
Early Stages of Ex Vivo Collagen Glycation Disrupt the Cellular Interaction and Its Remodeling by Mesenchymal Stem Cells—Morphological and Biochemical Evidence

 , , ,
, , ,
Abstract
:1. Introduction
2. Results
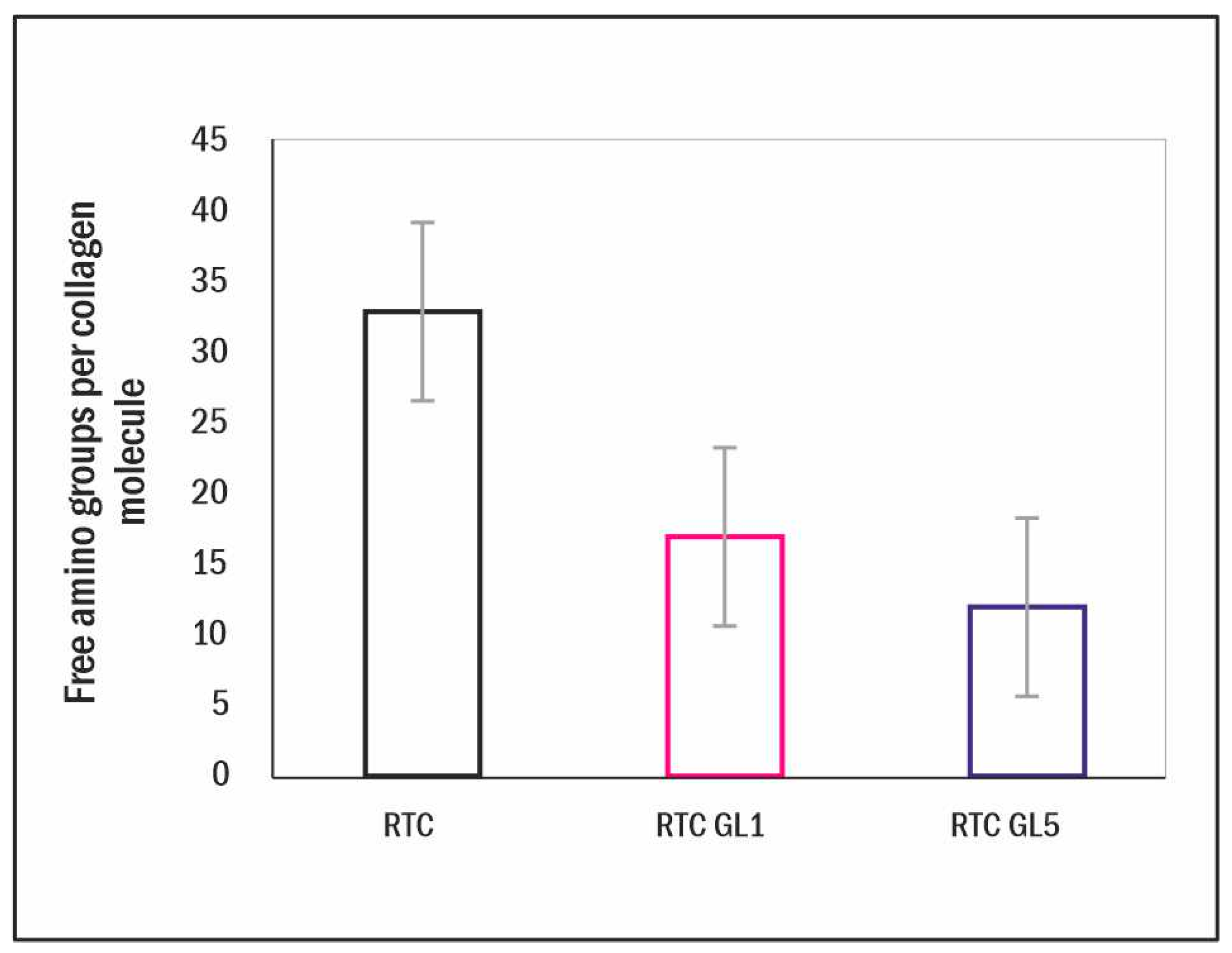
2.1. Measurements on the Extent of Glycation
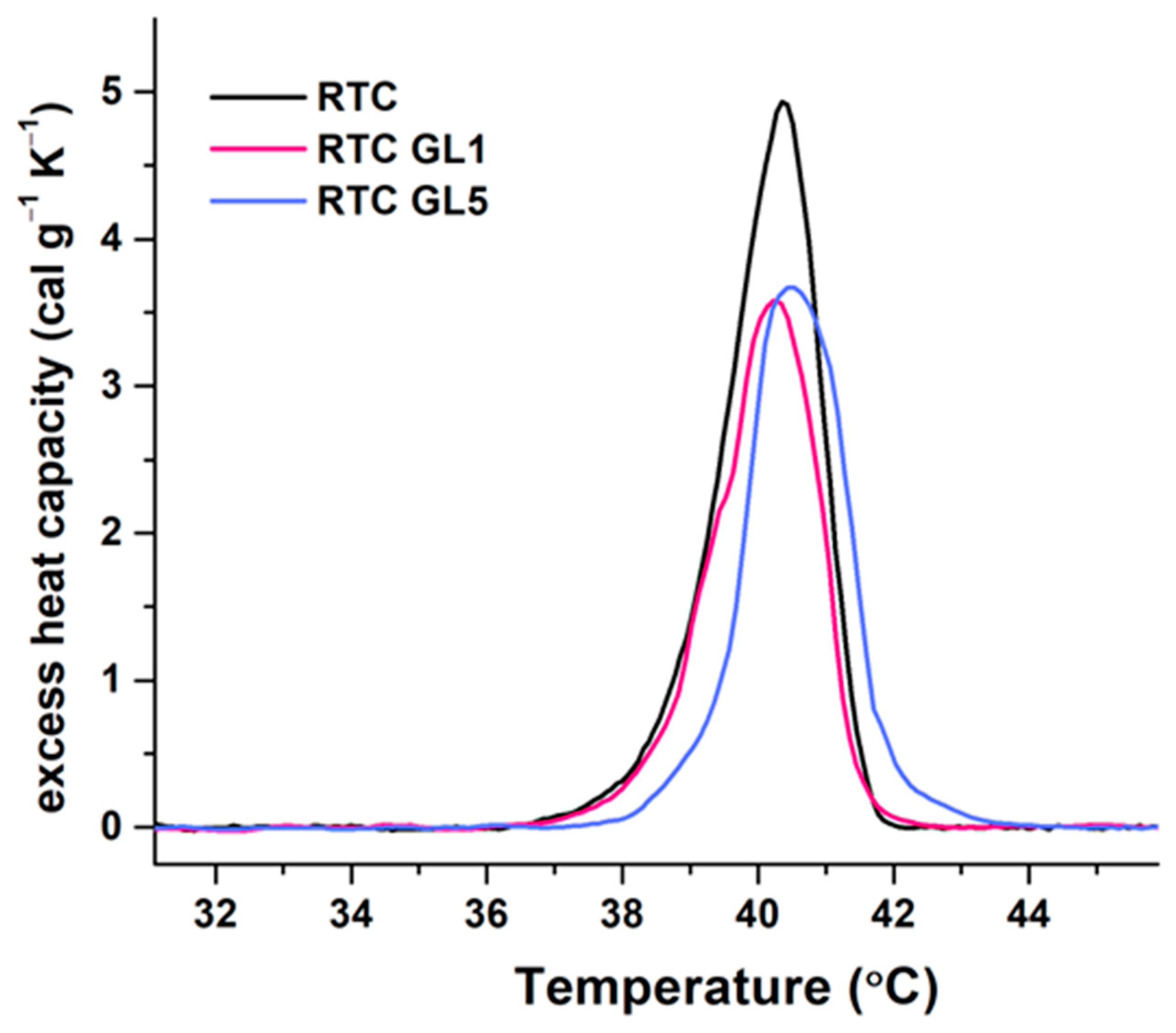
2.2. Differential Scanning Calorimetry (DSC) Measurements
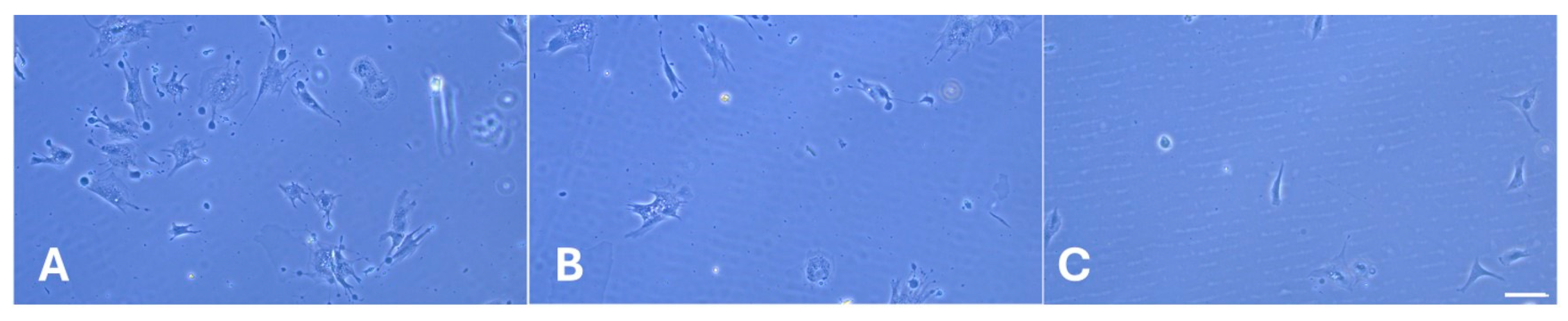
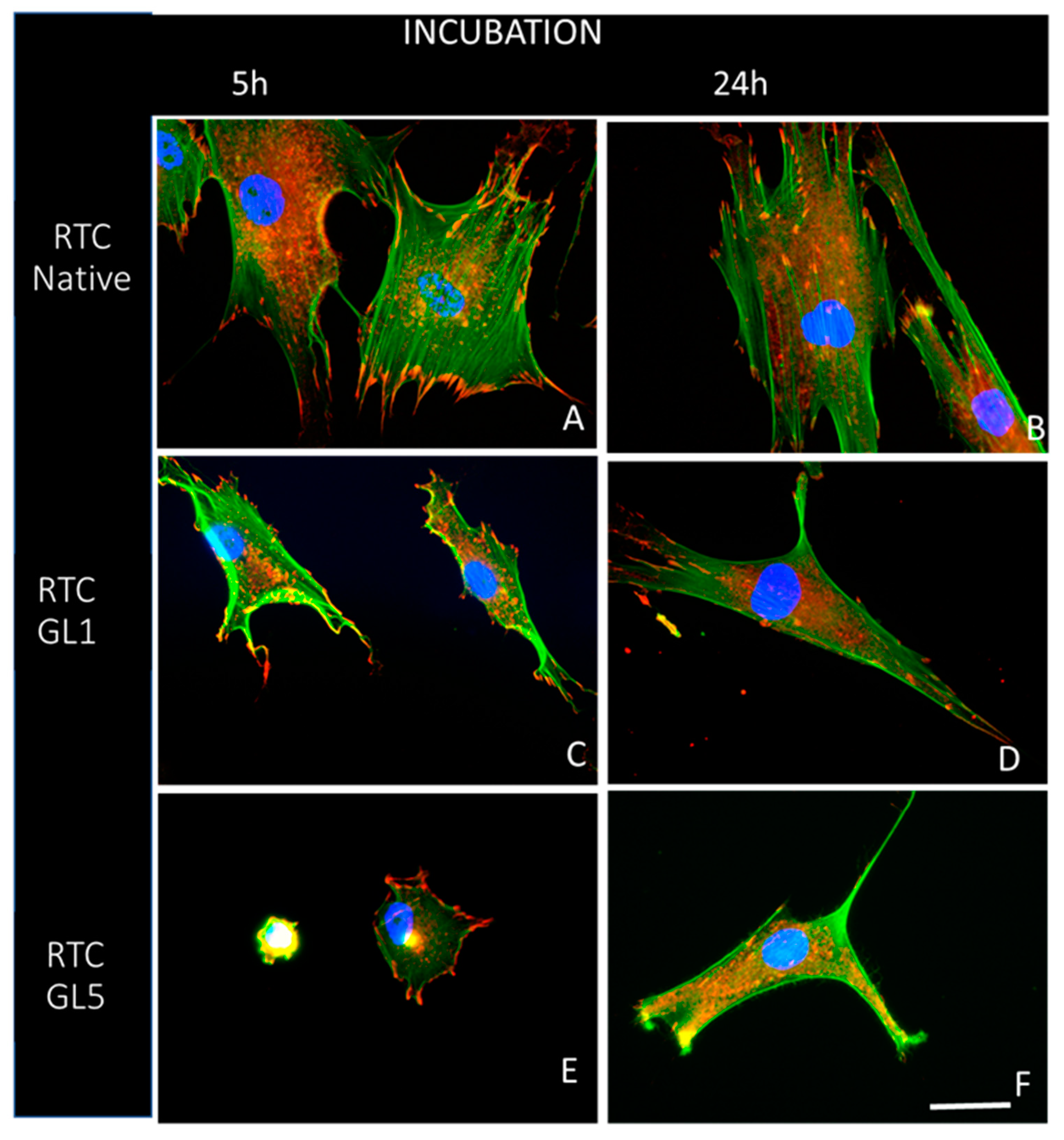
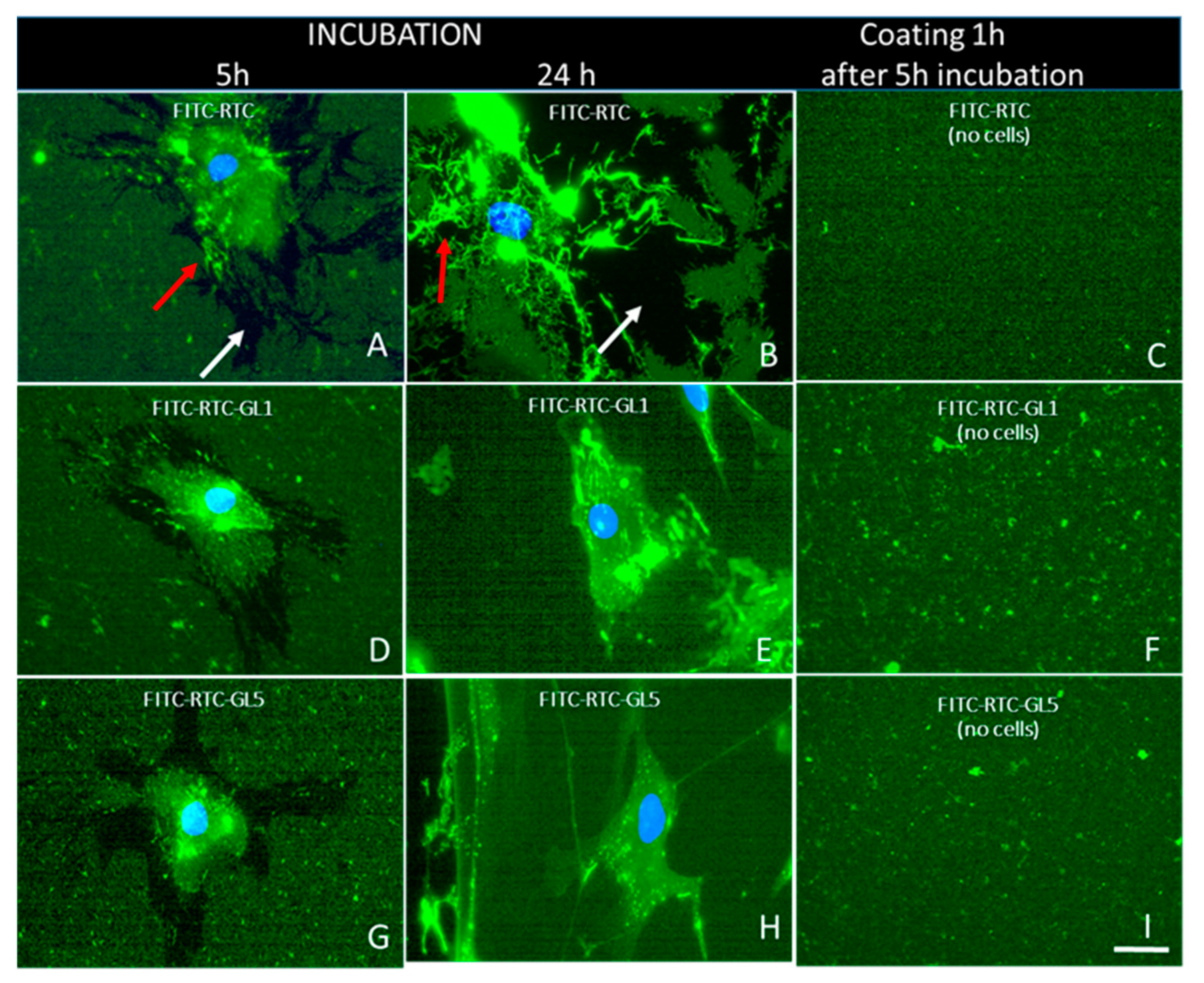
2.3. Morphological Observations
2.4. Cell Viability Testing
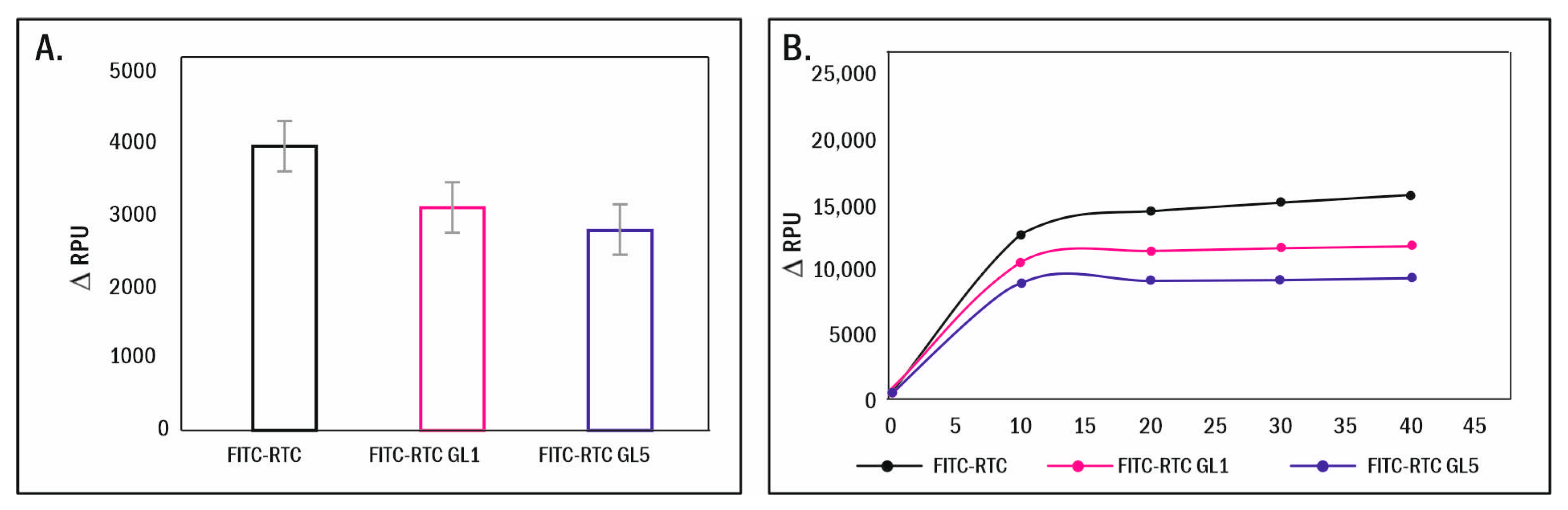
2.5. Enzymatic Remodelling of Glycated Collagen
3. Discussion
4. Materials and Methods
4.1. Collagen Procedures
4.1.1. Collagen Preparation
4.1.2. Fluorescent Labelling of Collagen
4.1.3. Preparation of Glycated Collagen
4.2. TNBS Method for Quantifying Free Amino Groups
4.3. Cells
4.3.1. Morphological Studies
4.3.2. Overall Cells Morphology and Focal Adhesion Formation
4.3.3. Quantitative Morphometry Analysis of Raw Format Images by ImageJ
4.3.4. Measurement of Cell Survival
4.3.5. Measurement of Collagen Degradation by ADMSC
4.4. FITC-Collagen Degradation in Cell-Free System
4.5. DSC Measurements
4.6. Statistical Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
References
- Bächinger, H.P.; Mizuno, K.; Vranka, J.A.; Boudko, S.P. Collagen Formation and Structure. Compr. Nat. Prod. II 2010, 5, 469–530. [Google Scholar] [CrossRef]
- Jiang, N.; Tian, X.; Wang, Q.; Hao, J.; Jiang, J.; Wang, H. Regulation Mechanisms and Maintenance Strategies of Stemness in Mesenchymal Stem Cells. Stem Cell Rev. Rep. 2024, 20, 455–483. [Google Scholar] [CrossRef] [PubMed]
- Rashid, U.; Yousaf, A.; Yaqoob, M.; Saba, E.; Moaeen-Ud-Din, M.; Waseem, S.; Becker, S.K.; Sponder, G.; Aschenbach, J.R.; Sandhu, M.A. Characterization and Differentiation Potential of Mesenchymal Stem Cells Isolated from Multiple Canine Adipose Tissue Sources. BMC Vet. Res. 2021, 17, 388. [Google Scholar] [CrossRef] [PubMed]
- Baglio, S.R.; Pegtel, D.M.; Baldini, N. Mesenchymal Stem Cell Secreted Vesicles Provide Novel Opportunities in (Stem) Cell-Free Therapy. Front. Physiol. 2012, 3, 359. [Google Scholar] [CrossRef] [PubMed]
- Kolf, C.M.; Cho, E.; Tuan, R.S. Mesenchymal Stromal Cells: Biology of Adult Mesenchymal Stem Cells: Regulation of Niche, Self-Renewal and Differentiation. Arthritis Res. Ther. 2007, 9, 204. [Google Scholar] [CrossRef]
- Rennert, R.C.; Sorkin, M.; Garg, R.K.; Gurtner, G.C. Stem Cell Recruitment after Injury: Lessons for Regenerative Medicine. Regen. Med. 2012, 7, 833–850. [Google Scholar] [CrossRef] [PubMed]
- Ku, C.; Johnson, P.; Batten, P.; Sarathchandra, P.; Chambers, R.; Taylor, P.; Yacoub, M.; Chester, A. Collagen Synthesis by Mesenchymal Stem Cells and Aortic Valve Interstitial Cells in Response to Mechanical Stretch. Cardiovasc. Res. 2006, 71, 548–556. [Google Scholar] [CrossRef]
- Alt, E.; Yan, Y.; Gehmert, S.; Song, Y.-H.; Altman, A.; Gehmert, S.; Vykoukal, D.; Bai, X. Fibroblasts Share Mesenchymal Phenotypes with Stem Cells, but Lack Their Differentiation and Colony-Forming Potential. Biol. Cell 2011, 103, 197–208. [Google Scholar] [CrossRef]
- Rogers, J.D.; Yeganegi, A.; Richardson, W.J. Mechano-Regulation of Fibrillar Collagen Turnover by Fibroblasts. In Mechanobiology Handbook, 2nd ed.; CRC Press: Boca Raton, FL, USA, 2018; ISBN 978-0-429-44498-2. [Google Scholar]
- Capobianco, E.; White, V.; Sosa, M.; Di Marco, I.; Basualdo, M.N.; Faingold, M.C.; Jawerbaum, A. Regulation of Matrix Metalloproteinases 2 and 9 Activities by Peroxynitrites in Term Placentas From Type 2 Diabetic Patients. Reprod. Sci. 2012, 19, 814–822. [Google Scholar] [CrossRef]
- Almalki, S.G.; Agrawal, D.K. Effects of Matrix Metalloproteinases on the Fate of Mesenchymal Stem Cells. Stem Cell Res. Ther. 2016, 7, 129. [Google Scholar] [CrossRef]
- Wu, M.; Cronin, K.; Crane, J.S. Biochemistry, Collagen Synthesis. In StatPearls; StatPearls Publishing: Treasure Island, FL, USA, 2024. [Google Scholar]
- Yamauchi, M.; Sricholpech, M. Lysine Post-Translational Modifications of Collagen. Essays Biochem. 2012, 52, 113–133. [Google Scholar] [CrossRef] [PubMed]
- Hennet, T. Collagen Glycosylation. Curr. Opin. Struct. Biol. 2019, 56, 131–138. [Google Scholar] [CrossRef] [PubMed]
- Rabbani, N.; Thornalley, P.J. Protein Glycation—Biomarkers of Metabolic Dysfunction and Early-Stage Decline in Health in the Era of Precision Medicine. Redox Biol. 2021, 42, 101920. [Google Scholar] [CrossRef] [PubMed]
- Baumann, S.; Hennet, T. Collagen Accumulation in Osteosarcoma Cells Lacking GLT25D1 Collagen Galactosyltransferase. J. Biol. Chem. 2016, 291, 18514–18524. [Google Scholar] [CrossRef] [PubMed]
- Perrone, A.; Giovino, A.; Benny, J.; Martinelli, F. Advanced Glycation End Products (AGEs): Biochemistry, Signaling, Analytical Methods, and Epigenetic Effects. Oxid. Med. Cell. Longev. 2020, 2020, 3818196. [Google Scholar] [CrossRef] [PubMed]
- Li, R.; Rajan, R.; Wong, W.C.V.; Reid, D.G.; Duer, M.J.; Somovilla, V.J.; Martinez-Saez, N.; Bernardes, G.J.L.; Hayward, R.; Shanahan, C.M. In Situ Characterization of Advanced Glycation End Products (AGEs) in Collagen and Model Extracellular Matrix by Solid State NMR. Chem. Commun. 2017, 53, 13316–13319. [Google Scholar] [CrossRef] [PubMed]
- Farhat, A.; Ferns, G.A.; Ashrafi, K.; Arjmand, M.-H. Lysyl Oxidase Mechanisms to Mediate Gastrointestinal Cancer Progression. Gastrointest. Tumors 2021, 8, 33–40. [Google Scholar] [CrossRef] [PubMed]
- Revell, C.K.; Jensen, O.E.; Shearer, T.; Lu, Y.; Holmes, D.F.; Kadler, K.E. Collagen Fibril Assembly: New Approaches to Unanswered Questions. Matrix Biol. Plus 2021, 12, 100079. [Google Scholar] [CrossRef] [PubMed]
- Komsa-Penkova, R.; Stavreva, G.; Belemezova, K.; Kyurkchiev, S.; Todinova, S.; Altankov, G. Mesenchymal Stem-Cell Remodeling of Adsorbed Type-I Collagen-The Effect of Collagen Oxidation. Int. J. Mol. Sci. 2022, 23, 3058. [Google Scholar] [CrossRef]
- Magin, C.M.; Alge, D.L.; Anseth, K.S. Bio-Inspired 3D Microenvironments: A New Dimension in Tissue Engineering. Biomed. Mater. 2016, 11, 022001. [Google Scholar] [CrossRef]
- Mikulíková, K.; Eckhardt, A.; Pataridis, S.; Mikšík, I. Study of Posttranslational Non-Enzymatic Modifications of Collagen Using Capillary Electrophoresis/Mass Spectrometry and High Performance Liquid Chromatography/Mass Spectrometry. J. Chromatogr. A 2007, 1155, 125–133. [Google Scholar] [CrossRef] [PubMed]
- Doyle, A.D. Fluorescent Labeling of Rat-Tail Collagen for 3D Fluorescence Imaging. Bio-Protocol 2018, 8, e2919. [Google Scholar] [CrossRef] [PubMed]
- de Almeida, L.G.N.; Thode, H.; Eslambolchi, Y.; Chopra, S.; Young, D.; Gill, S.; Devel, L.; Dufour, A. Matrix Metalloproteinases: From Molecular Mechanisms to Physiology, Pathophysiology, and Pharmacology. Pharmacol. Rev. 2022, 74, 714–770. [Google Scholar] [CrossRef] [PubMed]
- Avery, N.C.; Bailey, A.J. The Effects of the Maillard Reaction on the Physical Properties and Cell Interactions of Collagen. Pathol. Biol. 2006, 54, 387–395. [Google Scholar] [CrossRef] [PubMed]
- Fedintsev, A.; Moskalev, A. Stochastic Non-Enzymatic Modification of Long-Lived Macromolecules—A Missing Hallmark of Aging. Ageing Res. Rev. 2020, 62, 101097. [Google Scholar] [CrossRef] [PubMed]
- Evens, L.; Beliën, H.; Deluyker, D.; Bronckaers, A.; Gervois, P.; Hendrikx, M.; Bito, V. The Impact of Advanced Glycation End-Products (AGEs) on Proliferation and Apoptosis of Primary Stem Cells: A Systematic Review. Stem Cells Int. 2020, 2020, 8886612. [Google Scholar] [CrossRef]
- Sharma, S.D.; Pandey, B.N.; Mishra, K.P.; Sivakami, S. Amadori Product and Age Formation during Nonenzymatic Glycosylation of Bovine Serum Albumin in Vitro. J. Biochem. Mol. Biol. Biophys. 2002, 6, 233–242. [Google Scholar]
- Robins, S.P.; Bailey, A.J. Age-Related Changes in Collagen: The Identification of Reducible Lysine-Carbohydrate Condensation Products. Biochem. Biophys. Res. Commun. 1972, 48, 76–84. [Google Scholar] [CrossRef]
- Cohen, M.P.; Ziyadeh, F.N. Amadori Glucose Adducts Modulate Mesangial Cell Growth and Collagen Gene Expression. Kidney Int. 1994, 45, 475–484. [Google Scholar] [CrossRef]
- Ahmed, N. Advanced Glycation Endproducts—Role in Pathology of Diabetic Complications. Diabetes Res. Clin. Pract. 2005, 67, 3–21. [Google Scholar] [CrossRef]
- Reigle, K.L.; Lullo, G.D.; Turner, K.R.; Last, J.A.; Chervoneva, I.; Birk, D.E.; Funderburgh, J.L.; Elrod, E.; Germann, M.W.; Surber, C.; et al. Non-Enzymatic Glycation of Type I Collagen Diminishes Collagen–Proteoglycan Binding and Weakens Cell Adhesion. J. Cell. Biochem. 2008, 104, 1684–1698. [Google Scholar] [CrossRef] [PubMed]
- Yan, S.F.; Ramasamy, R.; Naka, Y.; Schmidt, A.M. Glycation, Inflammation, and RAGE. Circ. Res. 2003, 93, 1159–1169. [Google Scholar] [CrossRef] [PubMed]
- Singh, R.; Barden, A.; Mori, T.; Beilin, L. Advanced Glycation End-Products: A Review. Diabetologia 2001, 44, 129–146. [Google Scholar] [CrossRef] [PubMed]
- Dariya, B.; Nagaraju, G.P. Advanced Glycation End Products in Diabetes, Cancer and Phytochemical Therapy. Drug Discov. Today 2020, 25, 1614–1623. [Google Scholar] [CrossRef] [PubMed]
- Kawano, E.; Takahashi, S.-I.; Sakano, Y.; Fujimoto, D. Nonenzymatic Glycation Alters Properties of Collagen as a Substratum for Cells. Matrix 1990, 10, 300–305. [Google Scholar] [CrossRef] [PubMed]
- Verzijl, N.; DeGroot, J.; Zaken, C.B.; Braun-Benjamin, O.; Maroudas, A.; Bank, R.A.; Mizrahi, J.; Schalkwijk, C.G.; Thorpe, S.R.; Baynes, J.W.; et al. Crosslinking by Advanced Glycation End Products Increases the Stiffness of the Collagen Network in Human Articular Cartilage: A Possible Mechanism through Which Age Is a Risk Factor for Osteoarthritis. Arthritis Rheum. 2002, 46, 114–123. [Google Scholar] [CrossRef] [PubMed]
- Said, G.; Guilbert, M.; Millerot-Serrurot, E.; Van Gulick, L.; Terryn, C.; Garnotel, R.; Jeannesson, P. Impact of Carbamylation and Glycation of Collagen Type I on Migration of HT1080 Human Fibrosarcoma Cells. Int. J. Oncol. 2012, 40, 1797–1804. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Brodsky, S.V.; Goligorsky, D.M.; Hampel, D.J.; Li, H.; Gross, S.S.; Goligorsky, M.S. Glycated Collagen I Induces Premature Senescence-Like Phenotypic Changes in Endothelial Cells. Circ. Res. 2002, 90, 1290–1298. [Google Scholar] [CrossRef]
- Jost, T.; Zipprich, A.; Glomb, M.A. Analysis of Advanced Glycation Endproducts in Rat Tail Collagen and Correlation to Tendon Stiffening. J. Agric. Food Chem. 2018, 66, 3957–3965. [Google Scholar] [CrossRef]
- Fu, H.-L.; Valiathan, R.R.; Arkwright, R.; Sohail, A.; Mihai, C.; Kumarasiri, M.; Mahasenan, K.V.; Mobashery, S.; Huang, P.; Agarwal, G.; et al. Discoidin Domain Receptors: Unique Receptor Tyrosine Kinases in Collagen-Mediated Signaling. J. Biol. Chem. 2013, 288, 7430–7437. [Google Scholar] [CrossRef]
- Nørregaard, K.S.; Krigslund, O.; Behrendt, N.; Engelholm, L.H.; Jürgensen, H.J. The Collagen Receptor uPARAP/Endo180 Regulates Collectins through Unique Structural Elements in Its FNII Domain. J. Biol. Chem. 2020, 295, 9157–9170. [Google Scholar] [CrossRef]
- Olivares-Navarrete, R.; Rodil, S.E.; Hyzy, S.L.; Dunn, G.R.; Almaguer-Flores, A.; Schwartz, Z.; Boyan, B.D. Role of Integrin Subunits in Mesenchymal Stem Cell Differentiation and Osteoblast Maturation on Graphitic Carbon-Coated Microstructured Surfaces. Biomaterials 2015, 51, 69–79. [Google Scholar] [CrossRef] [PubMed]
- Heino, J. The Collagen Family Members as Cell Adhesion Proteins. BioEssays 2007, 29, 1001–1010. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.-M.; Käpylä, J.; Puranen, J.S.; Knight, C.G.; Tiger, C.-F.; Pentikäinen, O.T.; Johnson, M.S.; Farndale, R.W.; Heino, J.; Gullberg, D. A11β1 Integrin Recognizes the GFOGER Sequence in Interstitial Collagens. J. Biol. Chem. 2003, 278, 7270–7277. [Google Scholar] [CrossRef] [PubMed]
- Ludwig, B.S.; Kessler, H.; Kossatz, S.; Reuning, U. RGD-Binding Integrins Revisited: How Recently Discovered Functions and Novel Synthetic Ligands (Re-)Shape an Ever-Evolving Field. Cancers 2021, 13, 1711. [Google Scholar] [CrossRef] [PubMed]
- Bansode, S.; Bashtanova, U.; Li, R.; Clark, J.; Müller, K.H.; Puszkarska, A.; Goldberga, I.; Chetwood, H.H.; Reid, D.G.; Colwell, L.J.; et al. Glycation Changes Molecular Organization and Charge Distribution in Type I Collagen Fibrils. Sci. Rep. 2020, 10, 3397. [Google Scholar] [CrossRef] [PubMed]
- Rodrigues, M.; Griffith, L.G.; Wells, A. Growth Factor Regulation of Proliferation and Survival of Multipotential Stromal Cells. Stem Cell Res. Ther. 2010, 1, 32. [Google Scholar] [CrossRef] [PubMed]
- Hidalgo, A.; Sanz-Rodríguez, F.; Rodríguez-Fernández, J.L.; Albella, B.; Blaya, C.; Wright, N.; Cabañas, C.; Prósper, F.; Gutierrez-Ramos, J.C.; Teixidó, J. Chemokine Stromal Cell-Derived Factor-1α Modulates VLA-4 Integrin-Dependent Adhesion to Fibronectin and VCAM-1 on Bone Marrow Hematopoietic Progenitor Cells. Exp. Hematol. 2001, 29, 345–355. [Google Scholar] [CrossRef] [PubMed]
- Komsa-Penkova, R.; Stoycheva, S.; Tonchev, P.; Stavreva, G.; Todinova, S.; Georgieva, G.; Yordanova, A.; Kyurkchiev, S.; Altankov, G. Morphological and Quantitative Evidence for Altered Mesenchymal Stem Cell Remodeling of Collagen in an Oxidative Environment-Peculiar Effect of Epigallocatechin-3-Gallate. Polymers 2022, 14, 3957. [Google Scholar] [CrossRef]
- Carolina Florian, M.; Geiger, H. Concise Review: Polarity in Stem Cells, Disease, and Aging. Stem Cells 2010, 28, 1623–1629. [Google Scholar] [CrossRef]
- Komsa-Penkova, R.; Spirova, R.; Bechev, B. Modification of Lowry’s Method for Collagen Concentration Measurement. J. Biochem. Biophys. Methods 1996, 32, 33–43. [Google Scholar] [CrossRef]
- Kuznetsova, N.; Leikin, S. Does the Triple Helical Domain of Type I Collagen Encode Molecular Recognition and Fiber Assembly While Telopeptides Serve as Catalytic Domains? J. Biol. Chem. 1999, 274, 36083–36088. [Google Scholar] [CrossRef] [PubMed]
- Available online: https://www.sigmaaldrich.com/deepweb/assets/sigmaaldrich/product/documents/222/544/f7250pis.pdf (accessed on 20 December 2023).
- Dandia, H.; Makkad, K.; Tayalia, P. Glycated Collagen—A 3D Matrix System to Study Pathological Cell Behavior. Biomater. Sci. 2019, 7, 3480–3488. [Google Scholar] [CrossRef] [PubMed]
- Bubnis, W.A.; Ofner, C.M. The Determination of ϵ-Amino Groups in Soluble and Poorly Soluble Proteinaceous Materials by a Spectrophotometric Method Using Trinitrobenzenesulfonic Acid. Anal. Biochem. 1992, 207, 129–133. [Google Scholar] [CrossRef] [PubMed]
- Boudaoud, A.; Burian, A.; Borowska-Wykręt, D.; Uyttewaal, M.; Wrzalik, R.; Kwiatkowska, D.; Hamant, O. FibrilTool, an ImageJ Plug-in to Quantify Fibrillar Structures in Raw Microscopy Images. Nat. Protoc. 2014, 9, 457–463. [Google Scholar] [CrossRef]
- Jedeszko, C.; Sameni, M.; Olive, M.B.; Moin, K.; Sloane, B.F. Visualizing Protease Activity in Living Cells: From Two Dimensions to Four Dimensions. Curr. Protoc. Cell Biol. 2008, 39, 4–20. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample | Tm (°C) | ∆Hcal (cal g−1) | cPex (cal.g−1 K−1) | Tm ½ (°C) |
|---|---|---|---|---|
| RTC native | 40.4 | 8.76 | 4.93 | 1.57 |
| RTC GL1 (1 day glycated) | 40.2 | 6.82 | 3.58 | 1.72 |
| RTC GL5 (5 days glycated) | 40.5 | 6.98 | 3.67 | 1.71 |
| Parameter | Incubation Time | RTC | RTC-GL1 | RTC-GL5 |
|---|---|---|---|---|
| CSA (μm2) | 5 h | 246 ± 143 (n = 45) *a | 217 ± 146 (n = 42) | 164 ± 109 (n = 40) *a |
| 24 h | 249 ± 150 (n = 34) | 211 ± 171 (n = 32) | 226 ± 123 (n = 20) | |
| Perimeter (μm) | 5 h | 113 ± 56 (n = 30) *b | 95 ± 40 (n = 42) | 86 ± 42 (n = 40) *b |
| 24 h | 130 ± 54 (n = 34) *c | 72 ± 5 (n = 30) | 65 ± 4 (n = 30) *c | |
| CSI | 5 h | 0.30 ± 0.21 (n = 45) | 0.3 ± 0.2 (n = 45) | 0.3 ± 0.1 (n = 20) |
| 24 h | 0.21 ± 0.12 (n = 34) | 0.23 ± 0.17 (n = 34) *d | 0.14 ± 0.11 (n = 20) *d | |
| CAR | 5 h | 1.5 ± 0.4 (n = 45) *e | 1.9 ± 0.9 (n = 42) *e | 1.7 ± 0.6 (n = 40) |
| 24 h | 3.4 ± 0.23 (n = 34) | 3.6 ± 2.5 (n = 32) | 3.9 ± 2.7 (n = 20) |
| Incubation Time in Hours | 2 h | 5 h | 24 h |
|---|---|---|---|
| Viability (%) (Number of viable cells) | Live Cells | Live Cells | Live Cells |
| RTC native | 94.2 (114) | 95.4 (165) | 96.2 (230) |
| RTC GL1 | 88.4 (122) | 94.8 (251) | 95.1 (330) |
| RTC GL5 | 74.6 (94) | 94.7 (270) | 93.3 (252) |
| TC Polystyrene | 80.5 (124) | 88.5 (193) | 94.5 (329) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Komsa-Penkova, R.; Dimitrov, B.; Todinova, S.; Ivanova, V.; Stoycheva, S.; Temnishki, P.; Georgieva, G.; Tonchev, P.; Iliev, M.; Altankov, G. Early Stages of Ex Vivo Collagen Glycation Disrupt the Cellular Interaction and Its Remodeling by Mesenchymal Stem Cells—Morphological and Biochemical Evidence. Int. J. Mol. Sci. 2024, 25, 5795. https://doi.org/10.3390/ijms25115795
Komsa-Penkova R, Dimitrov B, Todinova S, Ivanova V, Stoycheva S, Temnishki P, Georgieva G, Tonchev P, Iliev M, Altankov G. Early Stages of Ex Vivo Collagen Glycation Disrupt the Cellular Interaction and Its Remodeling by Mesenchymal Stem Cells—Morphological and Biochemical Evidence. International Journal of Molecular Sciences. 2024; 25(11):5795. https://doi.org/10.3390/ijms25115795
Chicago/Turabian StyleKomsa-Penkova, Regina, Borislav Dimitrov, Svetla Todinova, Violina Ivanova, Svetoslava Stoycheva, Peter Temnishki, Galya Georgieva, Pencho Tonchev, Mario Iliev, and George Altankov. 2024. "Early Stages of Ex Vivo Collagen Glycation Disrupt the Cellular Interaction and Its Remodeling by Mesenchymal Stem Cells—Morphological and Biochemical Evidence" International Journal of Molecular Sciences 25, no. 11: 5795. https://doi.org/10.3390/ijms25115795