1. Introduction
Pyrogenic dynamics are an integral element of the functioning of treeless arid ecosystems in southern Russia. Steppe fires are characterized by lower fire intensities than crown fires in boreal forests. Probably for this reason, pyrogenic successions in treeless arid ecosystems have been poorly studied [
1]. In the Republic of Kalmykia, the territory most affected by fires is the State’s Natural Biosphere Reserve “Chernye Zemli” [
1], where the largest fires among all specially protected natural areas in the southeast of the European part of Russia were recorded. Over the period 1987–2022, 88% of the area of the “Chernye Zemli” Reserve and adjacent territories has been burned 4–7 times, which is significantly more than in Central Kazakhstan [
2]. Up to 90% of fires from the five-kilometer zone adjacent to the “Chernye Zemli” Reserve, and all fires from its protective zone, spread to the reserve core. However, since 2018, the number of fires on the territory of the “Chernye Zemli” Nature Reserve has decreased to almost zero [
3].
Periodic exposure to certain environmental factors can considerably change ecosystems. This is especially true for fires in arid ecosystems in recent decades [
4,
5,
6]. Burning of sagebrush communities significantly reduces the cover of dwarf shrubs and (sub)shrubs, that die due to the elimination of renewal buds located above the soil [
7,
8,
9]. As a result, in the first few years after fires, previously dominant (sub)shrubs are found only as single individuals that survived the fire. On the other hand, the projective cover of cespitose grasses increases after fires. In the Chernye Zemli, these are primarily small-turf
Poa bulbosa L. and large-turf species of the genera
Stipa and
Agropyron. Thus, pyrogenic communities are primarily characterized by the predominance of cespitose grasses and a small participation of (sub)shrubs. During further succession, the cover of (sub)shrubs is restored via seed propagation from soil seed bank and intake from outer territories, primarily under the strong influence of ungulates, which primarily eat grasses and contribute to the restoration of sagebrush community [
9,
10]. However, excessive grazing pressure prevents the restoration of (sub)shrubs and leads to the formation of communities with a predominance of
Poa bulbosa and annual species [
11]. There is an opinion that the predominance of (sub)shrub vegetation on the territory of Kalmykia is the result of many years of overgrazing, and steppe phytocenoses with a predominance of
Stipa and
Agropyron should be considered as climax communities [
12].
Despite the fact that the structure and dynamics of desert communities in the “Chernye Zemli” Nature Reserve have been studied previously [
13,
14,
15], there are many aspects that have not been well studied, including pyrogenic succession changes in the composition of plant communities in Kalmykia. There is a lack of understanding of the effects of frequent burning of the same area on vegetation composition. The reason is that only recently, thanks to the gradual accumulation of a homogeneous array of Landsat images covering the entire territory of the Earth over several decades, has it become possible to accurately determine the frequency of burning and the age of the last fire. Free and easy access to Landsat images has become available in 2008 (
https://www.usgs.gov/news/free-open-landsat-data-unleashed-power-remote-sensing-decade-ago) (accessed on 23 January 2024).
The purpose of our work was to evaluate the effects of time since the last fire and fire frequency on phytocenoses at the “Chernye Zemli” Reserve and adjacent areas. The study investigated three issues: (1) assessing the degree of differences in the composition of communities in and beyond the “Chernye Zemli” Nature Reserve, (2) identifying changes in plant communities during pyrogenic succession, and (3) estimating the impact of fire frequency on the state of plant communities.
3. Results
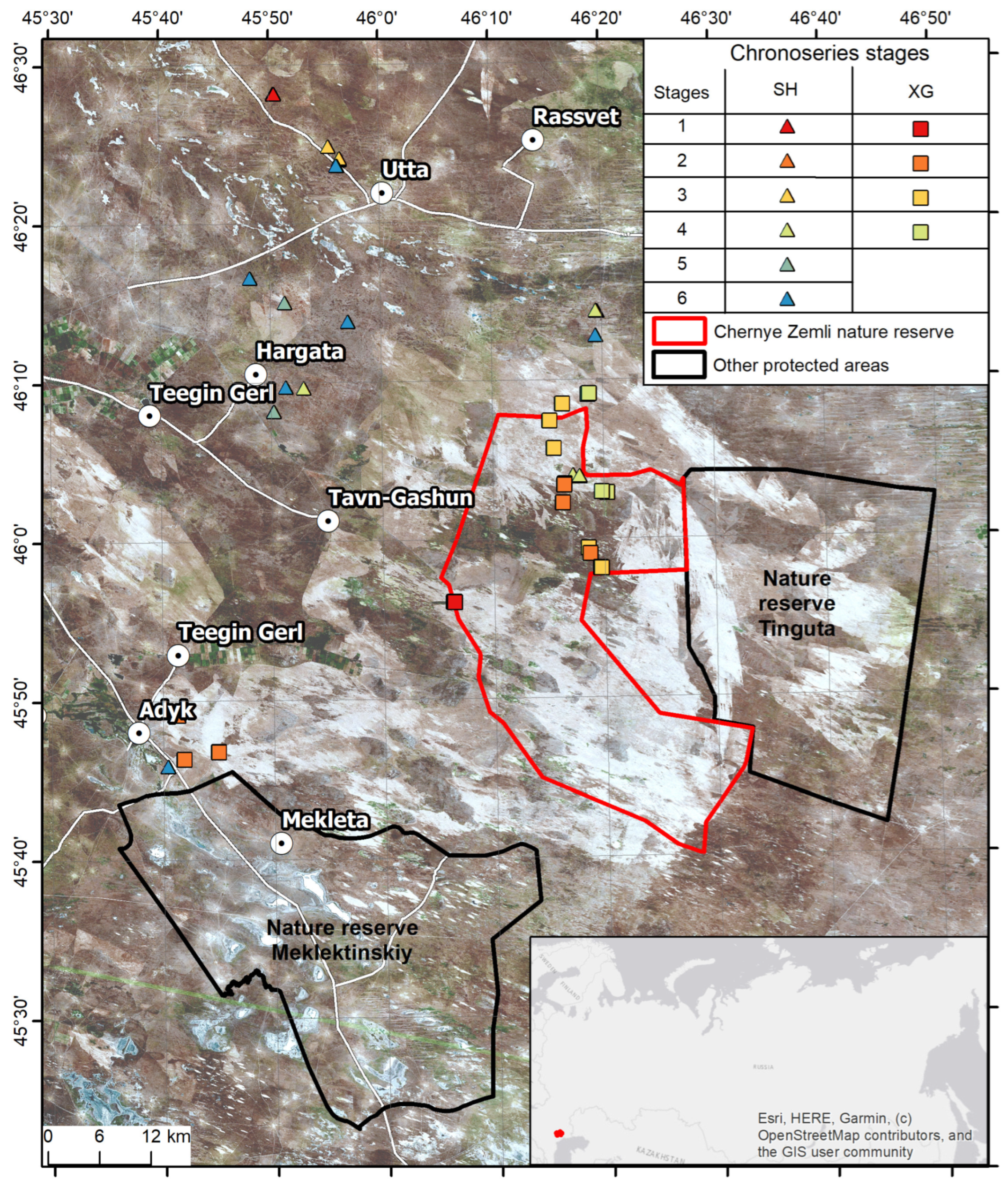
For 1987–2022, the territory under consideration (
Figure 1) was exposed to fire 20 times. The first major fire was recorded in 1991 (0.04% of the territory), and the last was recorded in 2015 (48%). There were no large fires in 1986–1990, 1996–1997, 2009, 2012 and 2016–2022. In 2006 and 2002, very large fires affected more than 96% and 57% of the territory, respectively. In 1998, 2001, 2004, 2007, 2011, 2014, 2015, fire affected about 30% of the area of the “Chernye Zemli” Nature Reserve. In other years, the percentage of the burned area varied from 0.04 to 18%. From 1986 to 2020, the maximum number of fires was 10, and such territories accounted for 0.0001% of the total area of the “Chernye Zemli” Reserve.
According to remote sensing data, all the territory of “Chernye Zemli” Reserve has been burnt at least once in the last 15 years. Therefore, the vegetation cover of the reserve is a mosaic of areas belonging to initial or middle chronological stages. For this reason later stages of pyrogenic succession and communities that did not burn out during the period of availability of Landsat images (since 1984) were examined only outside the reserve, in the area of the Utta and Adyk villages (see
Figure 1).
3.1. Comparative Analysis of Plant Communities in the “Chernye Zemli” Reserve and Beyond
Between the communities inside the reserve and outside of it with sagebrush projective cover <1%, the average number of species per 10 × 10 m plot did not significantly differ (
p = 0.8;
Table 2) in the reserve and in the pastures: 11 and 12 species, respectively. However, communities in the protected area and outside it differed significantly in projective cover of two species: the cover of
Stipa sareptana was significantly higher in the reserve, and
Prangos odontalgica (Pall.) Herrnst. And Heyn outside its border (
Table 3). The average number of species per plot with sagebrush projective cover from 1 to 10% also did not differ in the reserve and in adjacent pastures (17 and 14 species respectively;
p = 0.2) in the absence of species with significantly different projective cover in the reserve and outside it. The results obtained allow us to consider sites in the reserve and outside of it as a single data set.
3.2. Comparative Analysis of Plant Communities of the “Chernye Zemli” at Different Stages of Pyrogenic Succession
In the first few years of succession, (sub)shrubs are almost completely absent in pyrogenic communities, and sedges (
Carex stenophylla) and cespitose grasses (large-turf
Stipa sareptana, and less often,
S. lessingiana and small-turf
Poa bulbosa;
Table 1) predominate. Annuals are numerous, among which the most common are
Ranunculus testiculatus, Alyssum desertorum, Veronica verna or
V. triphyllos, Bromus tectorum, and
Ceratocarpus arenarius. Among the ephemeroids, the most typical are
Ranunculus oxyspermus, and the less common are tulips (
Tulipa suaveolens Roth,
Tulipa sylvestris subsp. australis,
T. biflora) and
Prangos odontalgica. Species richness in the first few years after a fire can vary from 5 to 10 or more species per 100 m
2 and the total projective cover is usually below 10–15%. Species richness is the highest in areas that burned 5–10 years ago, where it varies from 7 to 20 or more species per 100 m
2.
With age, shrubby species begin to appear in some burnt areas, e.g., different sagebrushes (Artemisia lercheana, A. santonicum, A. pauciflora Weber ex Stechmann, etc.), Bassia prostrata, Krascheninnikovia ceratoides, and in saline areas—Anabasis aphylla. Most often, (sub)shrubs grow in communities starting from stage 3 (9–13 years after the fire). But in a number of cases, communities with considerable participation of sagebrush are also characteristic of the second stage. On heavily overgrazed pastures near the reserve and along its periphery at stages 3 and 4, on the contrary, (sub)shrubs do not play an important role in the communities to the point of disappearance, and Poa bulbosa prevails in the plant cover.
Communities of late chronological stages outside the boundaries of the “Chernye Zemli” Nature Reserve are characterized by varying participation of (sub)shrubs, which can have a projective cover of 1–5% and never act as dominants, giving way to small-turf species. In unburned areas outside the “Chernye Zemli” Nature Reserve, the cover of (sub)shrubs can range from 1 to 40% or more, which may be due to edaphic habitat conditions and varying intensity of grazing. At the same time, small-turf P. bulbosa and, less commonly, C. stenophylla also have high coverage, taking the position of a dominant or subdominant in the community.
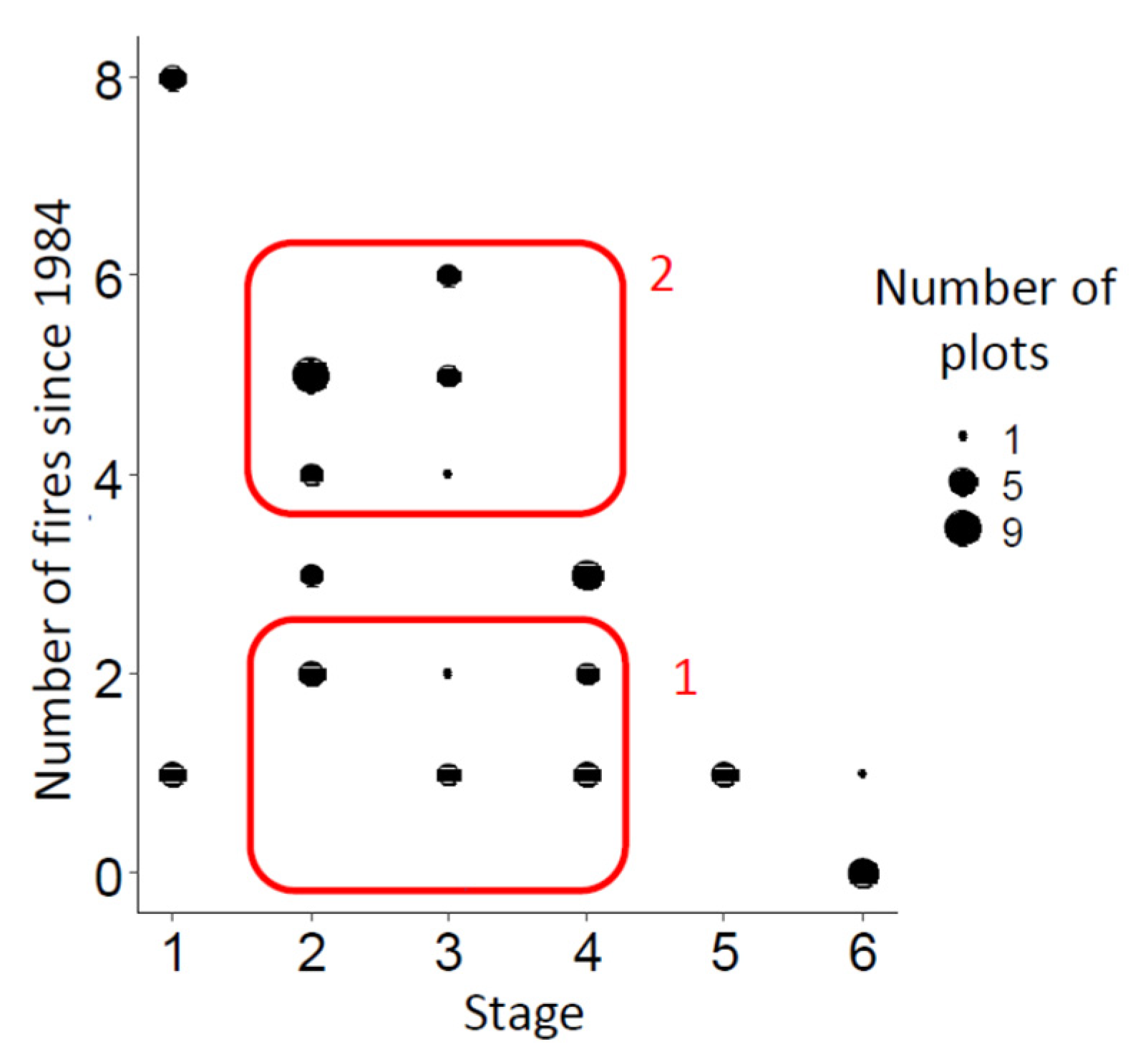
To assess changes in plant communities over time after a fire, for each pair of chronological stages we compared the mean projective cover of species common to them, combining data on plots from the xerophytic-grass and (sub)shrub chronoseries to increase the sample size. The correlation of the average projective cover of common species was most pronounced between successive stages, especially in pairs of early stages 1–2 and 2–3. This indicates a gradual change in the community for at least 10 years after the fire (
Table 4).
To determine whether the pre-fire cover of a species influences its likelihood to survive or regenerate after a fire, we compared the pre-fire projective cover of species that were detected and undetected in the early post-fire stage (stage 1). For each site from stage 1, the pre-fire stage was determined according to the age of the previous fire (see
Section 2.4). Of the 30 species found in the pre-fire stage of the xerophytic-grass chronoserie, seven species (
Carex stenophylla, Ranunculus falcatus, Poa bulbosa, Prangos odontalgica, Ranunculus oxyspermus, Stipa sareptana, Veronica triphyllos) were also found in the post-fire stage. In the (sub)shrub chronoserie, out of 33 species found in the pre-fire stage, 12 (
Alyssum desertorum, Artemisia lercheana, Carex stenophylla, Ranunculus testiculatus, Ranunculus falcatusr, Elymus repens, Lepidium perfoliatum, Poa bulbosa, Prangos odontalgica, Ranunculus oxyspermus, Tanacetum achilleifolium, Tulipa biflora) were also present after the fire. It is interesting that in the pre-fire stages of both chronoseries, three species (five in total) from the order Caryophyllales were found:
Amaranthus sp.,
Anabasis aphylla, Bassia prostrata (family Amaranthaceae),
Gypsophila paniculata,
Holosteum umbellatum (family Caryophyllaceae). But in the first year after the fire, not a single species of this order was discovered. In both chronoseries examined, species found in both pre-fire and post-fire stages had greater average projective cover in the pre-fire stage than species found only in the pre-fire stage (see
Section 2.4,
Table 5).
3.3. Assessment of the Impact of Fire Frequency on Plant Communities of the “Chernye Zemli” Nature Reserve
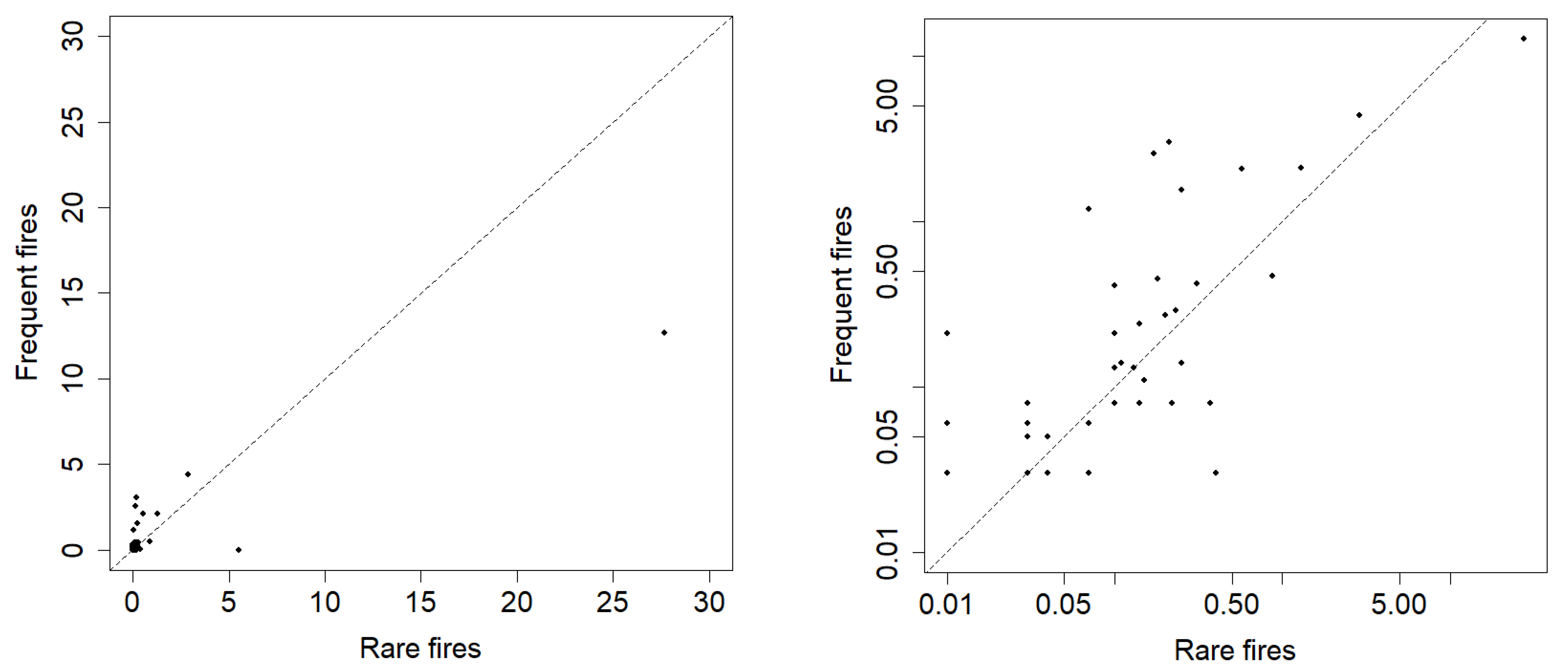
The majority of species found in one group according to fire frequency were also found in the other (79% of species in the group with rare fires and 76% of species in the group with frequent fires). Along with a significant correlation of the average projective cover of these species in two groups (
r = 0.64,
p = 0.00002,
Figure 3, right), this indicates the absence of significant differences in the species representation in areas with different frequencies of pyrogenic impact. When species that were found only in areas with rare or in areas with frequent fires were included in the analysis, correlation was similar (
r = 0.54,
p = 0.000004,
Figure 3, left).
In each of the two groups, four indicator species were identified. In rarely burned areas, these were
Lepidium perfoliatum (
p = 0.021),
Artemisia lercheana (
p = 0.008),
Chorispora tenella (Pall.) DC (
p = 0.030),
Tanacetum achilleifolium (
p = 0.025). In frequently burned ones, these were
Stipa sareptana (
p = 0.002),
Medicago orthoceras (
p = 0.013),
Crepis sancta (
p = 0.019), and
Filago arvensis L. (
p = 0.010). In the two groups, projective covers differed most noticeably in
Artemisia lercheana and two cespitose grasses—
Poa bulbosa and
Stipa sareptana (
Table 6).
3.4. Non-Metrical Multidimensional Scaling
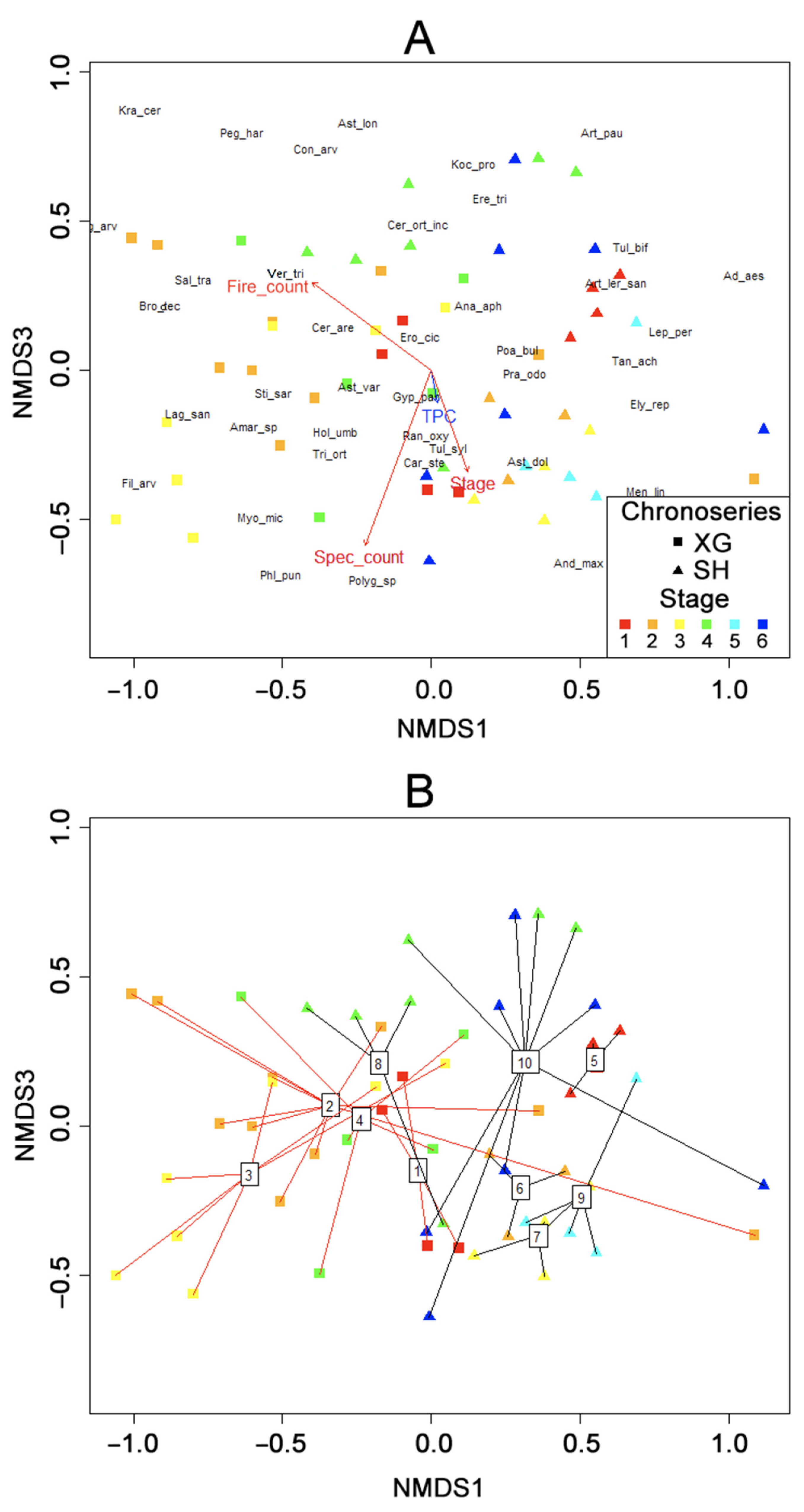
The results of NMDS (
Figure 4A) show a fairly uniform transition of communities from one chronoserie to another among the collected data array. At the same time, communities assigned to the (sub)shrub chronoserie are mainly presented on the right side of the diagram, and communities of the xerophytic-grass chronoserie are presented on the left. The centroids of groups of geobotanical descriptions (
Figure 4B), corresponding to the stages of different chronoseries, are also grouped in the same way. At the same time, the centroids of groups 4 and 8, corresponding to the fourth stages of the xerophytic-grass and (sub)shrub chronoseries, respectively, are located close together. This reflects the smooth transition of communities from the xerophytic-grass to subshrub chronoserie under the influence of grazing at this stage.
Communities of the xerophytic-grass chronoserie are mostly located at the end of the vector of fire frequency, whereas communities of the subshrub chronoserie are mostly located in the opposite direction. It reflects the generally low resistance of subshrubs to fire. At the same time, resistance to frequent fires apparently varies also among annuals. An increase in the frequency of pyrogenic impact is associated with an increase in the contribution of such annual species as Buglossoides arvensis (Bug_arv), Bromus tectorum (Bro_tec), Salsola tragus (Sal_tra), Veronica triphyllos (Ver_tri). Communities with a low frequency of fires are characterized by an increase in the contribution of Meniocus linifolius (Men_lin), Veronica verna (Ver_ver, in the diagram under the legend), and Androsace maxima (And_max).
4. Discussion
Pyrogenic dynamics of treeless communities are studied worldwide, e.g., sagebrush communities of Great Basin in North America [
30,
31,
32], Mediterranean heathlands [
33], meadows in Scandinavia [
34,
35,
36], etc. However, such studies are far fewer than studies of pyrogenic succession in forests [
37,
38]. Central Asia is especially understudied in this aspect due to social-economic problems after the USSR breakup in the early 1990s [
6].
An unambiguous interpretation of the results of assessing the impact of fires on the vegetation of the “Chernye Zemli” Nature Reserve and its vicinity is impossible due to the mosaic nature of the vegetation cover and the influence of grazing. Nevertheless, in this work a number of patterns have been discovered that make it possible to improve the understanding of pyrogenic successions in desert plant communities of the region.
The projective cover of most plant species did not differ between the protected area and beyond. Based on the data presented, we can conclude that the protective status of the “Chernye Zemli” Reserve does not affect the species richness of plant communities and has virtually no effect on the plant species representation. The lack of influence of territory conservation on species richness does not correspond to the known data on the consequences of excluding grazing in arid communities, where long-term absence of cattle reduces the species richness of phytocenoses (e.g., [
39,
40,
41]). We can assume that this is probably due to the absence of fences along the perimeter of the reserve, which allows the livestock grazing near the borders to invade its territory, increasing the grazing load. In addition, one can assume a considerable impact from wild ungulates (
Saiga tatarica L., 1766) on the territory of the reserve, especially in large calving aggregations in the spring, numbering a total of 4000–6000 individuals [
42]. Thus, even within the “Chernye Zemli” Reserve, a noticeable influence of large herbivores on the course of pyrogenic succession cannot be ruled out. Meanwhile, some studies provide information about the low impact of cattle on species richness for vegetation of some biomes, e.g., sagebrush communities in Wyoming, USA [
43,
44].
The correlation of the mean projective cover of species at different age stages of succession was calculated for a combined sample of xerophytic-grass and (sub)shrub chronoseries, since communities with a predominance of (sub)shrubs can be considered as the indigenous vegetation of the region under the conditions of previous land use [
15,
19,
45]. The smooth transition of communities from the xerophytic-grass to subshrub is confirmed by our NMDS analysis (
Figure 4B). The degree of participation of (sub)shrubs in pyrogenic communities depends on the parameters of fires and the intensity of grazing. At the same time, communities of the (sub)shrub chronoserie, despite the presence of sagebrush, at the early stages of pyrogenic succession have obvious features of xerophytic-grass communities. The most pronounced correlation of the average projective cover of species was observed between the early chronological stages of pyrogenic succession. Projective cover of individual species at chronological stages 1–2 and 2–3 significantly correlates with a coefficient of
r > 0.7. At the same time, for the pair of stages 1–3 the correlation is insignificant,
r < 0.5. Based on this, it can be assumed that during the first ten years after a fire, the role of species in the community changes gradually. For the three subsequent stages, the correlation is significant for all pairs of stages (4–5, 5–6 and 4–6), but modest with
r = 0.5–0.6. Based on this, we hypothesize that 10 years after a fire, the role of fire age in community formation declines and other factors become more important. Meanwhile, this change in priority factor may not mean the end of pyrogenic succession, and at this stage, communities are not equal to climax vegetation observed at non-burnt territories in Kalmykia. Though 10–15 years of pyrogen cyclic is typical for sagebrush communities in Western Kazakhstan, not far from the studied “Chernye Zemli” Reserve, for other sagebrush communities worldwide, the time of self-restoration after fires can last for decades [
46,
47], and so pyrogenic succession longer than 10–15 years can take place in Kalmykia in modern conditions.
Notwithstanding their limitations, space-for-time studies [
16,
26] provided valuable insights into the average state of plant communities with the time since fire. In fact, the actual state of communities in specific areas in different periods may vary due to the characteristics of weather conditions, micro topography, grazing and other factors. However, we consider this approximation sufficient for the analysis presented in this research, since we compare two groups of species: those found and not found after the fire, and do not draw conclusions based on each target species. The mean pre-fire projective cover was significantly higher for fire-tolerant species, i.e., those found after the fire, than for fire-intolerant species—not detected after the fire. There are several possible but not mutually exclusive reasons for this:
- (a)
For some species, conditions are not optimal both before and after the fire, so their projective cover is low both in the pre-fire and post-fire stages;
- (b)
During a fire, individuals of species with higher prevalence are more likely to be found in loci less affected by open fire and high temperatures, and thus survive;
- (c)
After a fire, seeds invade from adjacent territories, and the supply of diaspores is higher for species with a higher number of individuals (vicinism according to [
48]);
- (d)
For the introduction of some minor species, an important role can be played by the primary plant cover, which can provide partial protection of seedlings from negative environmental factors due to shading or moisture retention in the accumulated dead litter in the spring [
16]; therefore, such plants cannot appear in the early stages of pyrogenic succession.
Although it is often believed that frequent fires reduce the vascular plant species diversity in arid ecosystems and form special pyrogenic communities [
14,
49], the effects of fire on the characteristics of arid phytocenoses are ambiguous. Various publications note both positive and negative effects of the pyrogenic factor on communities [
50]. Some recent studies have investigated methods of excluding fire from some grasslands in order to maintain high biodiversity [
51].
According to our data, frequently and rarely burned areas at the territory of the “Chernye Zemli” Nature Reserve and its surroundings do not differ in the total number of species found (49 and 47, respectively) and are similar in species composition (>75% of common species). However, there are some species that are typical for areas with different fire frequencies. According to NMDS, subshrub communities are more likely to be found on rarely burned areas, whereas xerophytic-grass communities are more likely to be found on frequently burned areas. In addition to the time since fire, an important factor in the structure of pyrogenic communities is the history of pyrogenic impact, as has been shown for grass communities in South Africa [
52]. There, maximum species richness was achieved with a combination of different fire regimes. This idea corresponds with the basic strategy of using prescribed burning for maintaining biodiversity in managed territories, both for woods [
51] and treeless vegetation [
37]. It is also possible that communities in our study were transformed by frequent fire over millennia, as with some other treeless ecosystems [
53,
54]. This is consistent with the concept of the historical genesis of arid communities under the influence of the pyrogenic factor [
55]. It is known that periodic fires played a major role in the evolution of arid grasslands (including steppes) over millions of years [
53,
54], so the species in these communities are well adapted to persist with a range of fire regimes.
Despite the similar species richness, four indicator species were identified for frequently burned areas (
Stipa sareptan,
Medicago orthoceras,
Crepis sancta, and
Filago arvensis L.) and four for rarely burned areas (
Lepidium perfoliatum,
Artemisia lercheana,
Chorispora tenella (Pall.) DC, and
Tanacetum achilleifolium). Projective cover on frequently and rarely burned plots differed most markedly for
Artemisia lercheana and two cespitose grasses,
Poa bulbosa and
Stipa sareptana.
P. bulbosa was more common in areas with rare fires, and
S. sareptana was more common in areas with frequent fires. Since a lot of mortmass accumulates in
S. sareptana communities, and
P. bulbosa, on the contrary, forms very little mortmass due to its small size, it can be assumed that the probability of fire increases with the increasing role of
S. sareptana and decreases with the increasing role of
P. bulbosa. The high projective cover of
S. sareptana on frequently burned areas may be maintained due to fast recovery or high tolerance, as there is evidence that large-turf species tolerate fire better than small-turf species [
9,
56].
Artemisia lercheana is more common in rarely burned areas, which is consistent with previous studies showing a decreasing role of sagebrush under the influence of fires [
14,
57,
58].


 ,
, 






{kind=link}
{kind=link}
{kind=link}
{kind=link}