google Scholar]
- Alshaikh, N.A.; Eleftheriadou, A. Juvenile nasopharyngeal angiofibroma staging: An overview. Ear Nose Throat J. 2015, 94, E12–E22. [Google Scholar] [CrossRef] [Green Version]
- Overdevest, J.B.; Amans, M.R.; Zaki, P.; Pletcher, S.D.; El-Sayed, I.H. Patterns of vascularization and surgical morbidity in juvenile nasopharyngeal angiofibroma: A case series, systematic review, and meta-analysis. Head Neck 2018, 40, 428–443. [Google Scholar] [CrossRef]
- Mehan, R.; Rupa, V.; Lukka, V.K.; Ahmed, M.; Moses, V.; Shyam Kumar, N.K. Association between vascular supply, stage and tumour size of juvenile nasopharyngeal angiofibroma. Eur. Arch. Oto-Rhino-Laryngol. 2016, 273, 4295–4303. [Google Scholar] [CrossRef]
- Min, H.J.; Chung, H.J.; Kim, C.H. Delayed cerebrospinal fluid rhinorrhea four years after gamma knife surgery for juvenile angiofibroma. J. Craniofacial Surg. 2014, 25, e565–e567. [Google Scholar] [CrossRef]
- Sitenga, G.; Granger, P.; Hepola, K.; Aird, J.; Silberstein, P.T. The use of flutamide for the neoadjuvant treatment of juvenile nasopharyngeal angiofibroma: A review of the literature comparing results by pubertal status and tumor stage. Int. J. Dermatol. 2021. online ahead of print. [Google Scholar] [CrossRef]
- Le, T.; New, J.; Jones, J.W.; Usman, S.; Yalamanchali, S.; Tawfik, O.; Hoover, L.; Bruegger, D.E.; Thomas, S.M. Inhibition of fibroblast growth factor receptor with AZD4547 mitigates juvenile nasopharyngeal angiofibroma. Int. Forum Allergy Rhinol. 2017, 7, 973–979. [Google Scholar] [CrossRef]
- Wendler, O.; Dlugaiczyk, J.; Birk, S.; Schick, B. Anti-proliferative effect of glucocorticoids on mesenchymal cells in juvenile angiofibromas. Head Neck 2012, 34, 1615–1621. [Google Scholar] [CrossRef]
- Saylam, G.; Yucel, O.T.; Sungur, A.; Onerci, M. Proliferation, angiogenesis and hormonal markers in juvenile nasopharyngeal angiofibroma. Int. J. Pediatr. Otorhinolaryngol. 2006, 70, 227–234. [Google Scholar] [CrossRef]
- Gramann, M.; Wendler, O.; Haeberle, L.; Schick, B. Prominent collagen type VI expression in juvenile angiofibromas. Histochem. Cell Biol. 2009, 131, 155–164. [Google Scholar] [CrossRef]
- Ampofo, E.; Schmitt, B.M.; Menger, M.D.; Laschke, M.W. The regulatory mechanisms of NG2/CSPG4 expression. Cell Mol. Biol. Lett. 2017, 22, 4. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Burg, M.A.; Nishiyama, A.; Stallcup, W.B. A central segment of the NG2 proteoglycan is critical for the ability of glioma cells to bind and migrate toward type VI collagen. Exp. Cell Res. 1997, 235, 254–264. [Google Scholar] [CrossRef] [PubMed]
- Stallcup, W.B.; Huang, F.J. A role for the NG2 proteoglycan in glioma progression. Cell Adhes. Migr. 2008, 2, 192–201. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ferrara, G.; Errede, M.; Girolamo, F.; Morando, S.; Ivaldi, F.; Panini, N.; Bendotti, C.; Perris, R.; Furlan, R.; Virgintino, D.; et al. NG2, a common denominator for neuroinflammation, blood-brain barrier alteration, and oligodendrocyte precursor response in EAE, plays a role in dendritic cell activation. Acta Neuropathol. 2016, 132, 23–42. [Google Scholar] [CrossRef] [Green Version]
- Fukushi, J.; Makagiansar, I.T.; Stallcup, W.B. NG2 proteoglycan promotes endothelial cell motility and angiogenesis via engagement of galectin-3 and alpha3beta1 integrin. Mol. Biol. Cell 2004, 15, 3580–3590. [Google Scholar] [CrossRef] [Green Version]
- Ozerdem, U.; Stallcup, W.B. Pathological angiogenesis is reduced by targeting pericytes via the NG2 proteoglycan. Angiogenesis 2004, 7, 269–276. [Google Scholar] [CrossRef] [Green Version]
- Rivera, Z.; Ferrone, S.; Wang, X.; Jube, S.; Yang, H.; Pass, H.I.; Kanodia, S.; Gaudino, G.; Carbone, M. CSPG4 as a target of antibody-based immunotherapy for malignant mesothelioma. Clin. Cancer Res. 2012, 18, 5352–5363. [Google Scholar] [CrossRef] [Green Version]
- Schmitt, B.M.; Laschke, M.W.; Roessler, O.G.; Huang, W.; Scheller, A.; Menger, M.D.; Ampofo, E. Nerve/glial antigen (NG) 2 is a crucial regulator of intercellular adhesion molecule (ICAM)-1 expression. Biochim. Biophys. Acta 2018, 1865, 57–66. [Google Scholar] [CrossRef]
- Schmitt, B.M.; Boewe, A.S.; Becker, V.; Nalbach, L.; Gu, Y.; Götz, C.; Menger, M.D.; Laschke, M.W.; Ampofo, E. Protein Kinase CK2 Regulates Nerve/Glial Antigen (NG)2-Mediated Angiogenic Activity of Human Pericytes. Cells 2020, 9, 1546. [Google Scholar] [CrossRef]
- Schmitt, B.M.; Boewe, A.S.; Götz, C.; Philipp, S.E.; Urbschat, S.; Oertel, J.; Menger, M.D.; Laschke, M.W.; Ampofo, E. CK2 Activity Mediates the Aggressive Molecular Signature of Glioblastoma Multiforme by Inducing Nerve/Glial Antigen (NG)2 Expression. Cancers 2021, 13, 1678. [Google Scholar] [CrossRef]
- Castello, J.; Ragnauth, A.; Friedman, E.; Rebholz, H. CK2-An Emerging Target for Neurological and Psychiatric Disorders. Pharmaceuticals 2017, 10, 7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roffey, S.E.; Litchfield, D.W. CK2 Regulation: Perspectives in 2021. Biomedicines 2021, 9, 1361. [Google Scholar] [CrossRef] [PubMed]
- Chua, M.M.J.; Lee, M.; Dominguez, I. Cancer-type dependent expression of CK2 transcripts. PLoS ONE 2017, 12, e0188854. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zheng, Y.; McFarland, B.C.; Drygin, D.; Yu, H.; Bellis, S.L.; Kim, H.; Bredel, M.; Benveniste, E.N. Targeting protein kinase CK2 suppresses prosurvival signaling pathways and growth of glioblastoma. Clin. Cancer Res. 2013, 19, 6484–6494. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yenice, S.; Davis, A.T.; Goueli, S.A.; Akdas, A.; Limas, C.; Ahmed, K. Nuclear casein kinase 2 (CK-2) activity in human normal, benign hyperplastic, and cancerous prostate. Prostate 1994, 24, 11–16. [Google Scholar] [CrossRef] [PubMed]
- Perea, S.E.; Baladron, I.; Garcia, Y.; Perera, Y.; Lopez, A.; Soriano, J.L.; Batista, N.; Palau, A.; Hernandez, I.; Farina, H.; et al. CIGB-300, a synthetic peptide-based drug that targets the CK2 phosphoaceptor domain. Translational and clinical research. Mol. Cell. Biochem. 2011, 356, 45–50. [Google Scholar] [CrossRef] [PubMed]
- Wells, C.I.; Drewry, D.H.; Pickett, J.E.; Tjaden, A.; Kramer, A.; Muller, S.; Gyenis, L.; Menyhart, D.; Litchfield, D.W.; Knapp, S.; et al. Development of a potent and selective chemical probe for the pleiotropic kinase CK2. Cell Chem. Biol. 2021, 28, 546–558.e10. [Google Scholar] [CrossRef]
- Siddiqui-Jain, A.; Drygin, D.; Streiner, N.; Chua, P.; Pierre, F.; O’Brien, S.E.; Bliesath, J.; Omori, M.; Huser, N.; Ho, C.; et al. CX-4945, an orally bioavailable selective inhibitor of protein kinase CK2, inhibits prosurvival and angiogenic signaling and exhibits antitumor efficacy. Cancer Res. 2010, 70, 10288–10298. [Google Scholar] [CrossRef] [Green Version]
- Faust, M.; Schuster, N.; Montenarh, M. Specific binding of protein kinase CK2 catalytic subunits to tubulin. FEBS Lett. 1999, 462, 51–56. [Google Scholar] [CrossRef] [Green Version]
- Wemmert, S.; Willnecker, V.; Kulas, P.; Weber, S.; Lerner, C.; Berndt, S.; Wendler, O.; Schick, B. Identification of CTNNB1 mutations, CTNNB1 amplifications, and an Axin2 splice variant in juvenile angiofibromas. Tumour Biol. 2016, 37, 5539–5549. [Google Scholar] [CrossRef]
- Ampofo, E.; Rudzitis-Auth, J.; Dahmke, I.N.; Rössler, O.G.; Thiel, G.; Montenarh, M.; Menger, M.D.; Laschke, M.W. Inhibition of protein kinase CK2 suppresses tumor necrosis factor (TNF)-alpha-induced leukocyte-endothelial cell interaction. Biochim. Biophys. Acta 2015, 1852, 2123–2136. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Al-Mayhani, M.T.; Grenfell, R.; Narita, M.; Piccirillo, S.; Kenney-Herbert, E.; Fawcett, J.W.; Collins, V.P.; Ichimura, K.; Watts, C. NG2 expression in glioblastoma identifies an actively proliferating population with an aggressive molecular signature. Neuro-Oncology 2011, 13, 830–845. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schiffer, D.; Mellai, M.; Boldorini, R.; Bisogno, I.; Grifoni, S.; Corona, C.; Bertero, L.; Cassoni, P.; Casalone, C.; Annovazzi, L. The Significance of Chondroitin Sulfate Proteoglycan 4 (CSPG4) in Human Gliomas. Int. J. Mol. Sci. 2018, 19, 2724. [Google Scholar] [CrossRef] [Green Version]
- Midilli, R.; Karci, B.; Akyildiz, S. Juvenile nasopharyngeal angiofibroma: Analysis of 42 cases and important aspects of endoscopic approach. Int. J. Pediatr. Otorhinolaryngol. 2009, 73, 401–408. [Google Scholar] [CrossRef] [PubMed]
- Intemann, J.; Saidu, N.E.; Schwind, L.; Montenarh, M. ER stress signaling in ARPE-19 cells after inhibition of protein kinase CK2 by CX-4945. Cell. Signal. 2014, 26, 1567–1575. [Google Scholar] [CrossRef] [PubMed]
- Cozza, G.; Mazzorana, M.; Papinutto, E.; Bain, J.; Elliott, M.; di Maira, G.; Gianoncelli, A.; Pagano, M.A.; Sarno, S.; Ruzzene, M.; et al. Quinalizarin as a potent, selective and cell-permeable inhibitor of protein kinase CK2. Biochem. J. 2009, 421, 387–395. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Götz, C.; Gratz, A.; Kucklaender, U.; Jose, J. TF—A novel cell-permeable and selective inhibitor of human protein kinase CK2 induces apoptosis in the prostate cancer cell line LNCaP. Biochim. Biophys. Acta 2012, 1820, 970–977. [Google Scholar] [CrossRef]
- Prins, R.C.; Burke, R.T.; Tyner, J.W.; Druker, B.J.; Loriaux, M.M.; Spurgeon, S.E. CX-4945, a selective inhibitor of casein kinase-2 (CK2), exhibits anti-tumor activity in hematologic malignancies including enhanced activity in chronic lymphocytic leukemia when combined with fludarabine and inhibitors of the B-cell receptor pathway. Leukemia 2013, 27, 2094–2096. [Google Scholar] [CrossRef] [Green Version]
- Sarno, S.; Pinna, L.A. Protein kinase CK2 as a druggable target. Mol. Biosyst. 2008, 4, 889–894. [Google Scholar] [CrossRef]
- Salvi, M.; Borgo, C.; Pinna, L.A.; Ruzzene, M. Targeting CK2 in cancer: A valuable strategy or a waste of time? Cell Death Discov. 2021, 7, 325. [Google Scholar] [CrossRef]
- Di Maira, G.; Salvi, M.; Arrigoni, G.; Marin, O.; Sarno, S.; Brustolon, F.; Pinna, L.A.; Ruzzene, M. Protein kinase CK2 phosphorylates and upregulates Akt/PKB. Cell Death Differ. 2005, 12, 668–677. [Google Scholar] [CrossRef] [PubMed]
- Armstrong, S.A.; Barry, D.A.; Leggett, R.W.; Mueller, C.R. Casein kinase II-mediated phosphorylation of the C terminus of Sp1 decreases its DNA binding activity. J. Biol. Chem. 1997, 272, 13489–13495. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, S.; Kim, K.H. Protein kinase CK2 down-regulates glucose-activated expression of the acetyl-CoA carboxylase gene. Arch. Biochem. Biophys. 1997, 338, 227–232. [Google Scholar] [CrossRef] [PubMed]
- Dupuis-Maurin, V.; Brinza, L.; Baguet, J.; Plantamura, E.; Schicklin, S.; Chambion, S.; Macari, C.; Tomkowiak, M.; Deniaud, E.; Leverrier, Y.; et al. Overexpression of the transcription factor Sp1 activates the OAS-RNAse L-RIG-I pathway. PLoS ONE 2015, 10, e0118551. [Google Scholar] [CrossRef] [PubMed]
- Cattaruzza, S.; Nicolosi, P.A.; Braghetta, P.; Pazzaglia, L.; Benassi, M.S.; Picci, P.; Lacrima, K.; Zanocco, D.; Rizzo, E.; Stallcup, W.B.; et al. NG2/CSPG4-collagen type VI interplays putatively involved in the microenvironmental control of tumour engraftment and local expansion. J. Mol. Cell Biol. 2013, 5, 176–193. [Google Scholar] [CrossRef] [PubMed]
- Ponti, G.; Losi, L.; Pellacani, G.; Rossi, G.B.; Presutti, L.; Mattioli, F.; Villari, D.; Wannesson, L.; Alicandri Ciufelli, M.; Izzo, P.; et al. Wnt pathway, angiogenetic and hormonal markers in sporadic and familial adenomatous polyposis-associated juvenile nasopharyngeal angiofibromas (JNA). Appl. Immunohistochem. Mol. Morphol. 2008, 16, 173–178. [Google Scholar] [CrossRef]
- Calanca, N.; Binato, S.M.S.; da Silva, S.D.; Brentani, H.P.; Sennes, L.U.; Pinto, C.A.L.; Domingues, M.A.C.; Fonseca-Alves, C.E.; Rainho, C.A.; Rogatto, S.R. Master Regulators of Epithelial-Mesenchymal Transition and WNT Signaling Pathways in Juvenile Nasopharyngeal Angiofibromas. Biomedicines 2021, 9, 1258. [Google Scholar] [CrossRef]
- Seldin, D.C.; Landesman-Bollag, E.; Farago, M.; Currier, N.; Lou, D.; Dominguez, I. CK2 as a positive regulator of Wnt signalling and tumourigenesis. Mol. Cell. Biochem. 2005, 274, 63–67. [Google Scholar] [CrossRef]
- Janovska, P.; Bryja, V. Wnt signalling pathways in chronic lymphocytic leukaemia and B-cell lymphomas. Br. J. Pharm. 2017, 174, 4701–4715. [Google Scholar] [CrossRef] [Green Version]
- Martins, L.R.; Perera, Y.; Lucio, P.; Silva, M.G.; Perea, S.E.; Barata, J.T. Targeting chronic lymphocytic leukemia using CIGB-300, a clinical-stage CK2-specific cell-permeable peptide inhibitor. Oncotarget 2014, 5, 258–263. [Google Scholar] [CrossRef] [Green Version]
- Hsu, S.C.; Nadesan, P.; Puviindran, V.; Stallcup, W.B.; Kirsch, D.G.; Alman, B.A. Effects of chondroitin sulfate proteoglycan 4 (NG2/CSPG4) on soft-tissue sarcoma growth depend on tumor developmental stage. J. Biol. Chem. 2018, 293, 2466–2475. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rolih, V.; Barutello, G.; Iussich, S.; De Maria, R.; Quaglino, E.; Buracco, P.; Cavallo, F.; Riccardo, F. CSPG4: A prototype oncoantigen for translational immunotherapy studies. J. Transl. Med. 2017, 15, 151. [Google Scholar] [CrossRef] [PubMed]
- Khan, D.H.; He, S.; Yu, J.; Winter, S.; Cao, W.; Seiser, C.; Davie, J.R. Protein kinase CK2 regulates the dimerization of histone deacetylase 1 (HDAC1) and HDAC2 during mitosis. J. Biol. Chem. 2013, 288, 16518–16528. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bandyopadhyay, K.; Li, P.; Gjerset, R.A. CK2-mediated hyperphosphorylation of topoisomerase I targets serine 506, enhances topoisomerase I-DNA binding, and increases cellular camptothecin sensitivity. PLoS ONE 2012, 7, e50427. [Google Scholar] [CrossRef] [Green Version]
- Schneider, C.C.; Götz, C.; Hessenauer, A.; Gunther, J.; Kartarius, S.; Montenarh, M. Down-regulation of CK2 activity results in a decrease in the level of cdc25C phosphatase in different prostate cancer cell lines. Mol. Cell. Biochem. 2011, 356, 177–184. [Google Scholar] [CrossRef]
- Ahmad, K.A.; Wang, G.; Unger, G.; Slaton, J.; Ahmed, K. Protein kinase CK2--a key suppressor of apoptosis. Adv. Enzym. Regul. 2008, 48, 179–187. [Google Scholar] [CrossRef] [Green Version]
- Wang, G.; Ahmad, K.A.; Ahmed, K. Role of protein kinase CK2 in the regulation of tumor necrosis factor-related apoptosis inducing ligand-induced apoptosis in prostate cancer cells. Cancer Res. 2006, 66, 2242–2249. [Google Scholar] [CrossRef] [Green Version]
- Strum, S.W.; Gyenis, L.; Litchfield, D.W. CSNK2 in cancer: Pathophysiology and translational applications. Br. J. Cancer 2022, 126, 994–1003. [Google Scholar] [CrossRef]
- Pierre, F.; Chua, P.C.; O’Brien, S.E.; Siddiqui-Jain, A.; Bourbon, P.; Haddach, M.; Michaux, J.; Nagasawa, J.; Schwaebe, M.K.; Stefan, E.; et al. Pre-clinical characterization of CX-4945, a potent and selective small molecule inhibitor of CK2 for the treatment of cancer. Mol. Cell. Biochem. 2011, 356, 37–43. [Google Scholar] [CrossRef]
- Pagano, M.A.; Meggio, F.; Ruzzene, M.; Andrzejewska, M.; Kazimierczuk, Z.; Pinna, L.A. 2-Dimethylamino-4,5,6,7-tetrabromo-1H-benzimidazole: A novel powerful and selective inhibitor of protein kinase CK2. Biochem. Biophys. Res. Commun. 2004, 321, 1040–1044. [Google Scholar] [CrossRef]
- Ruzzene, M.; Penzo, D.; Pinna, L.A. Protein kinase CK2 inhibitor 4,5,6,7-tetrabromobenzotriazole (TBB) induces apoptosis and caspase-dependent degradation of haematopoietic lineage cell-specific protein 1 (HS1) in Jurkat cells. Biochem. J. 2002, 364, 41–47. [Google Scholar] [CrossRef] [PubMed] [Green Version]

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Boewe, A.S.; Wemmert, S.; Kulas, P.; Schick, B.; Götz, C.; Wrublewsky, S.; Montenarh, M.; Menger, M.D.; Laschke, M.W.; Ampofo, E. Inhibition of CK2 Reduces NG2 Expression in Juvenile Angiofibroma. Biomedicines 2022, 10, 966. https://doi.org/10.3390/biomedicines10050966
Boewe AS, Wemmert S, Kulas P, Schick B, Götz C, Wrublewsky S, Montenarh M, Menger MD, Laschke MW, Ampofo E. Inhibition of CK2 Reduces NG2 Expression in Juvenile Angiofibroma. Biomedicines. 2022; 10(5):966. https://doi.org/10.3390/biomedicines10050966
Chicago/Turabian StyleBoewe, Anne S., Silke Wemmert, Philipp Kulas, Bernhard Schick, Claudia Götz, Selina Wrublewsky, Mathias Montenarh, Michael D. Menger, Matthias W. Laschke, and Emmanuel Ampofo. 2022. "Inhibition of CK2 Reduces NG2 Expression in Juvenile Angiofibroma" Biomedicines 10, no. 5: 966. https://doi.org/10.3390/biomedicines10050966
APA StyleBoewe, A. S., Wemmert, S., Kulas, P., Schick, B., Götz, C., Wrublewsky, S., Montenarh, M., Menger, M. D., Laschke, M. W., & Ampofo, E. (2022). Inhibition of CK2 Reduces NG2 Expression in Juvenile Angiofibroma. Biomedicines, 10(5), 966. https://doi.org/10.3390/biomedicines10050966