


Analysis of Relative Expression of Key Enzyme Genes and Enzyme Activity in Nitrogen Metabolic Pathway of Two Genotypes of Potato (Solanum tuberosum L.) under Different Nitrogen Supply Levels

Abstract
:1. Introduction
2. Materials and Methods
2.1. Material
2.2. Methods
2.2.1. Test Treatment and Sampling
2.2.2. Obtaining Key Enzyme Gene Sequences
2.2.3. Quantitative Real-Time PCR
2.2.4. Determination of Key Enzyme Activity of N Metabolism
2.2.5. Statistical Analysis
3. Results
3.1. Acquisition of Key Enzyme Genes of Potato N Metabolism Pathway
3.2. Relative Expression Analysis of Key Enzyme Genes in the N Metabolism Pathway of Two Genotypes of Potato under Different N Supply Levels
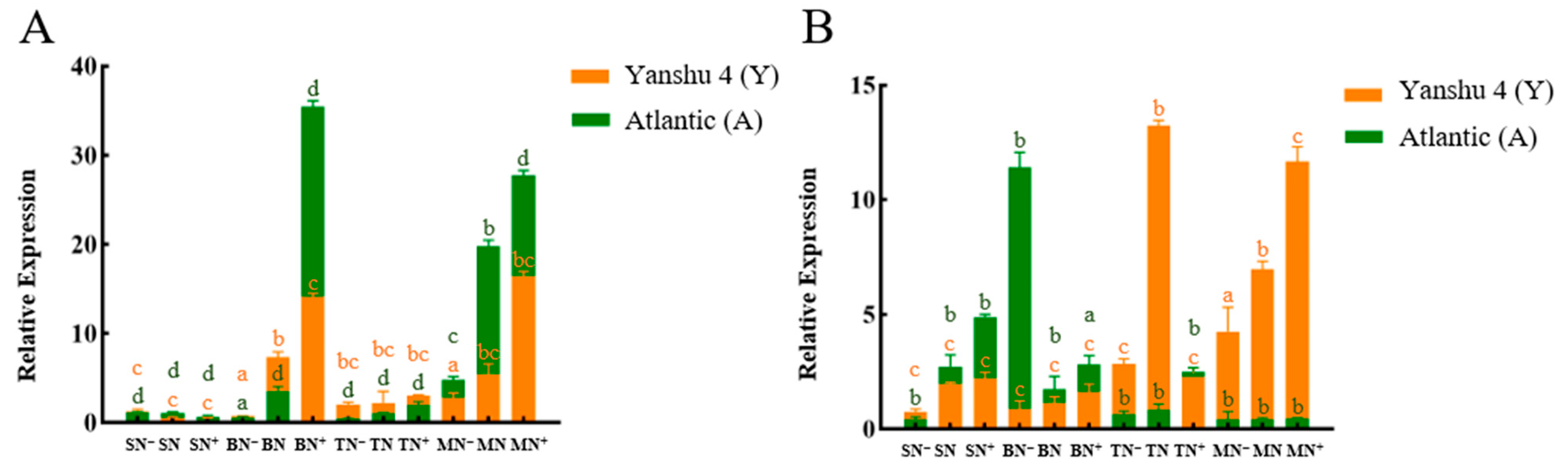
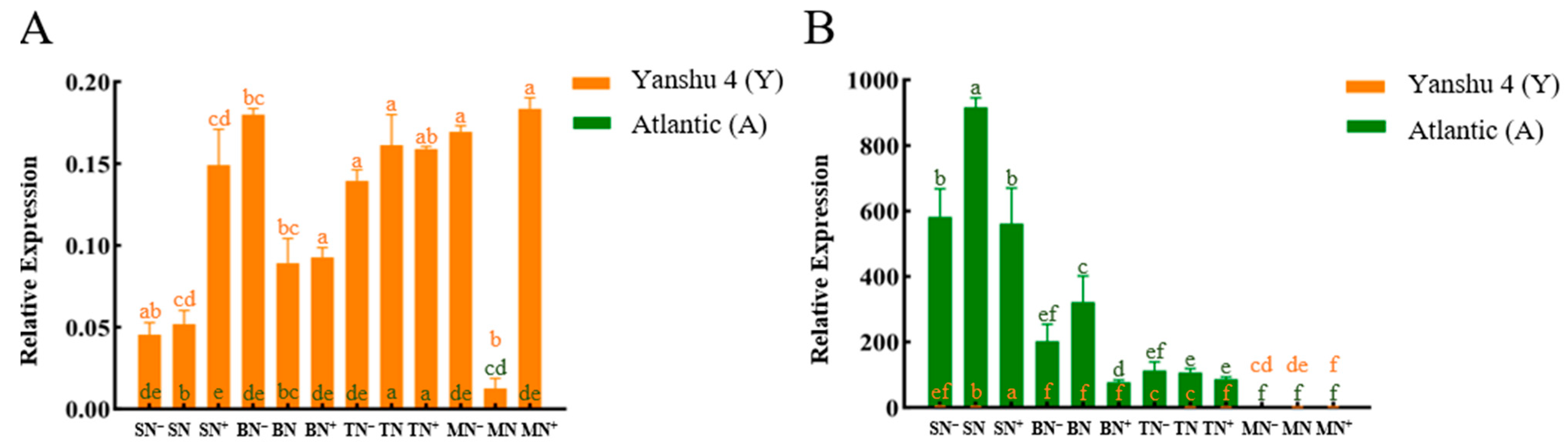
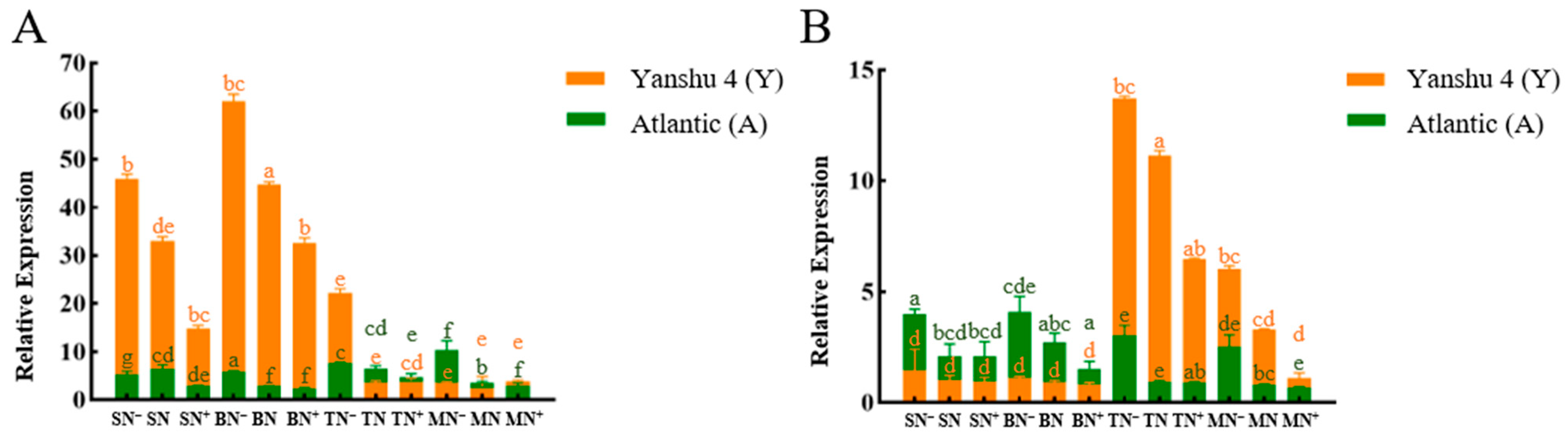
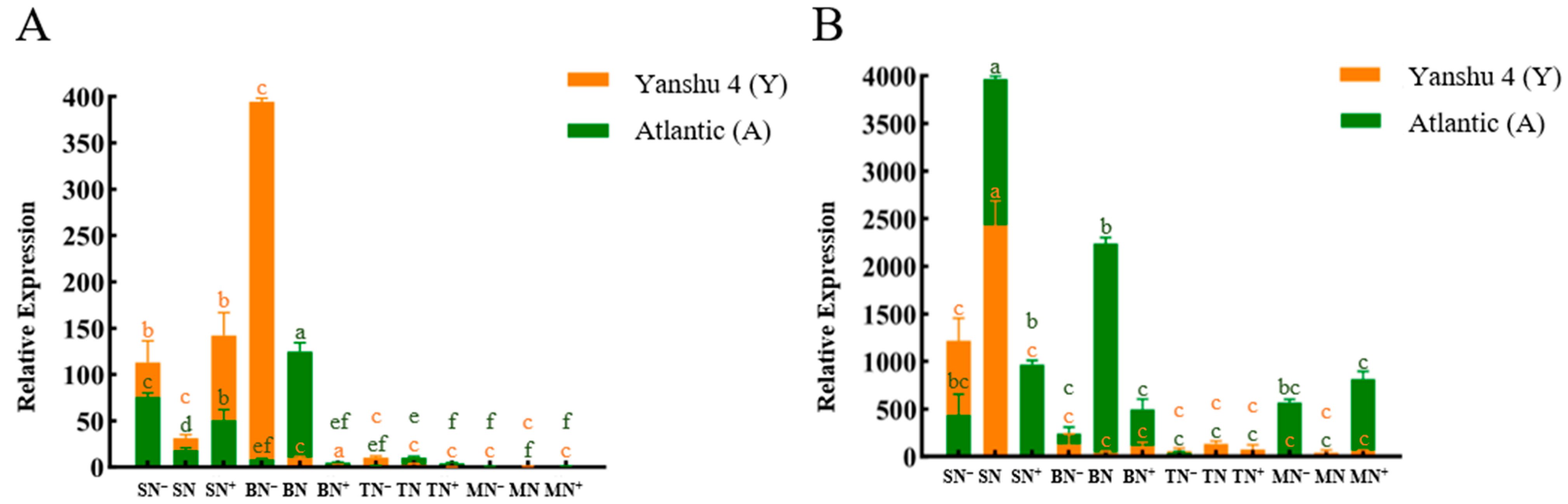
3.2.1. Analysis of Relative Expression of Nitrate Reductase Gene StNR in Two Genotypes of Potato under Different N Supply Levels
3.2.2. Analysis of Relative Expression of Nitrite Reductase Gene StNiR in Two Genotypes of Potato under Different N Supply Levels
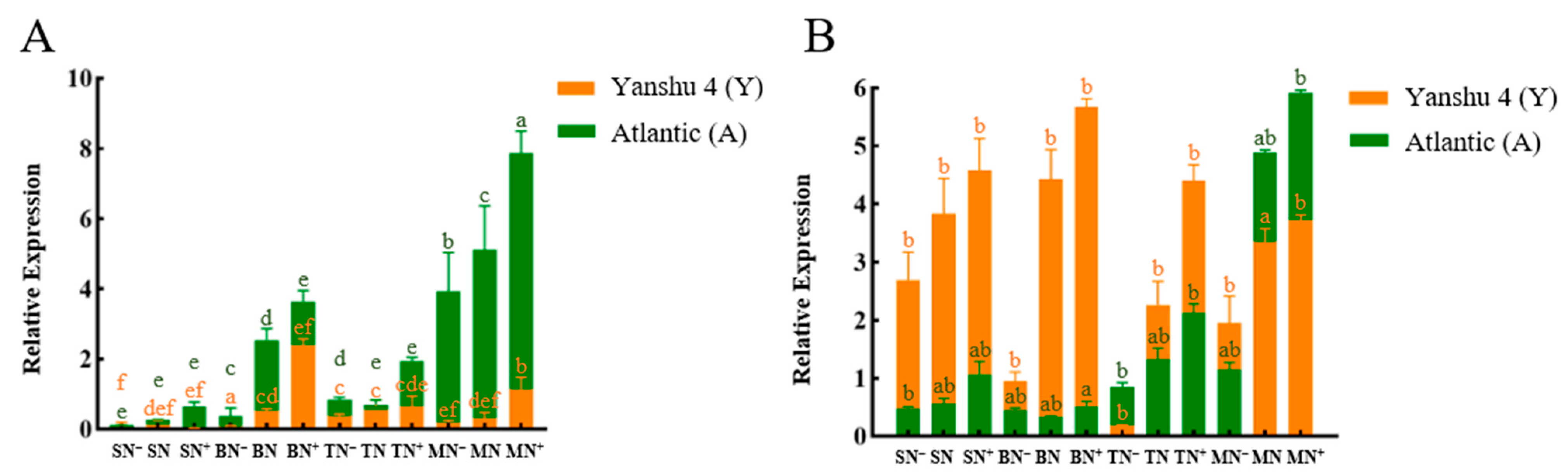
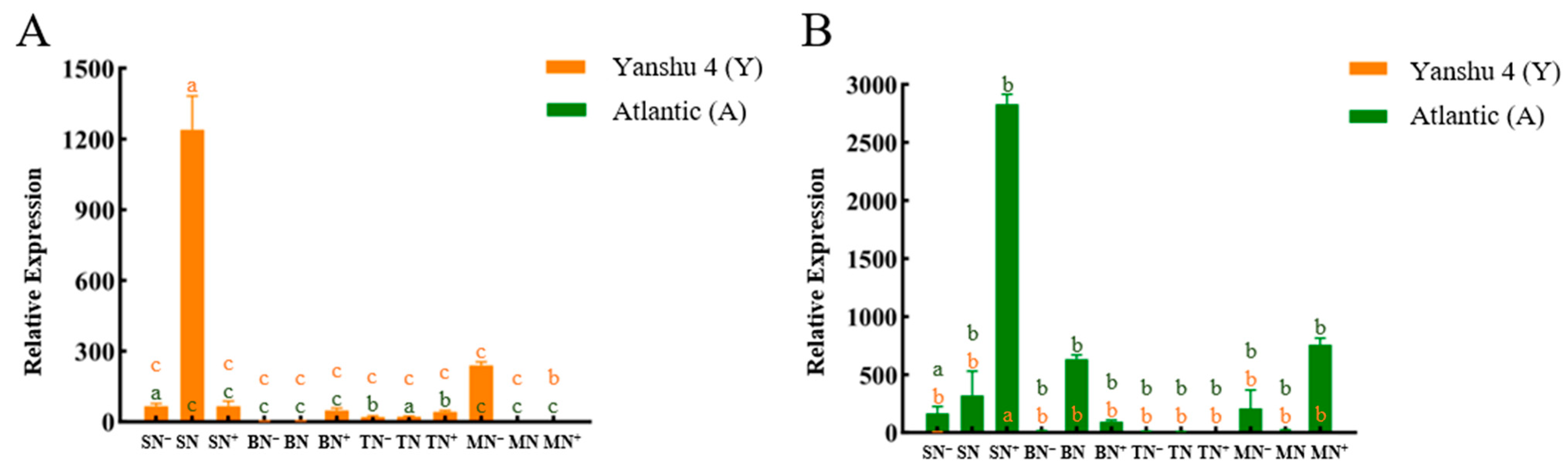
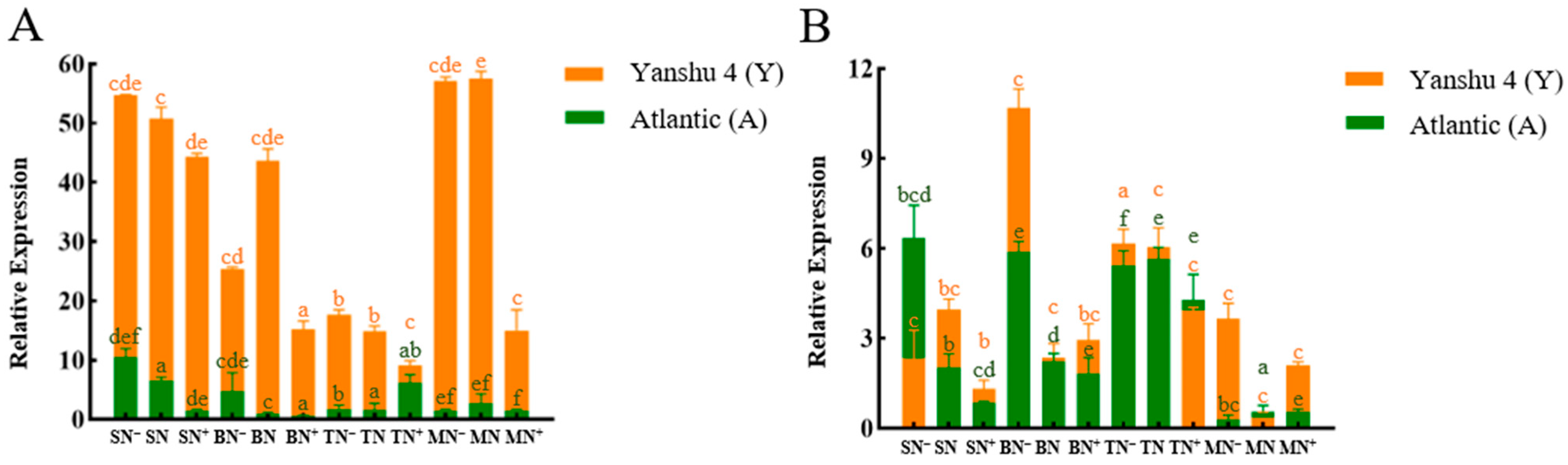
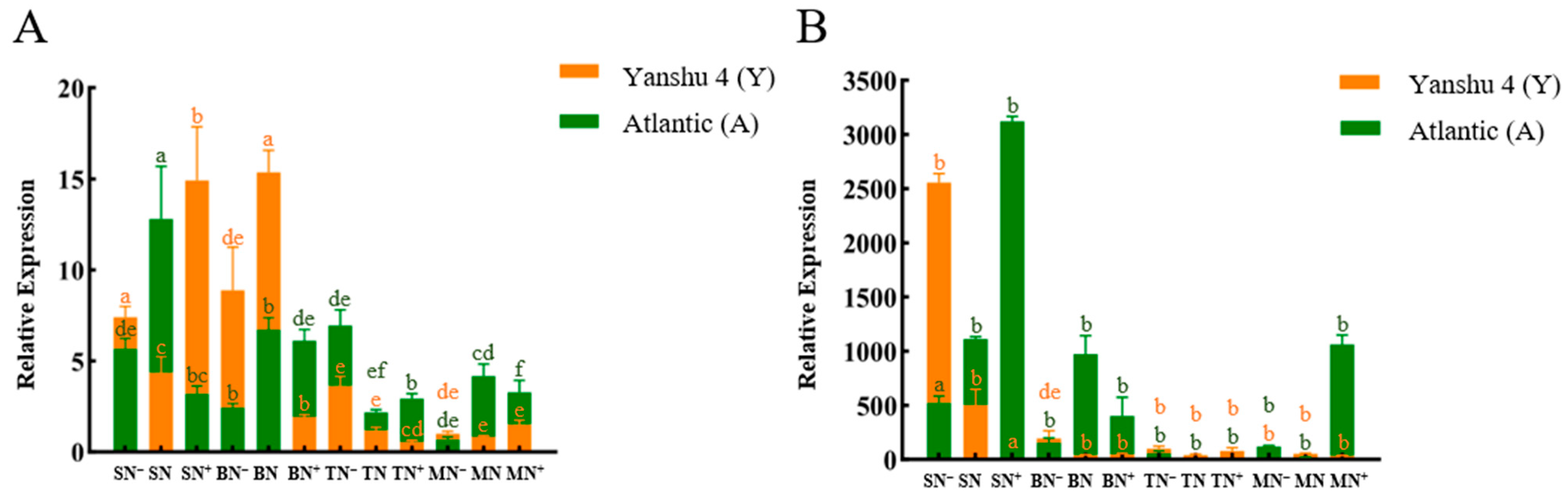
3.2.3. Relative Expression Analysis of Glutamine Synthase Gene StGSs in Two Genotypes of Potato under Different N Supply Levels
3.2.4. Relative Expression Analysis of Glutamate Synthase Gene StGOGATs in the Two Genotypes of Potato under Different N Supply Levels
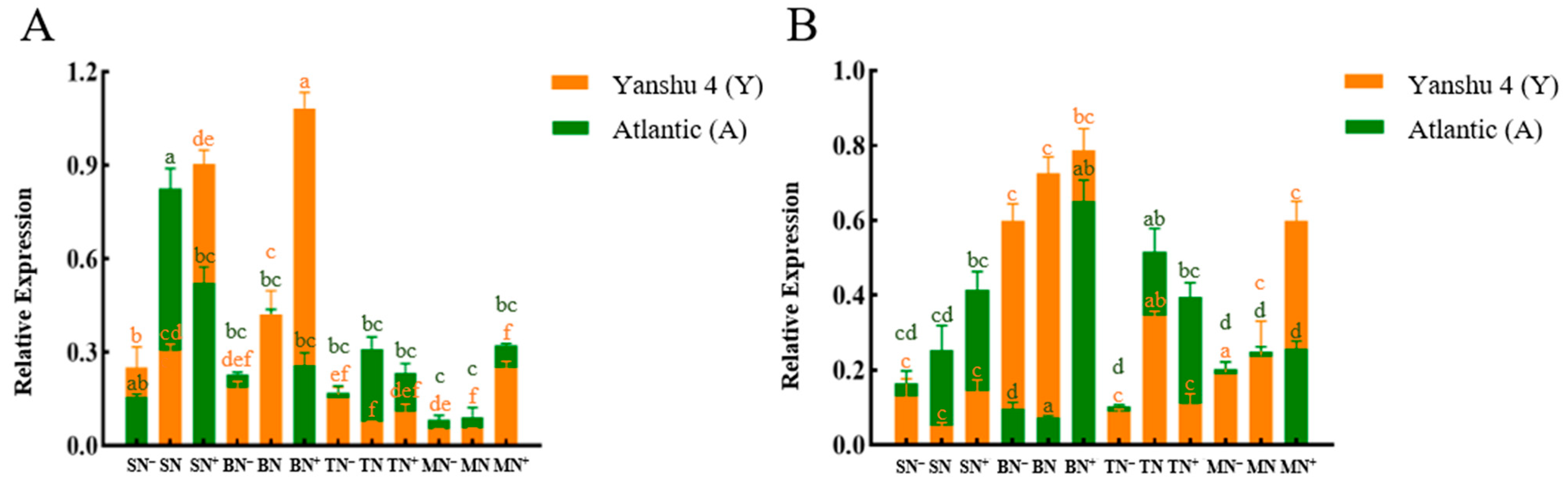
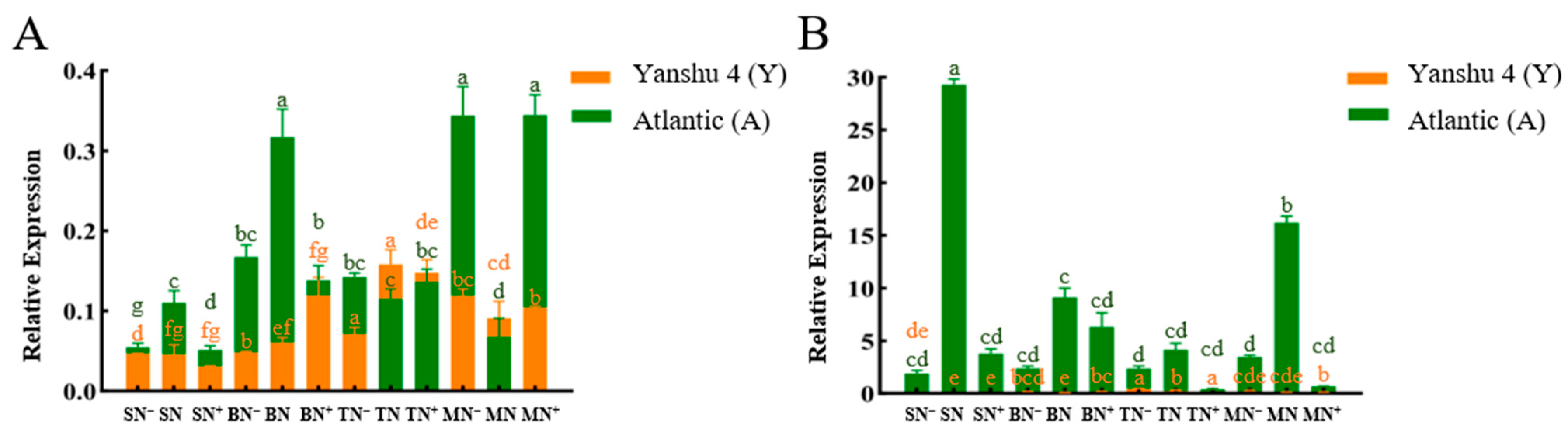
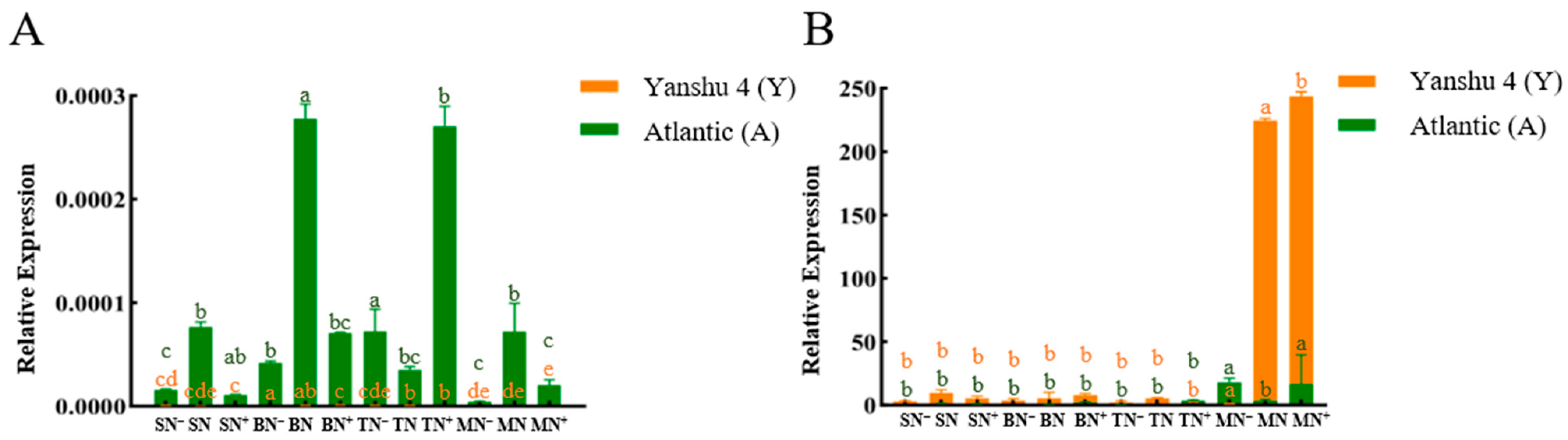
3.2.5. Analysis of Relative Expression of Glutamate Dehydrogenase Gene StGDH in Two Genotypes of Potato under Different N Supply Levels
3.2.6. Relative Expression Analysis of Nitrate Transporter StNRTs in Two Genotypes of Potato under Different N Supply Levels
3.3. Analysis of Key Enzyme Activity of N Metabolism in Leaves of Two Genotypes of Potato under Different N Supply Levels
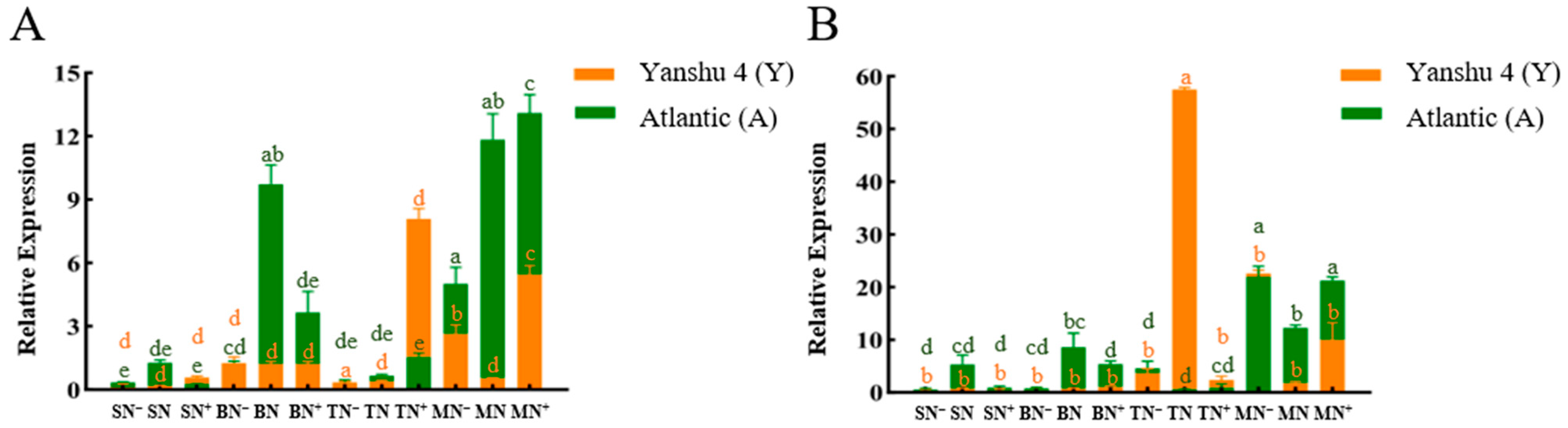
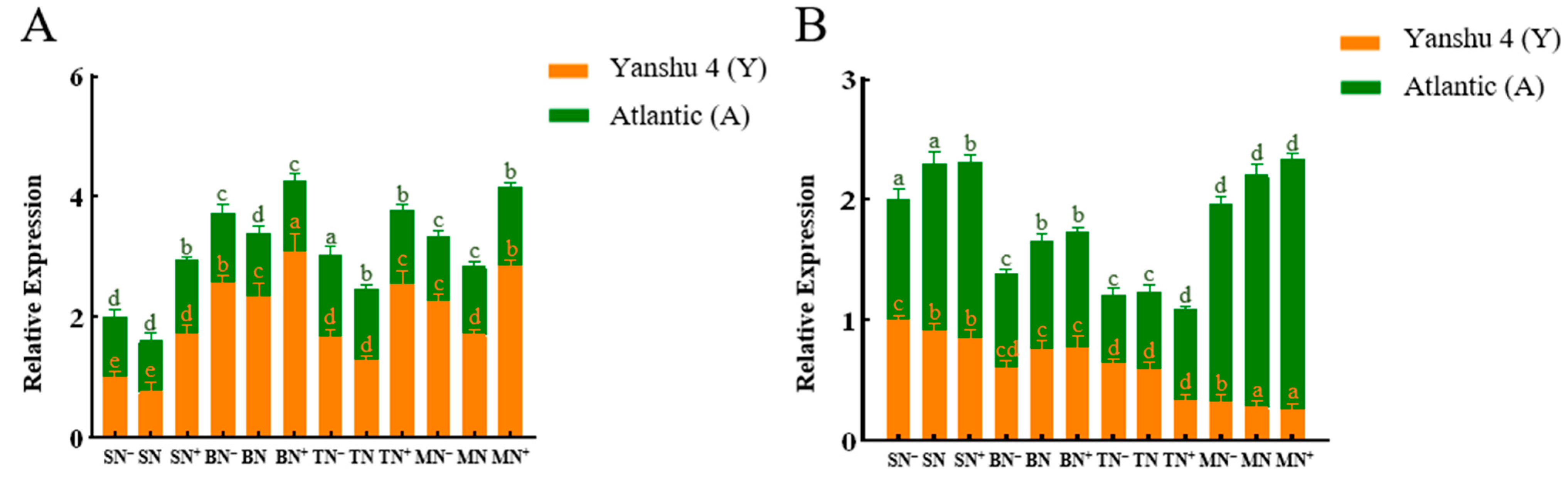
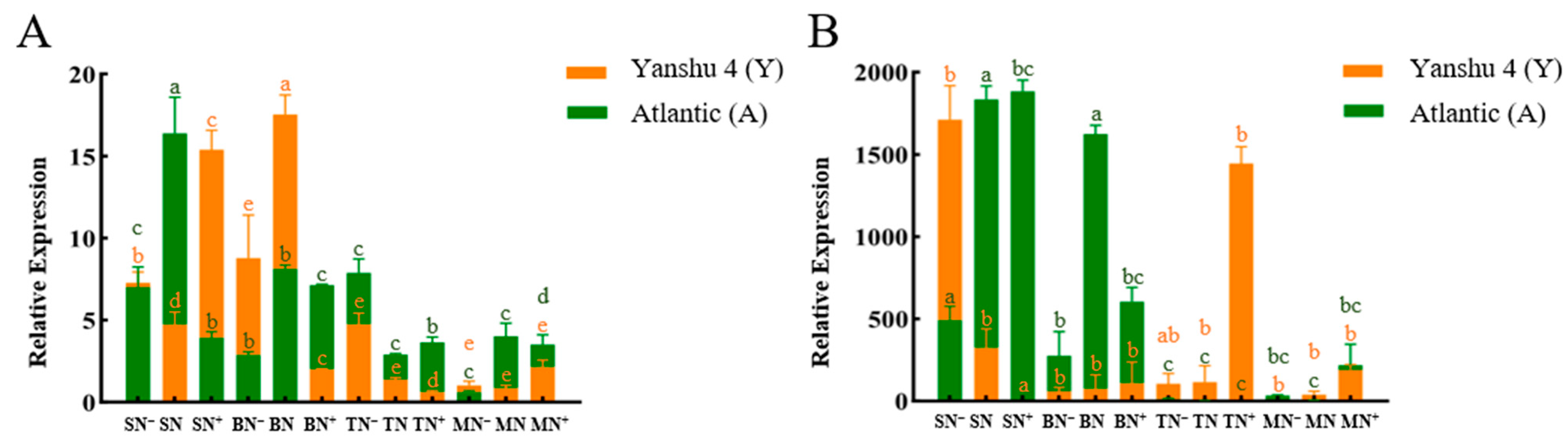
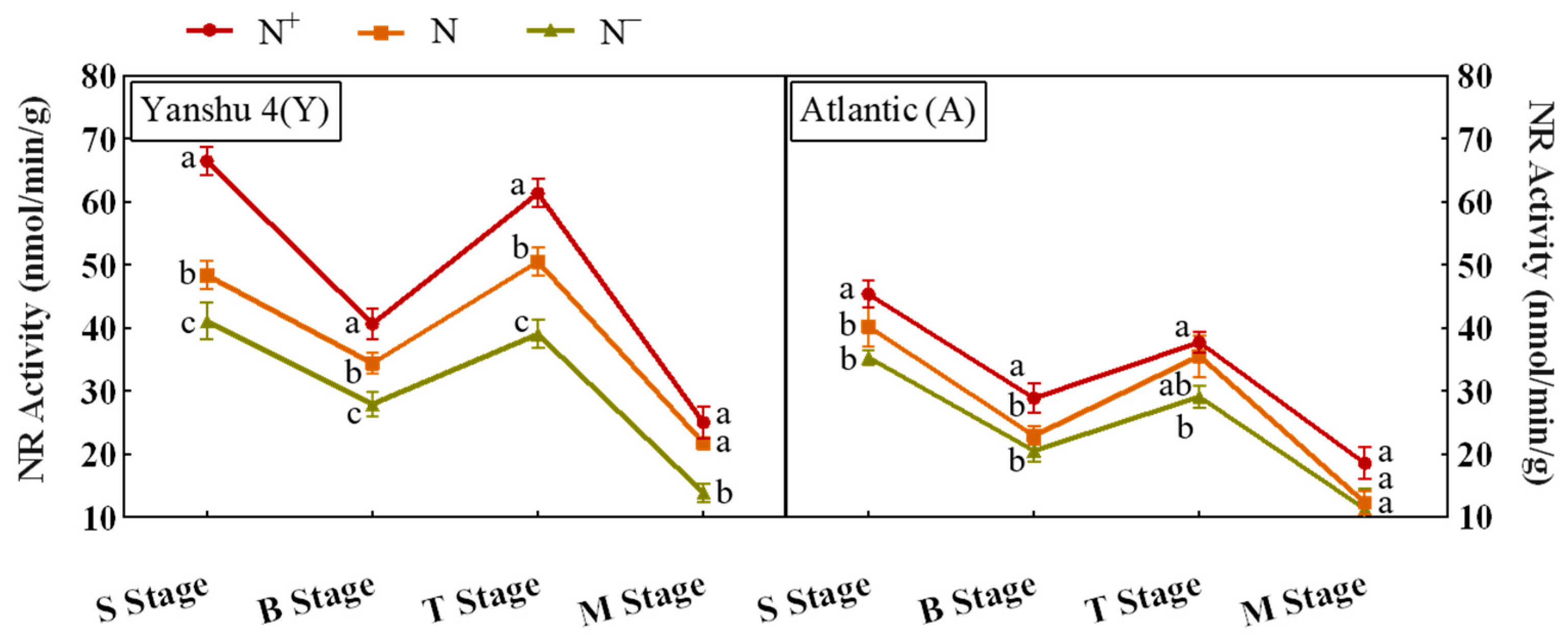
3.3.1. Analysis of Nitrate Reductase (NR) Activity in Leaves of Two Genotypes of Potato under Different N Supply Levels
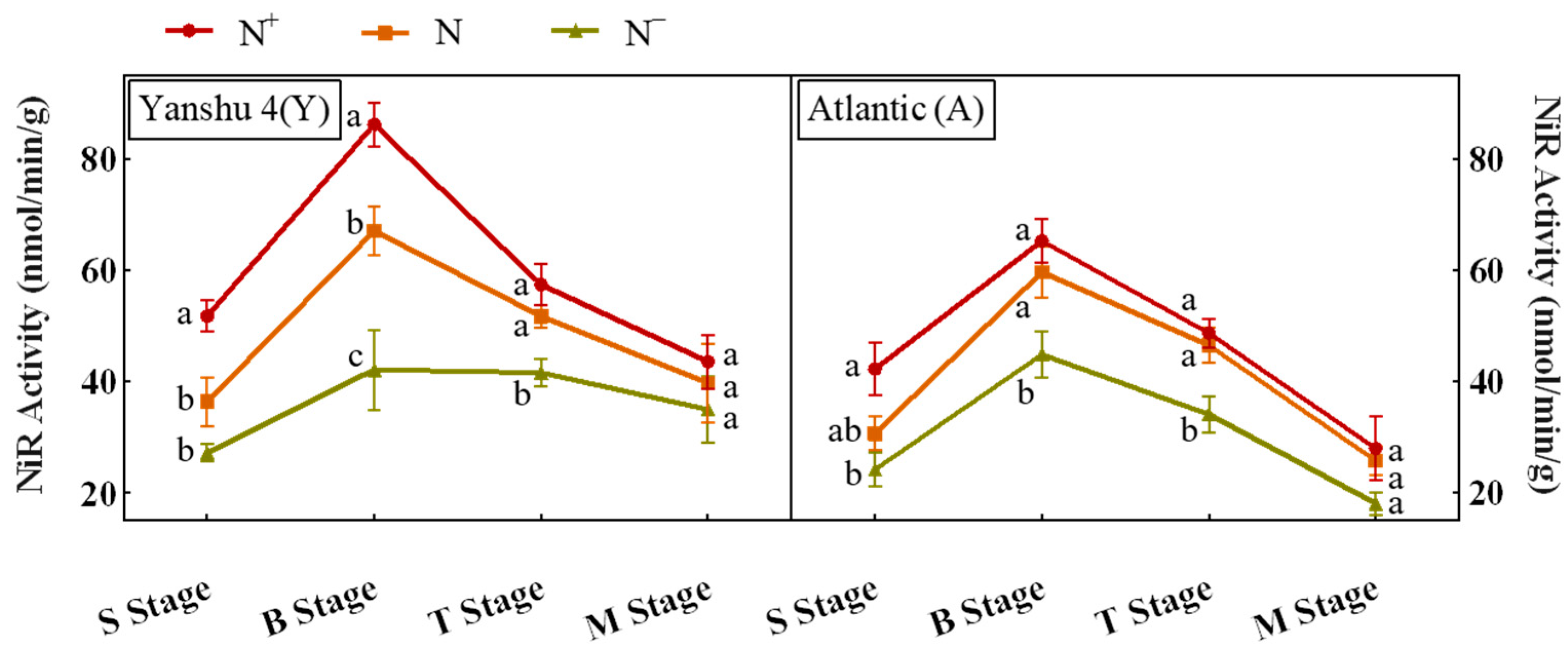
3.3.2. Analysis of Nitrite Reductase (NiR) Activity in Leaves of Two Genotypes of Potato under Different N Supply Levels
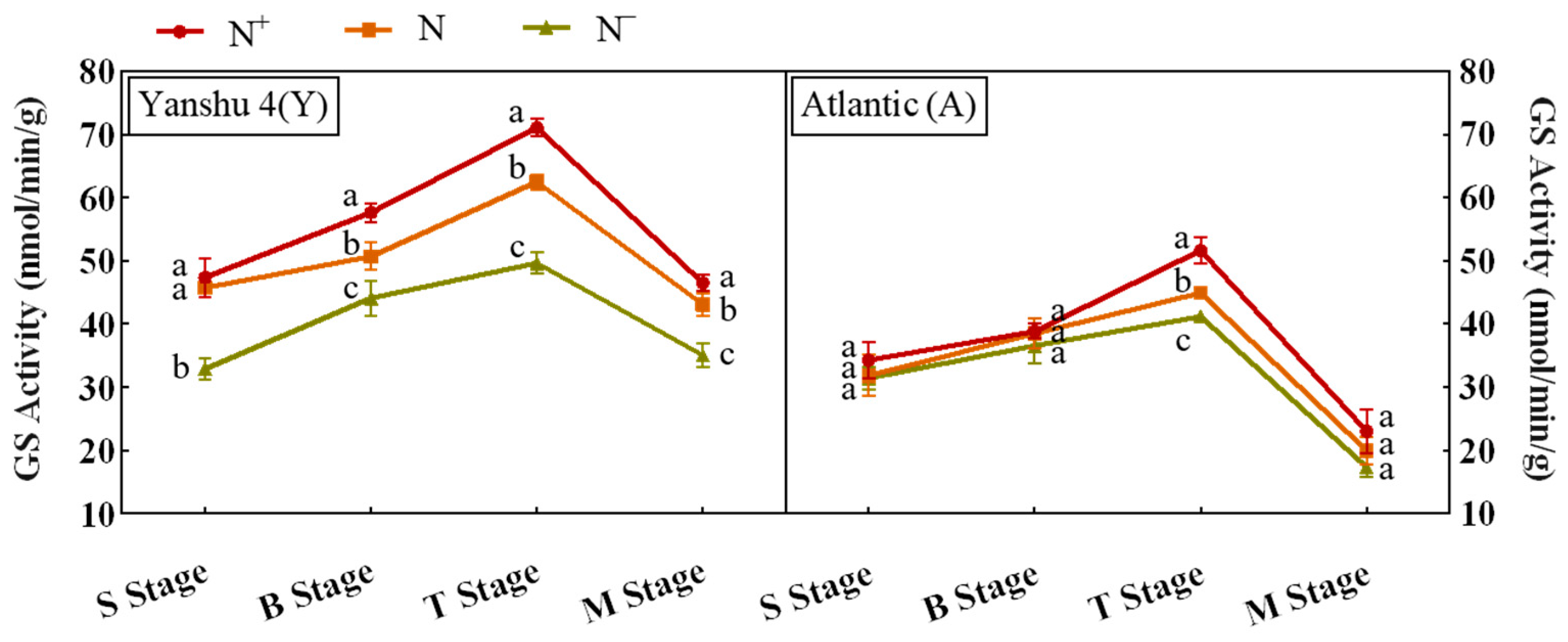
3.3.3. Analysis of Glutamine Synthase (GS) Activity in Leaves of Two Genotypes of Potato under Different N Supply Levels
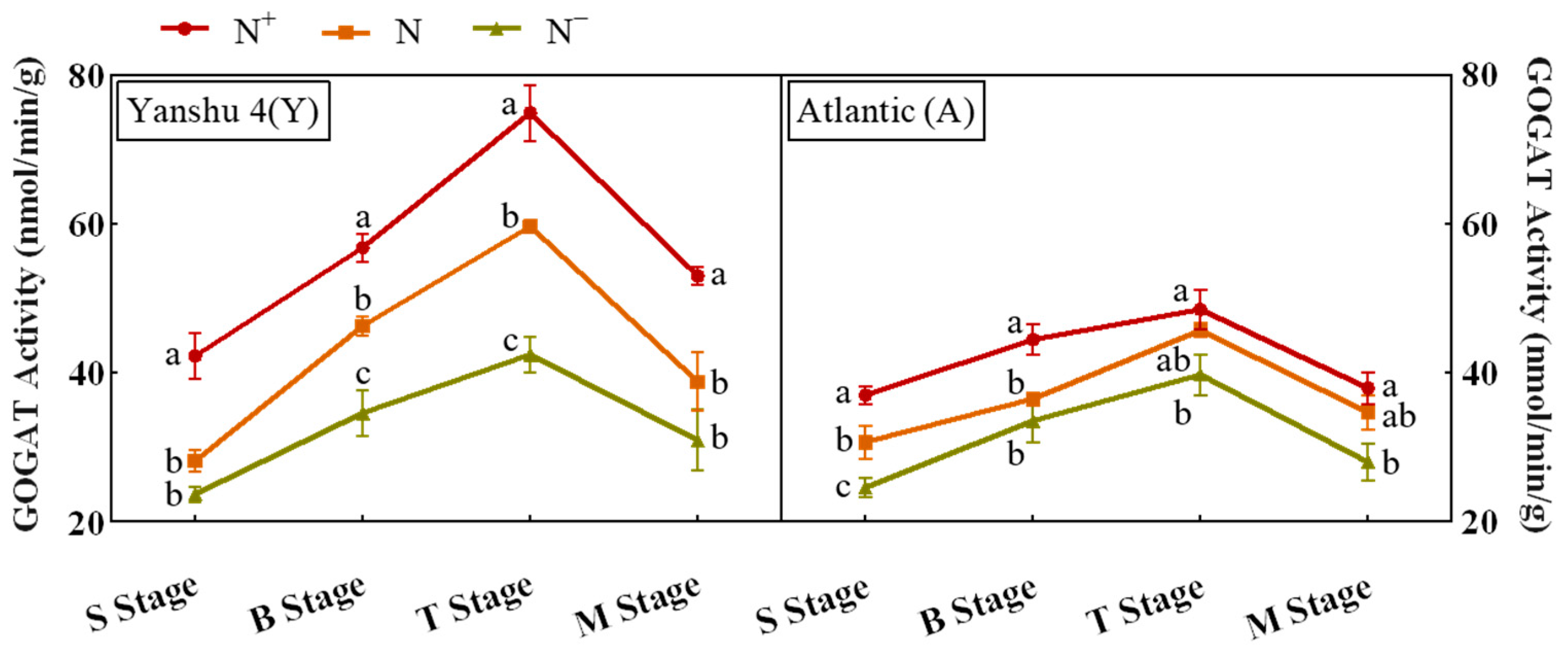
3.3.4. Activity Analysis of Glutamate Synthase (GOGAT) in Leaves of Two Genotypes of Potato under Different N Supply Levels
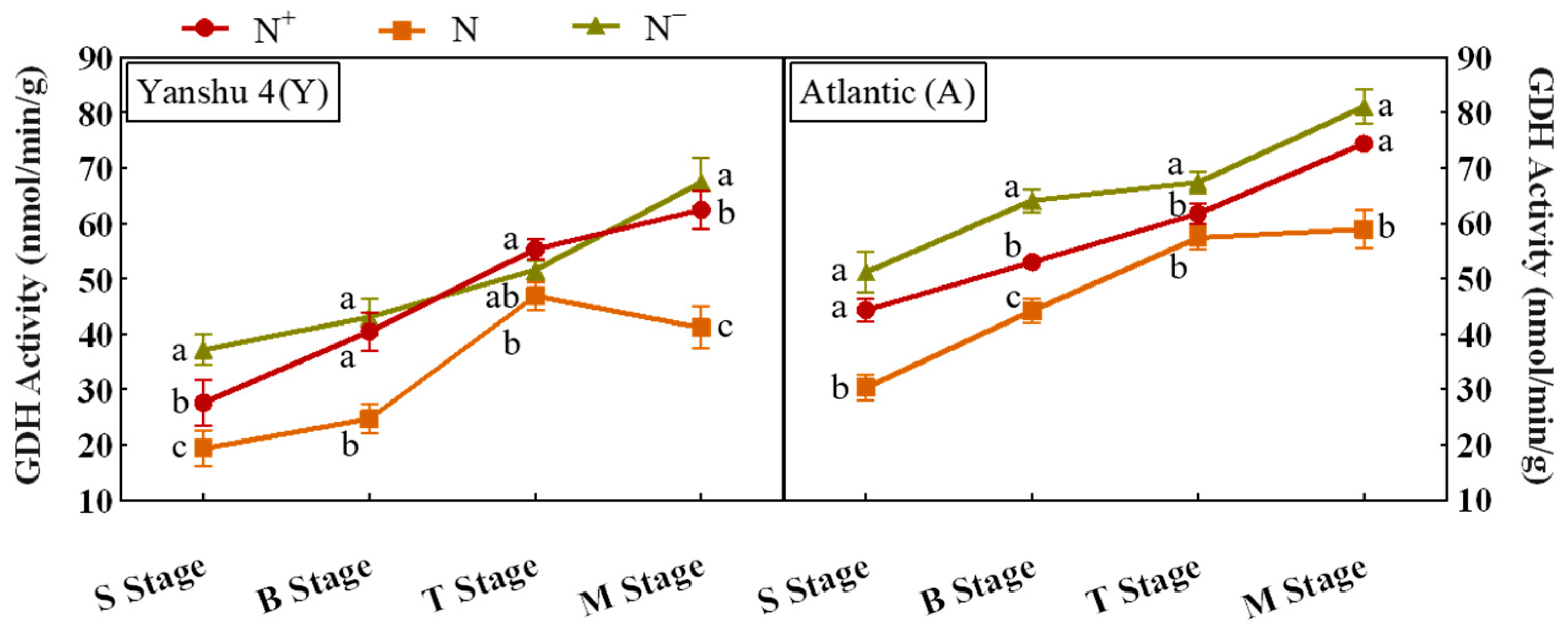
3.3.5. Analysis of Glutamate Dehydrogenase (GDH) Activity in Leaves of Two Genotypes of Potato under Different N Supply Levels
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Yang, Y.; Guo, Y.; Sun, J. The development status and future prospects of my country’s potato industry. J. Agric. Sci. Technol. 2017, 19, 29–36. [Google Scholar]
- Li, S.; Li, S.; Wang, Y.; Mou, B.; Zhao, Y.; Li, T.; Han, Y. Genetic diversity analysis of phenotypic traits of potato germplasm resources. Jilin Agric. Univ. 2020, 42, 269–279. [Google Scholar]
- Huang, F.; Zhang, L.; Li, X.; Cai, D. Current Situation and Countermeasures of China’s Potato Industry Development. Agric. Outlook 2017, 13, 25–31. [Google Scholar]
- Swaney, D.; Howarth, R.; Hong, B. Nitrogen use efficiency and crop production: Patterns of regional variation in the United States, 1987–2012. Sci. Total Environ. 2018, 635, 498–511. [Google Scholar] [PubMed]
- He, D. Genotypic Differences in Nitrogen Uptake and Use Efficiency in Potato. Master’s Thesis, Inner Mongolia Agricultural University, Hohhot, China, 2018. [Google Scholar]
- Jiao, J.; Wei, Q.; Pei, J.; Guo, Y.; Tang, L.; Shi, Y. Differences in response to nitrogen in different genotypes of potato. Acta Agric. Boreali-Sin. 2018, 33, 203–208. [Google Scholar]
- Wei, Q.; Cao, M.; Shi, Y.; Chen, B.; Zhang, L. Effects of nitrogen application levels on growth, development and quality traits of different potato varieties. Shenyang Agric. Univ. 2017, 48, 152–158. [Google Scholar]
- Williams, L.; Miller, A. Transporters responsible for the uptake and partitioning of nitrogenous solutes. Annu. Rev. Plant Phys. 2001, 52, 659–688. [Google Scholar]
- Zhang, L.; Deng, L.; Xu, N.; Pan, Z.; Li, Y.; Yang, Q. Differences in yield and nitrogen uptake and utilization of potato with different genotypes. Crop. Res. 2020, 34, 44–48. [Google Scholar]
- Zheng, S.; Li, G.; Yang, S.; Yuan, J.; Hao, K. Effects of nitrogen application amount and topdressing ratio on growth period and dry matter accumulation of winter potato. Sichuan Agric. Univ. 2009, 27, 270–274. [Google Scholar]
- Wei, D.; Wei, J.; Xiong, J.; Liang, H. Research progress on nitrogen nutrition in potato. Guangdong Agric. Sci. 2011, 38, 56–60. [Google Scholar]
- Huang, J.; Peng, Z.; Yu, J.; Wu, X.; Lin, Z.; Yang, L. Effects of Different Nitrogen Fertilizer Amounts on Yield, Quality and Nitrogen Utilization Efficiency of Winter Potato. J. Trop. Crops 2014, 35, 266–270. [Google Scholar]
- Sarkar, D.; Naik, P.S. Effect of inorganic nitrogen nutrition on cytokinein-induced potato microtuber production in vitro. Potato Res. 1998, 41, 211–217. [Google Scholar]
- Liu, H.; Zhao, Z. Research progress of plant nitrogen nutrition efficient genotype screening index. Anhui Agric. Sci. 2006, 34, 3265–3267. [Google Scholar]
- Masclaux-Daubresse, C.; Daniel-Vedele, F.; Dechorgnat, J.; Chardon, F.; Gaufichon, L.; Suzuki, A. Nitrogen uptake, assimilation and remobilization in plants: Challenges for sustainable and productive agriculture. Ann. Bot. 2010, 105, 1141–1157. [Google Scholar]
- McAllister, C.H.; Beatty, P.H.; Good, A.G. Engineering nitrogen use efficient crop plants: The current status. Plant Biotechnol. J. 2012, 10, 1011–1025. [Google Scholar] [PubMed]
- Liu, X.; Gong, Z. Research progress on nitrogen absorption and utilization by plants. Mod. Agric. 2012, 8, 20–21. [Google Scholar]
- Cheng, H. Screening of Potato Varieties with High Nitrogen Nutrient Utilization and Research on Their Physiological Mechanism. Master’s Thesis, Sichuan Agricultural University, Ya’an, China, 2010. [Google Scholar]
- Guo, Z.; Shen, A.; Kou, C.; Ma, Z.; Wang, S. The relationship between nitrate reductase activity and nitrogen efficiency after flowering in different wheat varieties. Chin. Agric. Sci. Bull. 2008, 5, 219–223. [Google Scholar]
- Liu, L.; Jing, W.; Han, Z.; Sun, X.; Lu, Y. Molecular analyses of tomato GS, GOGAT and GDH gene families and their response to abiotic stresses. Acta Physiol. Plant 2016, 38, 229. [Google Scholar]
- Masclaux-Daubresse, C.; Reisdorf-Cren, M.; Pageau, K.; Lelandais, M.; Grandjean, O.; Kronenberger, J.; Valadier, M.-H.; Feraud, M.; Jouglet, T.; Suzuki, A. Glutamine Synthetase-Glutamate Synthase Pathway and Glutamate Dehydrogenase Play Distinct Roles in the Sink-Source Nitrogen Cycle in Tobacco. Plant Physiol. 2006, 140, 444–456. [Google Scholar]
- Wang, J.; Han, J.; Yang, M.; Yao, D.; Zhou, Y.; Wang, W.; Wu, Z.; Yang, Q. Nitrogen uptake, transport and metabolism of different nitrogen-efficient maize varieties. J. Nucl. Agric. Sci. 2020, 34, 2800–2812. [Google Scholar]
- Yamaya, T. Genetic manipulation and quantitative-trait loci mapping for nitrogen recycling in rice. J. Exp. Bot. 2002, 53, 917–925. [Google Scholar] [PubMed] [Green Version]
- Chen, M.; Liu, C.; Liu, D.; Tang, Q.; Lin, J.; Liu, X. Heterologous expression of Alternaria brassica glutamate dehydrogenase gene AbGDH to improve nitrogen use efficiency in rice. Life Sci. Res. 2019, 23, 1–12. [Google Scholar]
- Zhang, X.; Cai, W.; Kan, H.; Xue, B.; Jia, Z.; Wang, S.; Wu, Y. Differential analysis of carbon and nitrogen metabolism and related gene expression in different varieties of flue-cured tobacco during aging. Chin. J. Tob. 2020, 26, 66–73. [Google Scholar]
- Jia, B.; Sun, Z.; Zhang, J.; Zhu, X. Nitrogen metabolism, key enzyme activity and related gene expression analysis of flue-cured tobacco during maturity. Henan Agric. Univ. 2020, 54, 559–565. [Google Scholar]
- Zhang, Y.; Zhao, Y.; Wang, Y.; Mou, B.; Zhang, J.; Ma, H.; Han, Y. Physiological and biochemical responses of potato Yanshu 4 to nitrogen and transcriptome analysis. Guangdong Agric. Sci. 2021, 48, 56–66. [Google Scholar]
- Gelli, M.; Duo, Y.; Konda, A.R. Identification of differentially expressed genes between sorghum genotypes with contrasting nitrogen stress tolerance by genome-wide transcriptional profiling. BMC Genom. 2014, 15, 179. [Google Scholar]
- Zhang, Y. The Regulation Mechanism of Nitrogen Application on Nitrogen Assimilation and Carbon Metabolism in Sugar Beet. Ph.D. Thesis, Northeast Agricultural University, Harbin, China, 2013. [Google Scholar]
- Huang, H.; Shen, Y.; Wang, W.; Lian, W.; Chen, X.; Zhai, X.; Yu, Q.; Yang, Z.; Jia, H. Cloning and expression analysis of tobacco nitrate transporter gene NtNRT2.4. Chin. J. Tob. 2016, 22, 84–91. [Google Scholar]
- Lezhneva, L.; Kiba, T.; Feria-Bourrellier, A.B. The Arabidopsis nitrate transporter NRT2.5 plays a role in nitrate acquisition and remobilization in nitrogen-starved plants. Plant J. 2014, 80, 230–241. [Google Scholar]
- Orsel, M.; Krapp, A.; Daniel-Vedele, F. Analysis of the NRT2 nitrate transporter family in Arabidopsis. Structure and gene expression. Plant Physiol. 2002, 129, 886–896. [Google Scholar]
- Yu, J.; Zhu, Z.; Zhang, Z.; Shu, J.; Yang, C.; Song, H.; Guan, C. Effects of glutamine synthase and glutamate synthase on nitrogen reuse in rapeseed under different nitrogen supply conditions. Crops J. 2014, 6, 81–85. [Google Scholar]
- Wang, X. Cloning and Functional Analysis of Key Genes in Response to Nitrate Nitrogen Signal in Fruit Tree Roots. Ph.D. Thesis, Shandong Agricultural University, Tai’an, China, 2012. [Google Scholar]
- Sun, M. Nitrogen Utilization Evaluation of Introgressed Tomato Strains of Pannari and Preliminary Research on the Mechanism of Action of High-Efficiency Strains. Master’s Thesis, Shenyang Agricultural University, Shenyang, China, 2018. [Google Scholar]
- Wu, Y.; Li, Q.; Dou, P.; Ma, X.; Yu, D.; Luo, Y.; Kong, F.; Yuan, J. Effects of nitrogen fertilizer on chlorophyll content and nitrogen-metabolizing enzyme activity of different low-nitrogen-tolerant maize varieties in late growth stage. J. Herbol. 2017, 26, 188–197. [Google Scholar]
- Ma, D.; Wang, Y.; Zhao, C.; Lin, Q. Effects of nitrogen application rate and post-flowering soil water content on nitrogen metabolism characteristics and grain protein content of wheat. J. Plant Nutr. Fertil. 2008, 6, 1035–1041. [Google Scholar]
- Shah, J.M.; Bukhari, S.A.H.; Zeng, J.B. Nitrogen (N) metabolism related enzyme activities, cell ultrastructure and nutrient contents as affected by N level and barley genotype. J. Integr. Agric. 2017, 16, 190–198. [Google Scholar]
- Zhang, T. Physiological Response and Differential Gene Expression of Efficient Nitrogen Utilization in Potato. Ph.D. Thesis, Inner Mongolia Agricultural University, Hohhot, China, 2020. [Google Scholar]
- Duan, Y.; Zhang, Y.; Wang, S.; Shen, Q. Response of rice with different nitrogen efficiencies to nitrification and its physiological mechanism during the whole growth period. Chin. J. Ecol. 2007, 3, 1086–1092. [Google Scholar]




















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer Name | Primer Sequence |
|---|---|
| Reference-StEF1α F | GATGGTCAGACCCGTGAACA |
| Reference-StEF1α R | CCTTGGAGTACTTCGGGGTG |
| StNADH-GOGAT F | GTGGTGGTGTTGCCTATGTTCTTGA |
| StNADH-GOGAT R | TTGTGTTACGCTGGTGTTGCTGTAT |
| StFd-GOGAT F | CAAGCGGATTCGTAGAAGGATTGGT |
| StFd-GOGAT R | CGTTGAAGGTCAAGAACAGCATTGC |
| StGS1 F | AATGAGCGTCGTCTCACTGG |
| StGS1 R | GGCTGGCCTGTCCTCAAAG |
| StGS1-2 F | GGAAGTGATGCCTGGACAGTGG |
| StGS1-2 R | CAGCAACCTCAGCAATCCTCTCG |
| StGS1-3 F | TGGACAAGCACCTGGAGAAGACA |
| StGS1-3 R | CCAAGATGTTGTTACCACCACGGAA |
| StGS1-4 F | CTGGAAAGGCTTTTGGACGC |
| StGS1-4 R | CCAGGCATCACTTCTCCGTT |
| StGS2 F | TGCTGCTTATGGGGTTGGC |
| StGS2 R | CTGTGTGTCCCTTCCGATCCT |
| StNR F | ACGCTGAACTTGCTAACGCTGAA |
| StNR R | ATGGCACGGAGTTGTAATGACAGAG |
| StNiR F | TGTGCTACCTGATGTGCCTGAGA |
| StNiR R | TCCTGACATTATCCATGCCACTCTG |
| StGDH F | TCGGCGGTGGTGATGTAATGAGTA |
| StGDH R | CTCCTCCCAGAGCACAGGGTATAAG |
| StNRT1.5 F | CGACGATGACACAACTGGTGGTAG |
| StNRT1.5 R | CCTGCTTCTGGCTCTCAAGTTCC |
| StNRT2.4 F | GCAGACGGTGGCTGGATTGATG |
| StNRT2.4 R | AAGCACGCCATAACCAGAACAGAC |
| StNRT2.5 F | ACCTGCTCTGTTTCAGGCTT |
| StNRT2.5 R | GTTTCGCGTAATCTCCATCGG |
| StNRT2.7 F | GCAGACGGTGGCTGGATTGATG |
| StNRT2.7 R | AAGCACGCCATAACCAGAACAGAC |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Han, Z.; Lu, Y.; Zhao, Y.; Wang, Y.; Han, Z.; Han, Y.; Zhang, J. Analysis of Relative Expression of Key Enzyme Genes and Enzyme Activity in Nitrogen Metabolic Pathway of Two Genotypes of Potato (Solanum tuberosum L.) under Different Nitrogen Supply Levels. Horticulturae 2022, 8, 769. https://doi.org/10.3390/horticulturae8090769
Han Z, Lu Y, Zhao Y, Wang Y, Han Z, Han Y, Zhang J. Analysis of Relative Expression of Key Enzyme Genes and Enzyme Activity in Nitrogen Metabolic Pathway of Two Genotypes of Potato (Solanum tuberosum L.) under Different Nitrogen Supply Levels. Horticulturae. 2022; 8(9):769. https://doi.org/10.3390/horticulturae8090769
Chicago/Turabian StyleHan, Zhijun, Yue Lu, Yanfei Zhao, Yaping Wang, Zhongcai Han, Yuzhu Han, and Jingying Zhang. 2022. "Analysis of Relative Expression of Key Enzyme Genes and Enzyme Activity in Nitrogen Metabolic Pathway of Two Genotypes of Potato (Solanum tuberosum L.) under Different Nitrogen Supply Levels" Horticulturae 8, no. 9: 769. https://doi.org/10.3390/horticulturae8090769
APA StyleHan, Z., Lu, Y., Zhao, Y., Wang, Y., Han, Z., Han, Y., & Zhang, J. (2022). Analysis of Relative Expression of Key Enzyme Genes and Enzyme Activity in Nitrogen Metabolic Pathway of Two Genotypes of Potato (Solanum tuberosum L.) under Different Nitrogen Supply Levels. Horticulturae, 8(9), 769. https://doi.org/10.3390/horticulturae8090769