1. Introduction
The grand challenges of climate change and sustainability have led to a growing focus on the production and use of renewable materials, such as macro and microalgae. As photosynthetic organisms that sequester CO
2 during growth, algae can help society transition away from fossil fuels and assist in achieving emission reduction goals (Paris Agreement 2015 [
1]). Furthermore, algae are a renewable biomaterial that can be used to produce a range of products, including fertilizers and animal feeds, human food supplements, nutraceuticals, specialty chemicals, and bioenergy [
2,
3]. Algae have also been demonstrated to assist in bioremediation applications as diverse as mine-tailings water treatment [
4,
5] and aquaculture wastewater treatment [
6,
7]. Such examples demonstrate the potential to use algae to develop circular economy solutions within existing production systems.
Macroalgae are a plant-like organism that can grow in either fresh or saltwater, depending on species. Growth rates of macroalgae are very high and saltwater species such as
Ulva ohnoi (Chlorophyta) can achieve substantial growth rates [
7]. Other species, such as the freshwater macroalgae
Oedogonium intermedium (Chlorophyta), show promise in energy production applications [
8] and as a supplement to reduce cattle greenhouse gas emissions [
9,
10]. Such examples demonstrate potential for such algae species to be useful products of industrial processes.
There has been comparatively little attention to the optimization of macroalgae production processes. Raw algae products are typically of low value, and optimization of all processing steps is required for economic viability. The high water content of the centrifuged or filtered macroalgae (80–90%
WB) is an issue for many of the potential production pathways. For example, high moisture content can cause substantial problems in biodiesel production [
2], and conventional pyrolysis of algae for the production of biogas or biochar requires very low moisture (0–15%) for efficient processing [
11]. The very high ratio of moisture to dry mass also incentivizes drying to reduce weight and volume for transportation to markets and/or additional processing. Furthermore, the removal of moisture increases the storage life of algal products through the inhibition of spoilage due to unwanted microbial growth. However, drying is an energy-intensive process, potentially responsible for the single highest energy cost in an algae production process [
12]. Optimization and careful design of drying equipment are therefore priorities for cost-effective development of macroalgae bioresources.
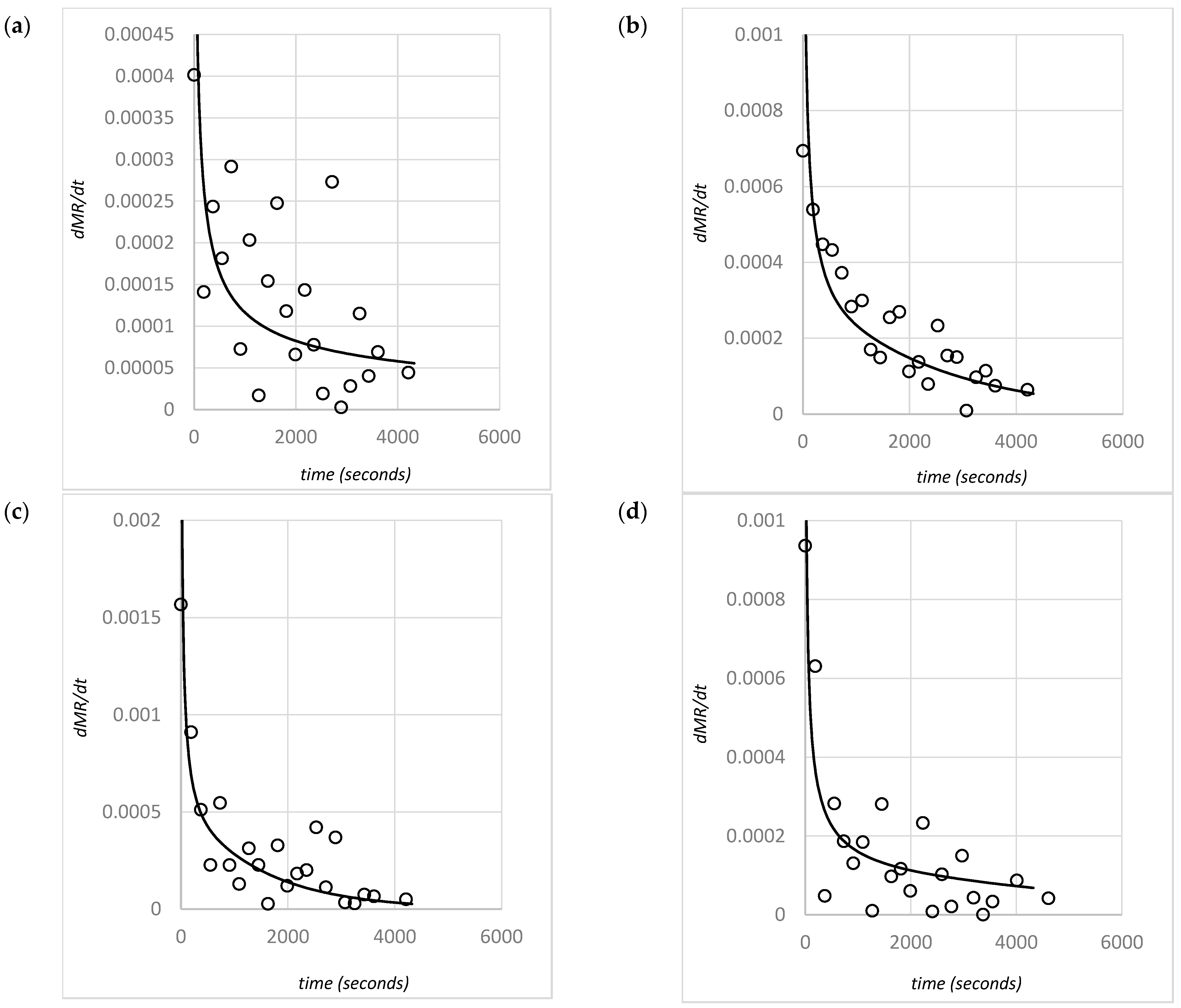
Convective dryers are widely used in biomass drying applications and provide significantly enhanced drying rates compared with other dryer types. In a convective dryer, the drying process for materials with internally bound moisture, such as macroalgae, involves three simultaneous stages: heat transfer to the material (usually via convective transfer); mass transfer of moisture (typically diffusion) to the exposed material surfaces; and convective mass transfer of moisture to the surroundings [
13]. The mass transfer through the material is partially driven by a moisture concentration gradient between the material and the surroundings. This leads to a rate of drying that typically reduces over time (i.e., the falling rate) as the concentration of moisture throughout the algae material decreases [
14], and such drying is referred to as diffusion-controlled drying. The drying rate, typically obtained through experimentation, is an important characteristic of a material as it directly enables the determination of dryer residence times and equipment sizing. Modeling such rates can also be useful to understanding the fundamental processes that are occurring. Key dryer design variables that can potentially affect the drying rate include drying gas temperatures and gas flow rates (i.e., the gas velocity). However, it is also important to understand the impact of other material characteristics that influence biomaterial drying rates, such as the compressible nature of algae (represented by bulk density) as well as material shrinkage (represented by a material’s characteristic length).
While there is some evidence to suggest that macroalgae shrink during the drying process [
15], there have been relatively few studies of macroalgae drying reported in the literature. Vega-Galvez et al., 2008 [
16] modeled drying of the brown macroalgae
Macrocystis pyrifera, Gupta et al., 2011 [
17] studied drying of the macroalgae
Himanthalia enlongata, Uribe et al., 2017 [
18] studied the drying kinetics for the brown macroalgae
Durvillaea antarctica, and Lemus et al., 2008 [
19] and Tello-Ireland et al., 2011 [
20] investigated drying of the red macroalgae
Gracilaria chilensis. In all these examples, the impact of temperature on the diffusion coefficient and drying rate was examined (in the range 40 to 80 °C), while the gas velocity remained constant, with a value chosen in the range 1.5–2 m/s. Recent work by Walker et al., 2020 [
14], which we build on in this paper, looked at the drying of
U. ohnoi and
O. intermedium for a range of temperatures, but also under constant gas velocity conditions. In a purely diffusion-controlled drying process where surface convection is not rate limiting, the impact of gas velocity on drying rates would be expected to be negligible. For example, when Mohamed et al., 2007 [
21] investigated the thin layer drying kinetics of the macroalga
Gelidium sesquipedale at very low gas velocities (0.04 to 0.15 m/s), they found purely falling rate drying (i.e., diffusion-controlled) and a minimal influence of gas velocity. However, when Djaeni and Sari., 2015 [
22] undertook convective drying of the macroalga
Eucheuma cottonii at much higher gas velocities of 5 m/s and 7 m/s, results showed an increase in drying rate with increasing gas velocity. Similar observations of the influence of gas velocity on drying rates in diffusion-controlled systems have also been made for biomaterials that have similar compressible and porous characteristics to macroalgae. This includes Chkir et al., 2015 [
23] in their work on drying brewers’ grain and research undertaken by Bezzina et al., 2018 [
24] on drying sugar cane bagasse fiber. Both these studies found that drying rates increased with increasing gas velocity, despite being entirely falling-rate or diffusion-controlled processes.
In Bezzina et al.’s, 2018 [
24] study of bagasse fiber, bulk density was also found to have a significant influence on the drying rate. However, studies of the influence of bulk density on drying of compressible materials are extremely rare in the literature and our understanding of the mechanisms that occur in these types of biomaterials is very limited. Some studies report relationships between effective diffusivity and a material’s porosity and tortuosity (see, for example, [
25,
26]). However, utilizing such relations requires a priori knowledge of both porosity and tortuosity, with the latter being very challenging to define for a heterogenous and tangled fiber such as macroalgae. Use of such relations is further complicated by materials that exhibit shrinkage, such as macroalgae, for which applicable relations are not well-developed. A more practical and measurable material property is bulk density, which can be more useful when developing models for industrial applications such as dryer control and dryer design.
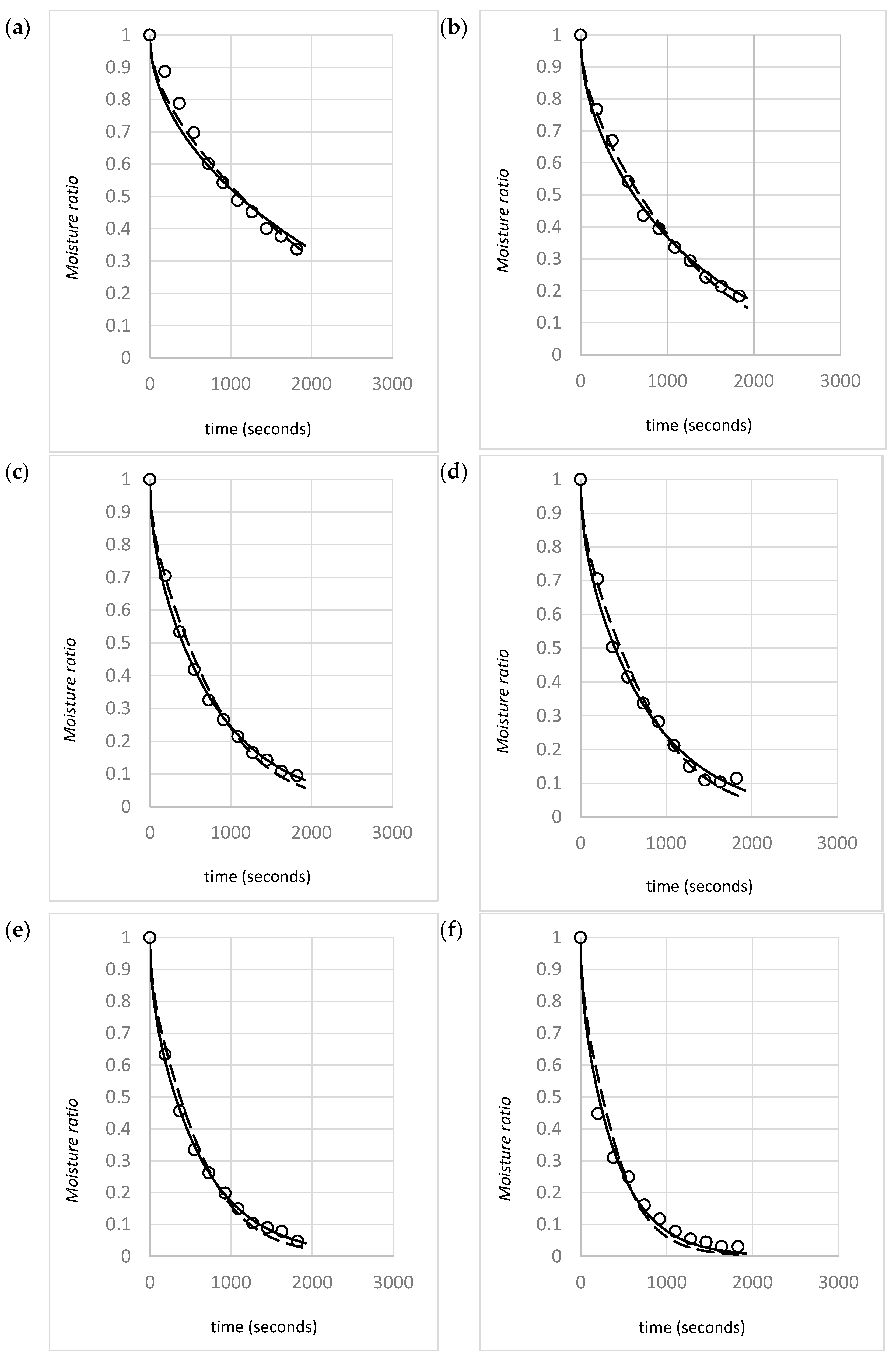
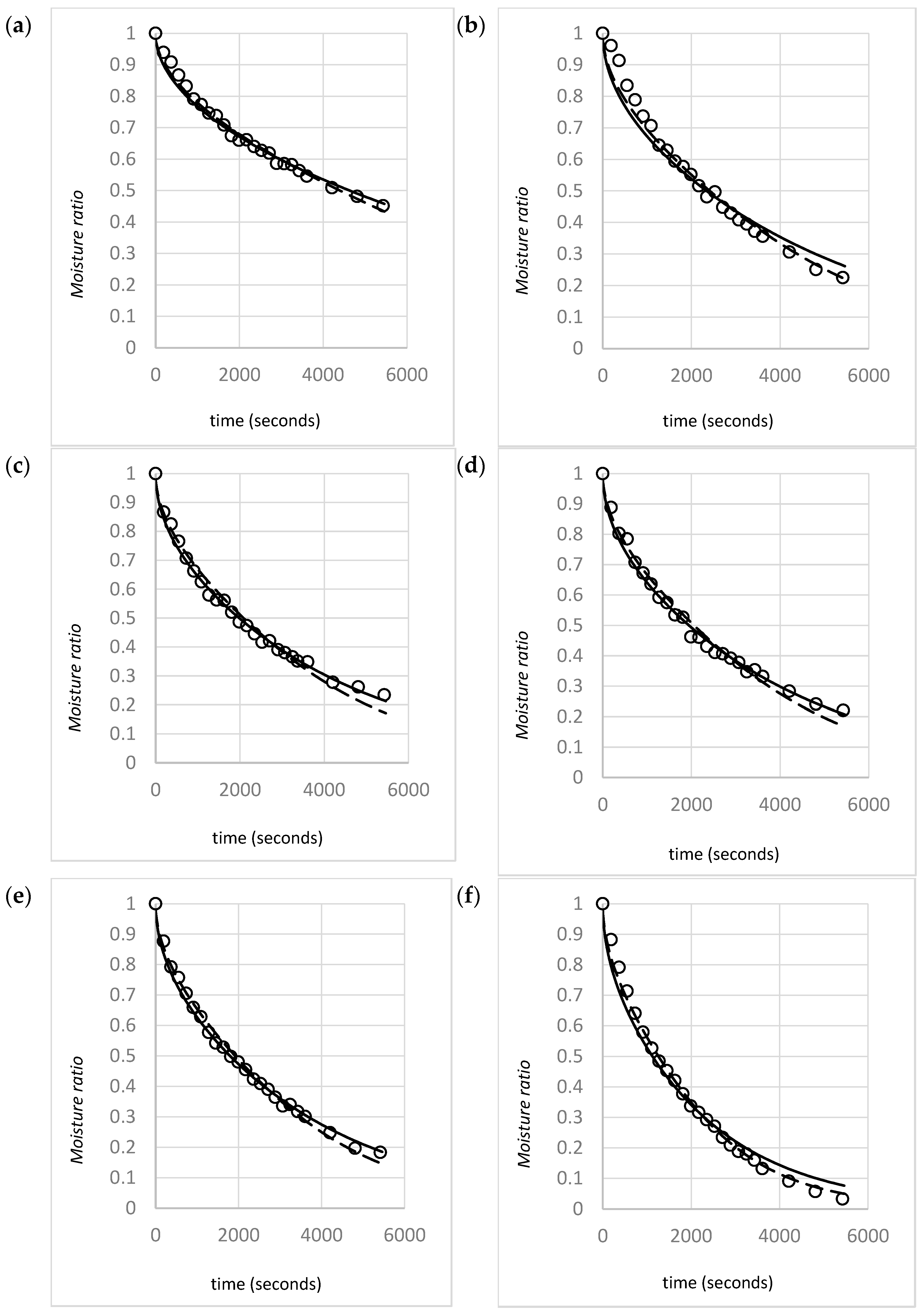
This study investigated the influence of gas velocity, density, and shrinkage for two different species of macroalgae (U. ohnoi and O. intermedium). Pragmatic drying rate models based on Fick’s Second Law were developed for a range of initial material bulk densities and typical industrial drying air velocities, with inclusion of the effects of material shrinkage. Models of the effects of these variables on drying rate parameters, such as the effective diffusivity, are provided. The aim of the modeling is to provide an accurate representation of the drying rate for the target species. The model is intended to be sufficiently pragmatic that it can utilized by industry to predict dryer performance or optimize dryer design. Furthermore, the modeling also seeks to provide insights into the underlying mechanisms involved in drying porous biomaterials. Based on the observed effects of density and gas velocity, an analysis of the validity of model assumptions for compressible biomaterials is provided, and a mechanism describing the effect of gas velocity on drying rates is proposed.
3. Theory
Modeling of drying for materials with internal moisture is typically performed by taking a mechanistic approach that solves fundamental heat and mass transfer relations using differential equations or via an empirical/semi-empirical approach, using fitted equations and lumped parameters. The mechanistic approach is limited by both a lack of fundamental properties describing macroalgae (such as heat and mass transfer coefficients) as well as the difficulty in solving the resulting partial differential equations, which limit the potential industry uptake. In this work, a more pragmatic semi-empirical approach is taken that solves Fick’s Second Law of Diffusion (F2L) and lumps the effects of material density and gas velocity into a single diffusion coefficient (
), whilst still accounting for material shrinkage. Such an approach is widely accepted for biomaterial drying and has been previously used to represent drying of many different types of biomaterials, including several other macroalgae species [
16,
18,
19,
29,
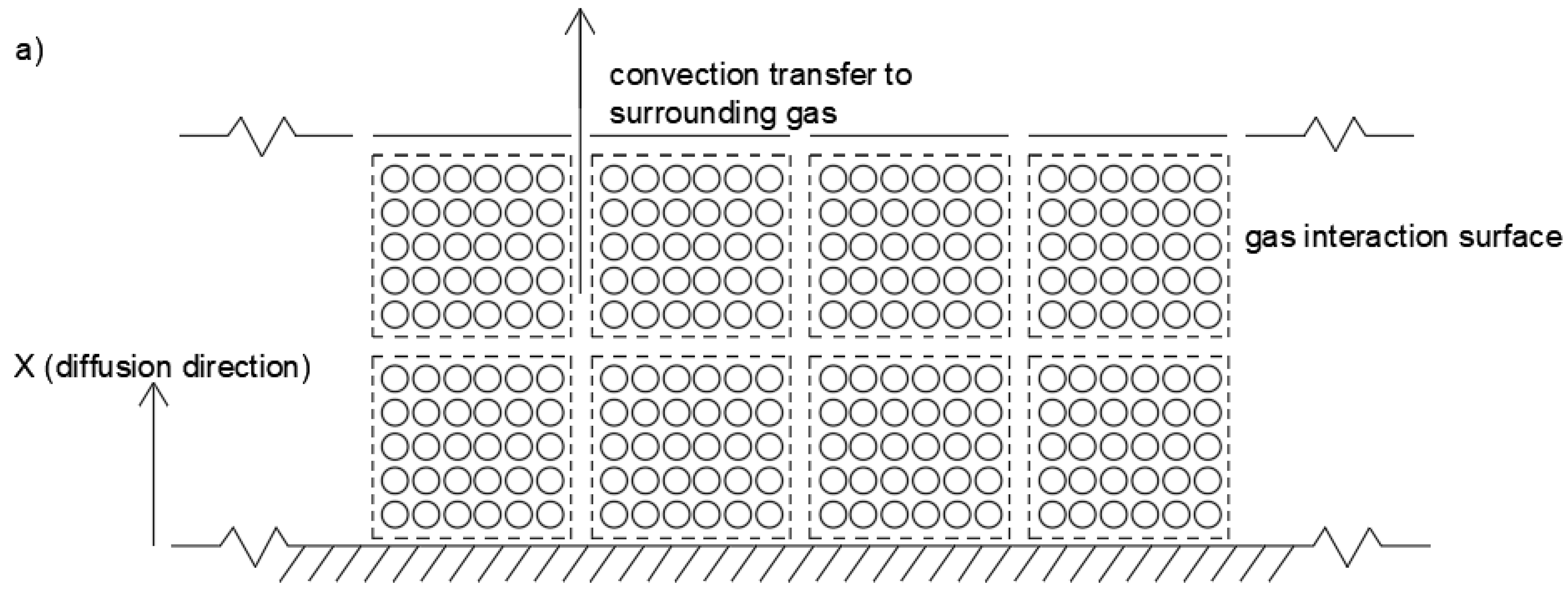
30]. F2L for a one-dimensional semi-infinite slab of material (shown in
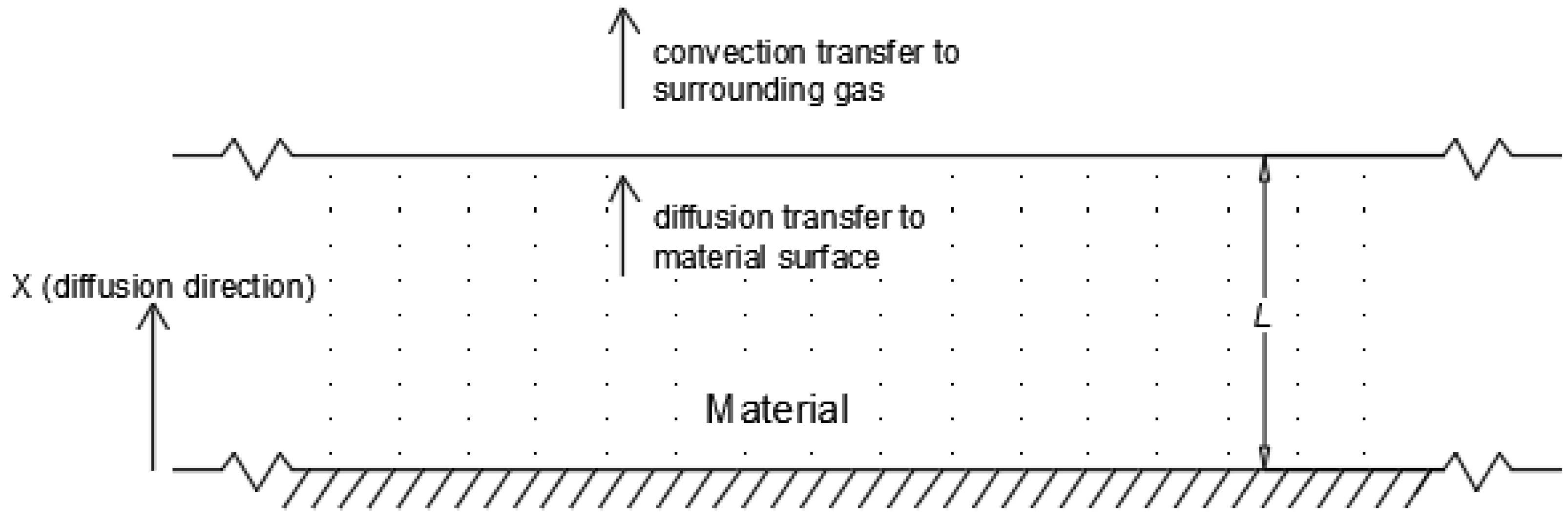
Figure 3) is given by in the following equation.
where
is the concentration at a given point and time,
is the distance of the point from the boundary surface,
is time, and
is the effective diffusion constant. In this model, a number of assumptions are utilized: diffusion represents the overall drying mechanism; there is a uniform initial distribution of moisture throughout the material; drying is one-directional and moisture vapor travels out of the sheet surface; moisture on the sheet surface is immediately removed via bulk convective mass transfer, ensuring that the surface concentration is constant and at the equilibrium concentration and that diffusion is the rate-limiting step; and the diffusion coefficient is not a function of distance or time. A conceptualization of the model geometry can be seen in
Figure 3 and the mathematical expression of the model boundary conditions for the PDE can be found elsewhere [
27]. When F2L is solved analytically for the moisture ratio (
) for a slab geometry of thickness
the following series solution is obtained:
The parameter
is a lumped parameter that represents the effective diffusivity of the material and is therefore some function of all variables affecting the drying rate. The
parameter is used as the manipulated variable in parameter estimations and optimized values for
for each experimental data set are calculated. However, the impact of temperature is often described using an Arrhenius expression for
(Equation (3)), whereby
is the lumped parameter used to describe the effects of other variables on the drying rate. A strong Arrhenius relationship between effective diffusivity and temperature has been demonstrated in previous radiative drying research for both species of algae (
U. ohnoi and
O. intermedium) [
14].
The analytical solution to Fick’s Second Law was fit to experimentally derived moisture ratio versus time data sets. Model fitting was performed by minimization of the total sum of square errors between the model and experimental data (summarized in Equation (4)) for each data set, with the model representing the best fit to all three sets of data at a given condition. Ten terms of the infinite series (
n = 10 in Equation (2)) were used in modeling to ensure convergence in the infinite series solution [
31]. These calculations were performed using Microsoft Excel using the solver function with the GRG nonlinear solver method assuming constant relative variance in the experimental data. The objective function is summarized in Equation (4), where
m corresponds to the number of data sets. The objective function (
) was minimized by varying the magnitude of the effective diffusivity parameter (
).
Modeling Shrinkage
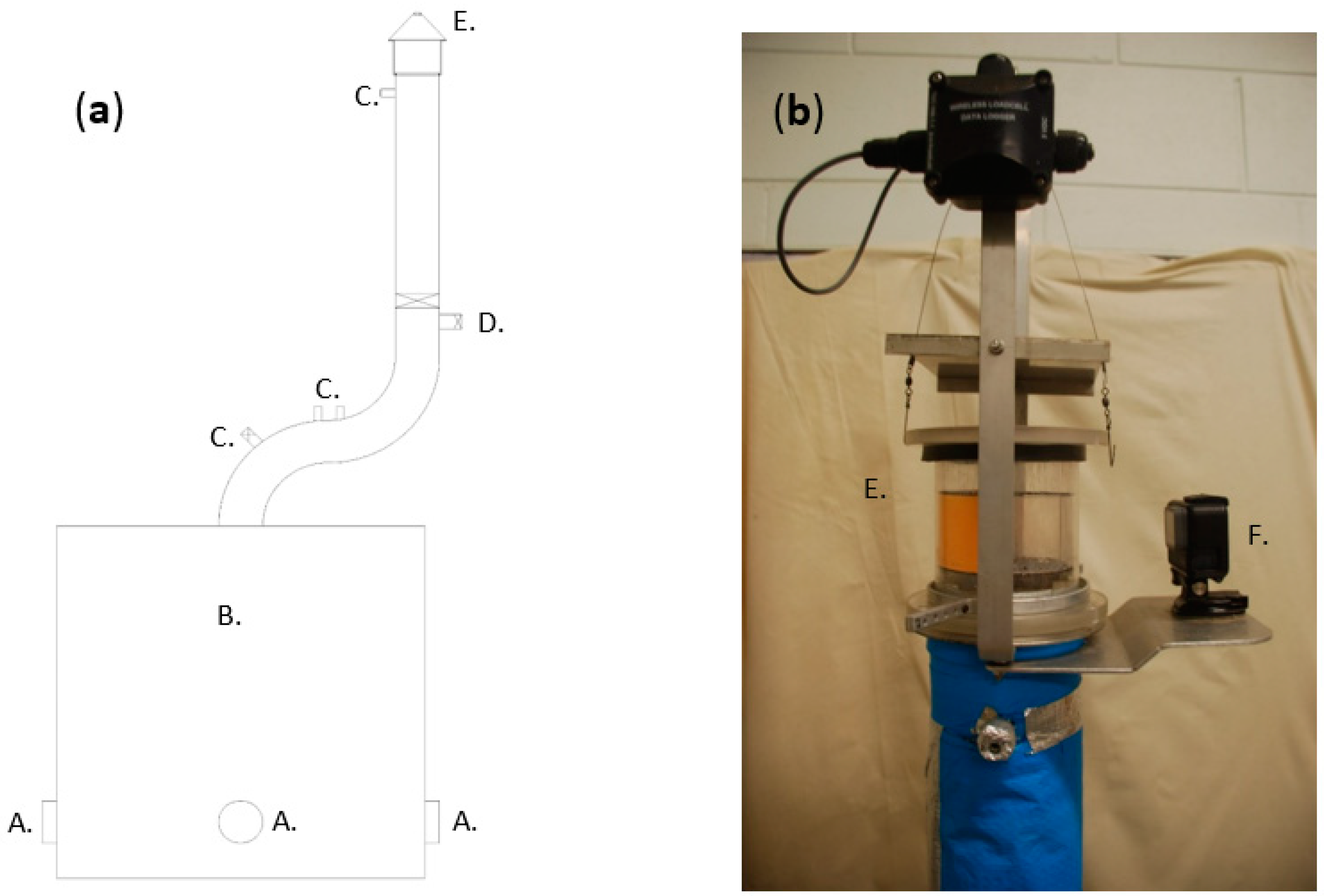
Image capture and shrinkage image analysis were performed in triplicate for both algae species. For
U. ohnoi, the drying conditions analyzed were 50 °C, 0.7 m/s air velocity, and 66 kg/m
3 bulk density, while for
O. intermedium the conditions were 50 °C, 2.0 m/s air velocity, and 66 kg/m
3 bulk density. These images were used to determine the change in the slab width (i.e.,
) during drying for both species. Slab widths were converted to a non-dimensional property via:
, where
is the slab width at time
,
is the initial slab width, and
is the final slab width (at
). The difference between the initial and final slab width is given by
. The properties of
and
are straightforward values to determine experimentally. The value
represents the fraction of volume losses that are yet to occur during drying. For example, at
,
is equal to 1 and indicates that no volume loss has occurred; at
,
is equal to zero and indicates that all volume losses from drying have occurred. This makes both the moisture content and slab width non-dimensional values with the same range, facilitating comparison of moisture and volume losses over time. Examples of
vs.
are shown in
Figure 4 for the raw data sets. Correlation of
and
obtained from the experimental data was performed in SPSS and statistical analysis demonstrated
and
to be linearly correlated with statistical significance at the 95% confidence level across all data sets.
To include the impacts of volume loss in the F2L drying rate model described in Equation (2),
was equated to
(Equation (5)). The slab width at any time
(
) was related to the
value at the same time (
) via Equations (6) and (7) rather than from direct measurement of length versus time.
For convenience and in the interests of developing a pragmatic model to predict
versus time profiles, the predicted
utilizes the previous time-step’s
. In essence, it assumes that each new length represents the slab’s dimensions up to that point. Although not formally correct, this approach avoids having to solve the full PDE system described in Equation (1), but with varying boundary conditions. The method calculates
MR at time
t through approximating drying of a slab of width
Lt from
t = 0 to
t =
t, but given a sufficiently small time-step (one-minute steps were used in this study) this approximation works well and is simple to implement. Thus, by substitution, F2L including shrinkage is as follows (Equation (8)):
Model fitting for the optimized value that best fit the experimental MR versus time data was performed with both the constant model (Equation (2)) and the shrinkage model (Equation (8)).
5. Discussion
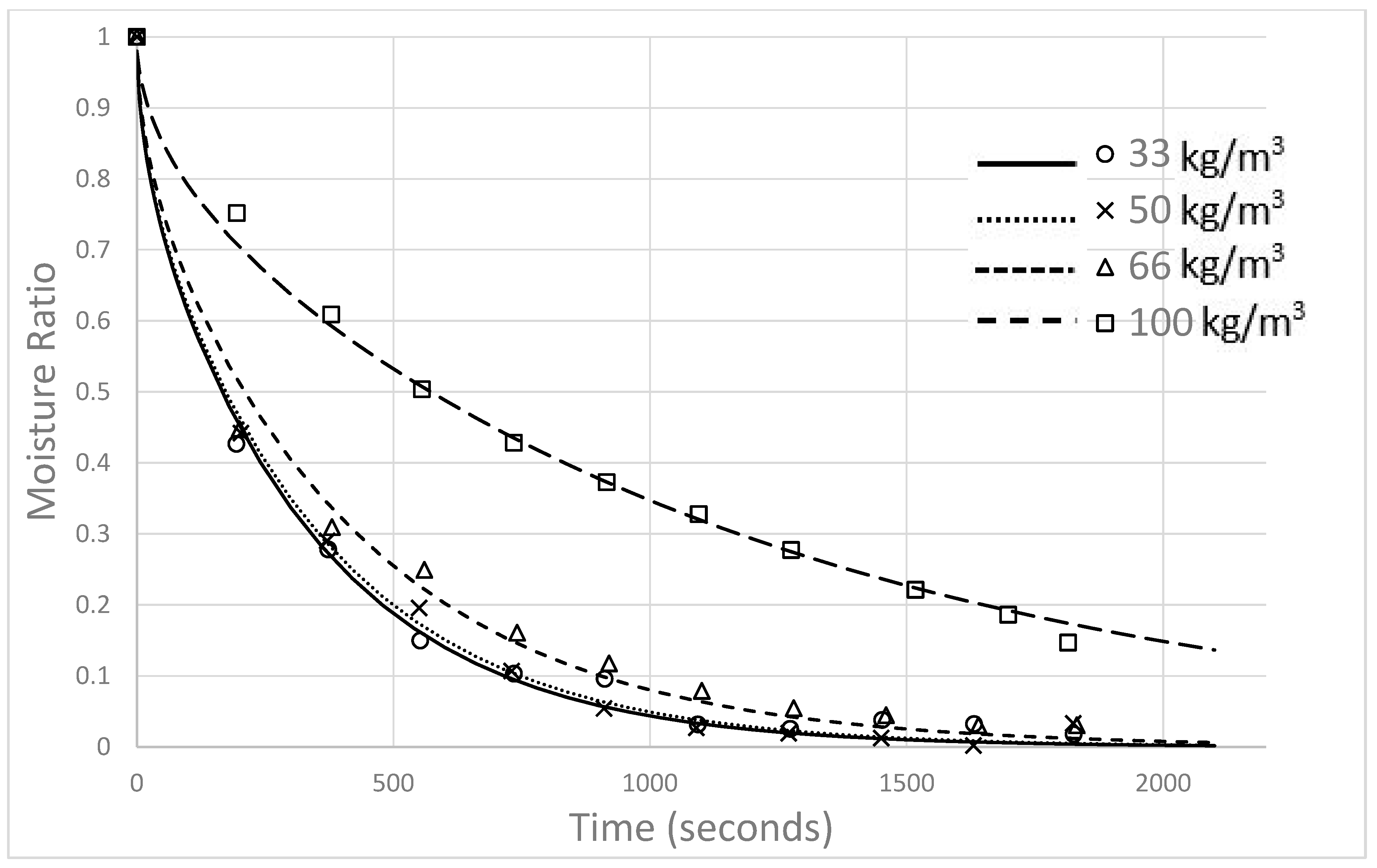
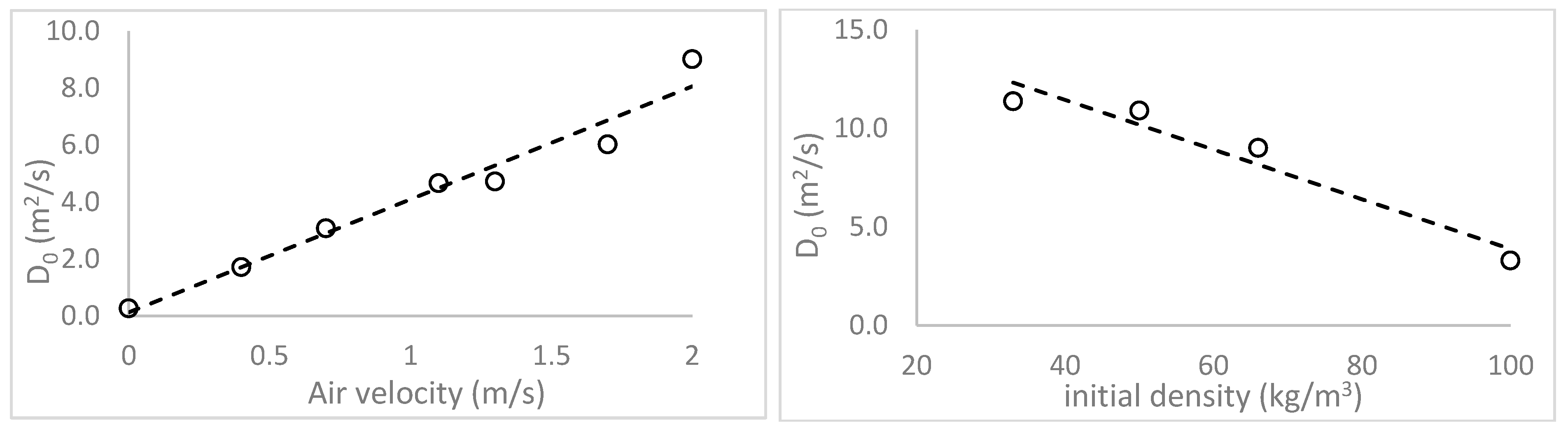
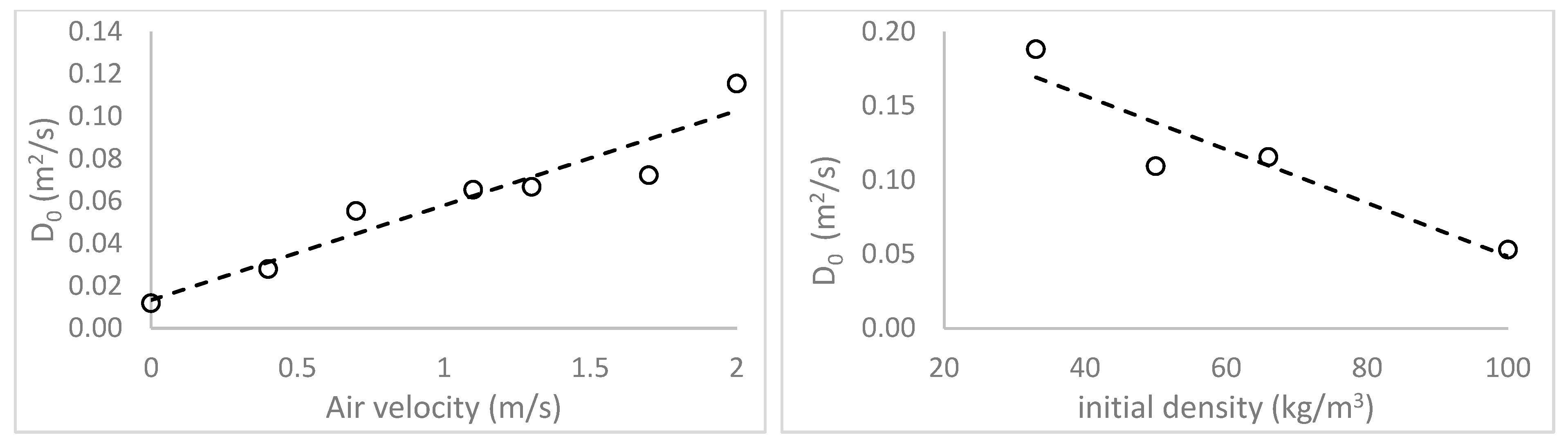
The impact of material density on the drying rates of macroalgae biomaterials was as expected, with higher drying rates for less-dense clumps of algae. Only two studies were identified in the literature that examined material bulk density effects on drying rates [
24,
33]. Both were focused on the drying of sugar cane bagasse and showed trends similar to those observed in this research. However, the observed increases in drying rate for increasing gas velocity values are less intuitive. At first glance, such results appear to violate the fundamental assumption of a diffusion-limited system, whereby surface convection (driven by gas velocity) is assumed to
not be the rate-limiting step. Typically, in a purely diffusion-based process, increased surface convection driven by increased gas velocity would not be expected to lead to greater rates of drying. However, several other studies have also shown a dependency between air velocity and diffusivity [
22,
23,
24,
25,
26,
27,
28,
29,
30,
31,
32,
33,
34]. For example, Chkir et al., 2015 [
23] tested effects of gas velocity on the drying kinetics and properties of brewer’s grain and also showed an increase in drying rate as the gas velocity increased. Furthermore, Bezzina et al., 2018 [
24] studied convective drying kinetics for sugar cane bagasse and found both diffusion-limited drying and that increased gas velocity increased the drying rate, similar to the gas velocity effects shown in this research, for macroalgae. A common theme in these results is that materials exhibiting such effects are all compressible and porous with minimum particle dimensions being smaller than bulk material dimensions.
To explain the impact of bulk material density, a mechanism involving gas penetration is proposed. Increases in material bulk density decrease the void fraction in the macroalgae slab. A decrease in void fraction results in an increase in material physically blocking the diffusion pathways to the material surface (i.e., presenting a physical resistance to diffusion) and therefore a decrease in the effective diffusivity is observed with increased density. The upper limit of this effect would be at zero void fraction, where the material can effectively be considered as a solid block, such as that represented in
Figure 3, with
as the full slab width. Zero void fraction would imply that there should be no opportunity for gas to penetrate into the slab, as would be expected for drying vegetable and fruit slices.
Considering the impact of gas velocity on the drying rate, experimental results in this work show that drying rates for both macroalgae increased as the drying gas velocity increased. In a porous biomaterial, it is expected that increases or decreases in air velocity cause more or less gas penetration into the particle bed, respectively. The hypothesis posed in this work is that increased penetration via a higher gas velocity works to reduce the average distance between the particle surface and the surrounding gas interaction surface (i.e., reduces the distance denoted as
in
Figure 3). However, in this mechanism it should be emphasized that the drying gas interaction surface is not the external edge of the material slab as defined in
Figure 3, but, instead, the gas interaction surface where convection occurs is internal to the material slab due to the effect of drying gas penetration.
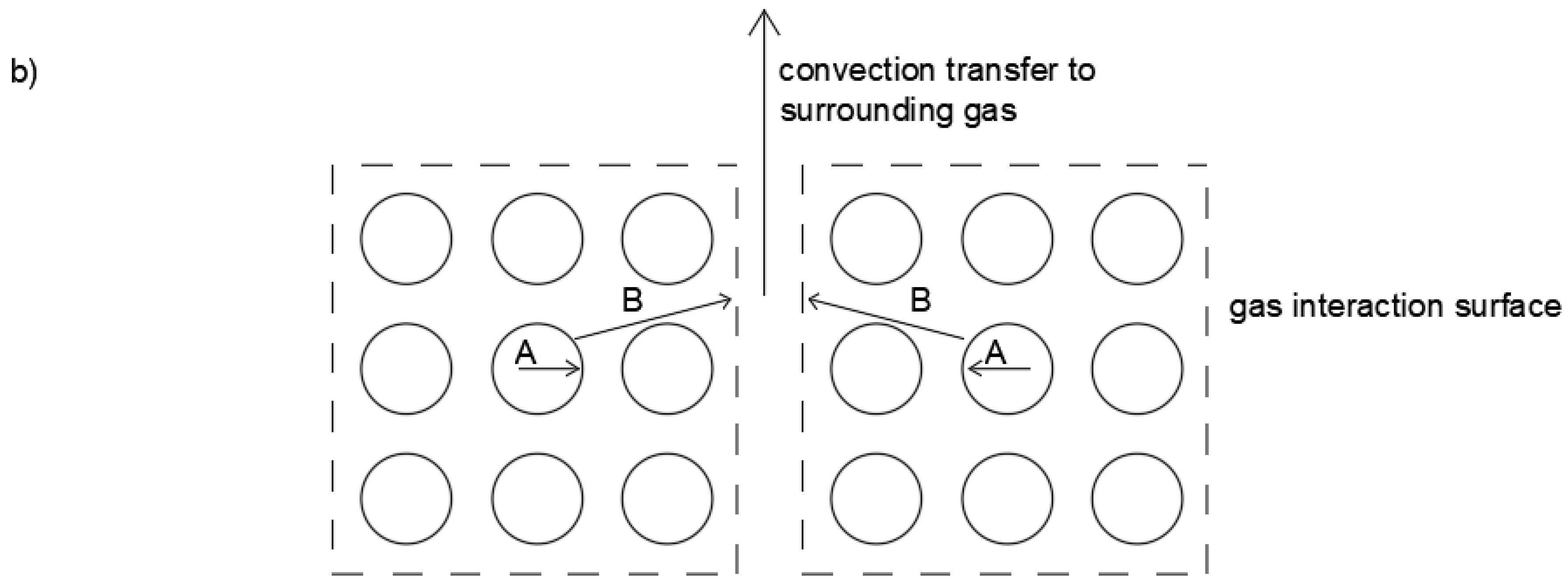
Figure 16 illustrates this concept.
Comparing the traditional view of diffusion through a slab (
Figure 3) with
Figure 16, the impact of air flow can be understood by considering flow penetration acting to separate the bulk material into smaller ‘blocks’ of particles. Convection would then occur at the surface of these internal blocks. Thus, increases in the gas flow rate cause increased slab penetration, which breaks up the bulk material into even smaller blocks. The lower and upper limits of this mechanism are where no flow results in no penetration and
is best represented as the full slab width (
Figure 3). The upper limit occurs at some maximum gas velocity where the blocks are reduced to the dimensions of an individual particle or cell width. A maximum gas velocity would reduce the geometry to that described in
Figure 16b, with
being equal to a single particle width. The point where this occurs could also be referred to as a ‘critical’ velocity in that further increases in gas flow beyond this maximum should not affect drying rates. Such a critical velocity was not observed in this work, although the existence of such a limit has been hypothesized elsewhere [
32].
In the specific case of macroalgae where moisture is trapped within discrete cells and also within a tangled matrix of algae strands, we hypothesize that the transport process occurs via two mechanisms. Referring to
Figure 16b where dotted lines represent the surfaces where convection mass transfer occurs, Mechanism A represents internal moisture diffusion through an individual cell or particle to that cell’s/particle’s surface, and Mechanism B represents moisture transfer through a given ‘block’ to the gas interaction surface. Given the incompressible nature of a single particle or cell consisting almost entirely of water, it is, instead, moisture transfer through the block (Mechanism B) that is assumed to be affected by changes in both bulk density and gas velocity. Thus, it is expected that this stage is the rate-limiting step for conditions where a relationship between air velocity and/or material bulk density and drying rates exists. Furthermore, because experimental drying data demonstrate a single falling-rate period under all experimental conditions, Mechanism B (as the rate-limiting step) is likely a diffusive transfer, supporting the use of F2L as a theoretical framework for such materials. Targeted experiments would be necessary to prove the validity of the proposed mechanism. However, it is recommended that materials with more defined dimensions and exhibiting less variability in drying rate data, such as bagasse fiber, be utilized for such work.





















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}