
Transglutaminase-Cross-Linked Tofu Suppressed Soybean-Induced Allergic Reactions by Enhancing Intestinal Mucosa Immune Tolerance
 , , ,
, , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Preparation of Samples
2.3. Mice
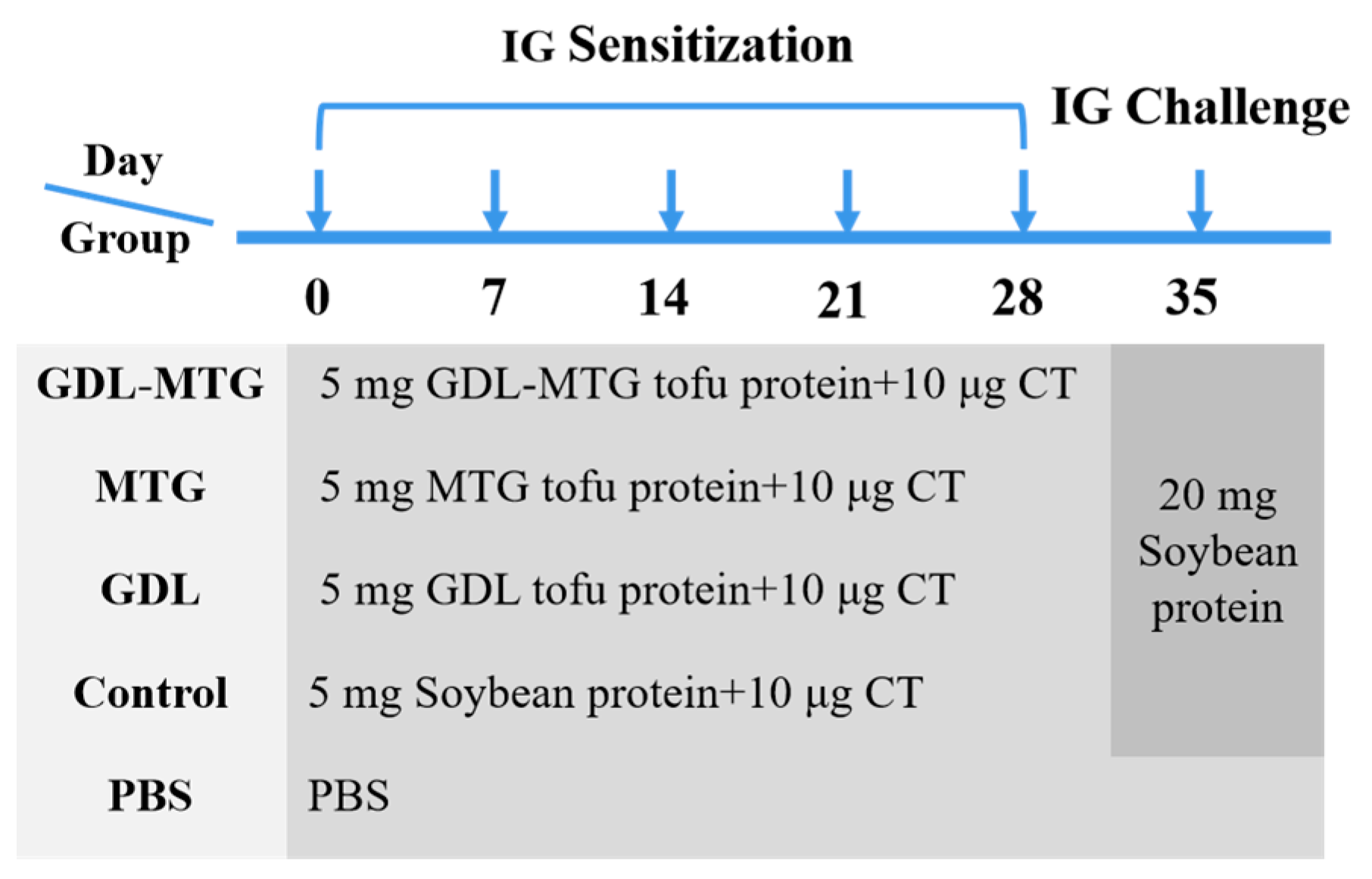
2.4. Experimental Design
2.5. Analysis of Immunoglobulin E (IgE)
2.6. Preparation of PP and MLN Cell Suspension
2.7. Flow Cytometric Analysis of Immune Cells
2.7.1. Detection of DCs
2.7.2. Detection of T Lymphocyte Subsets
2.8. HE
2.9. Statistical Analysis
3. Results
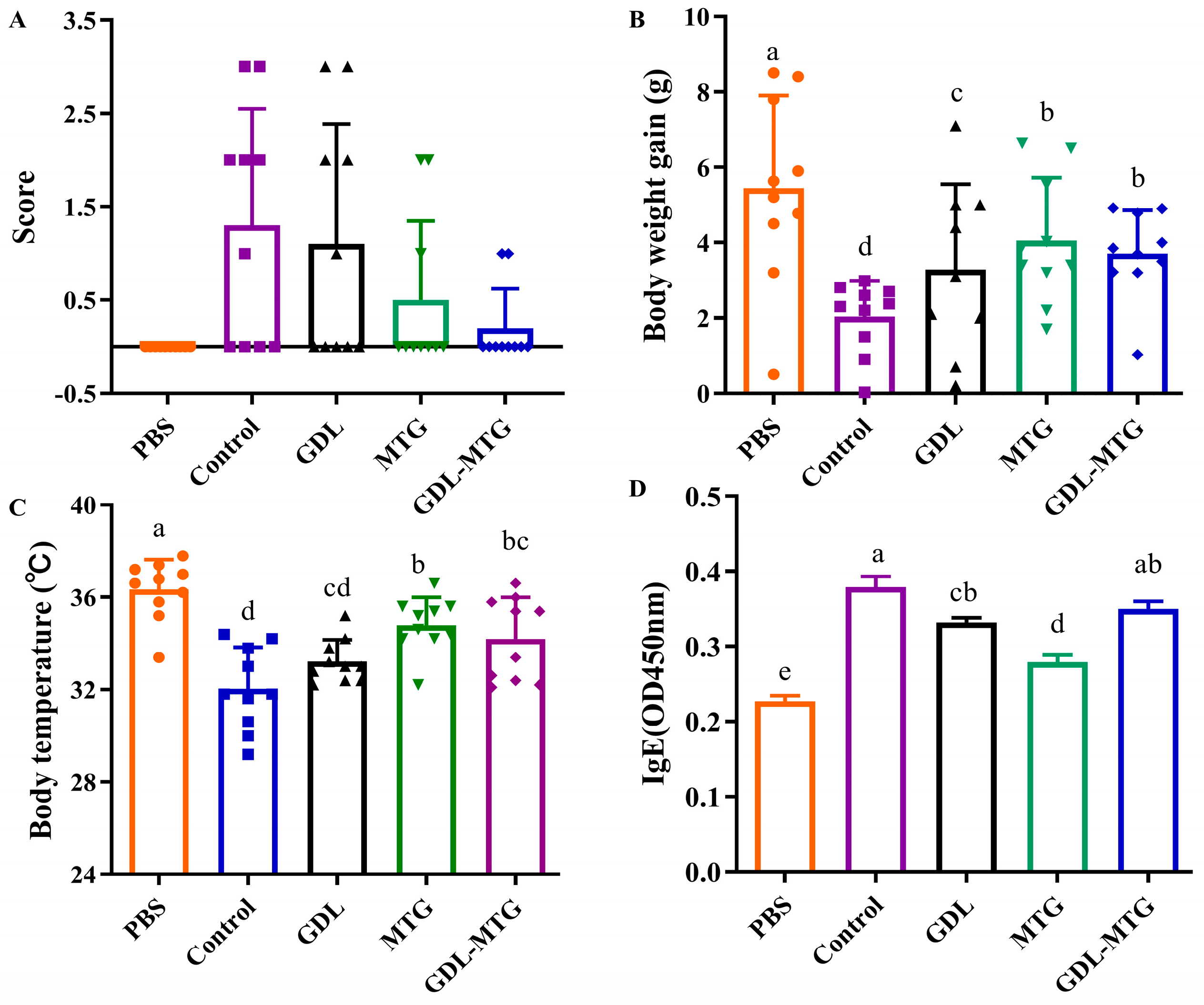
3.1. Evaluation of Allergic Mouse Model
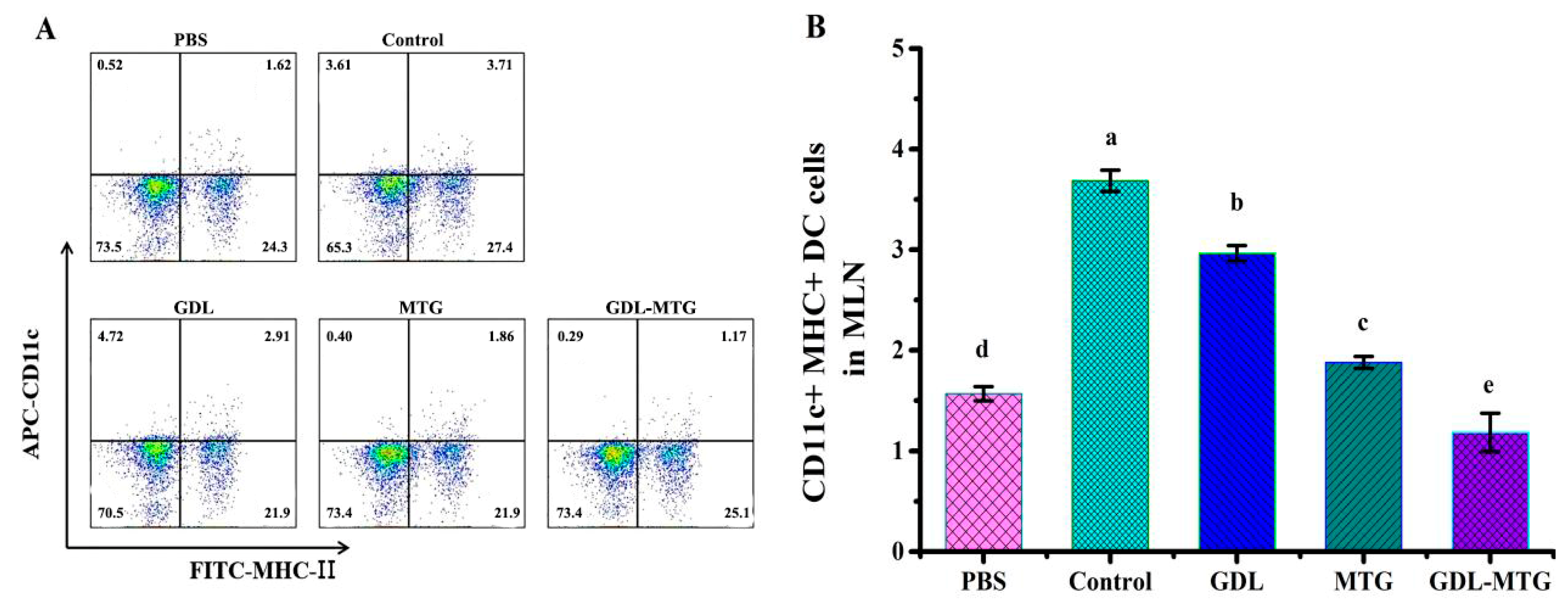
3.2. Role of DC Presentation
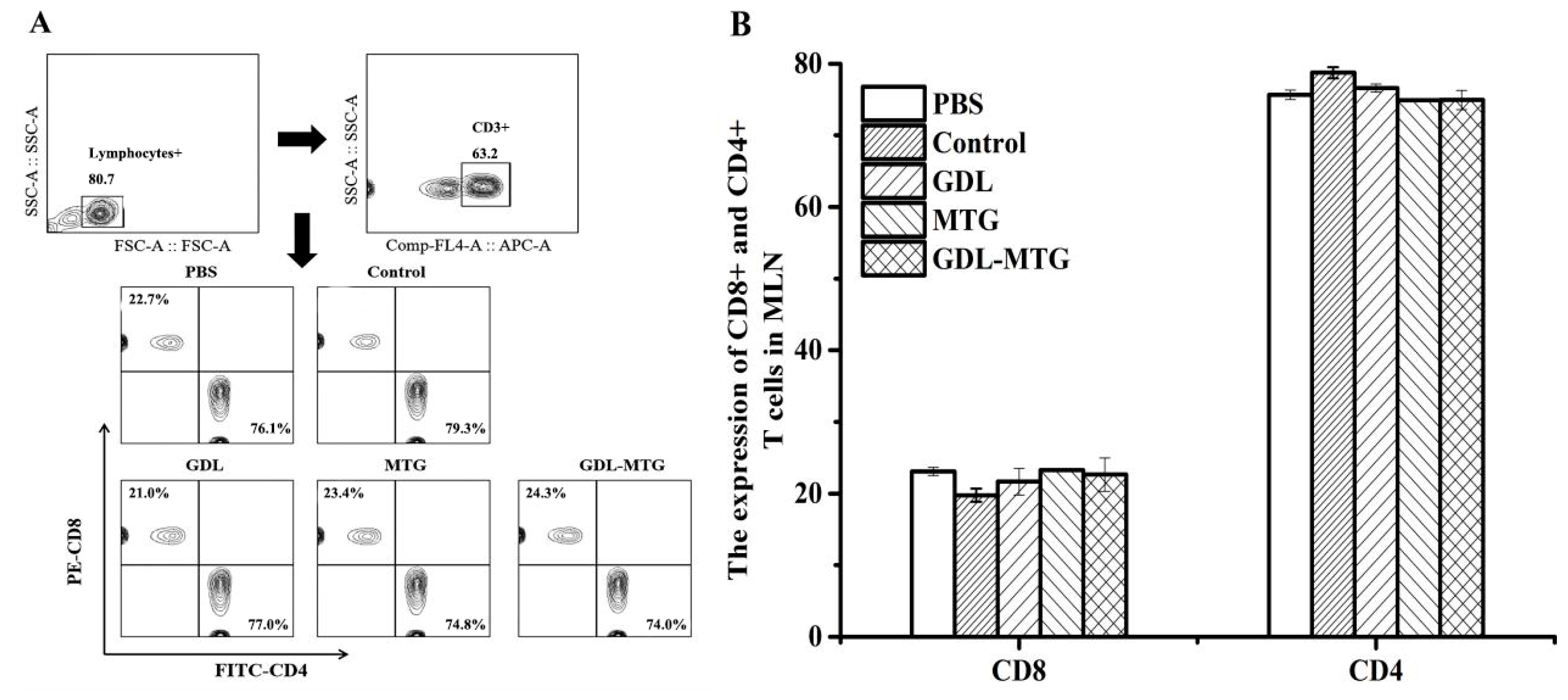
3.3. Detection of CD4/CD8 Cells
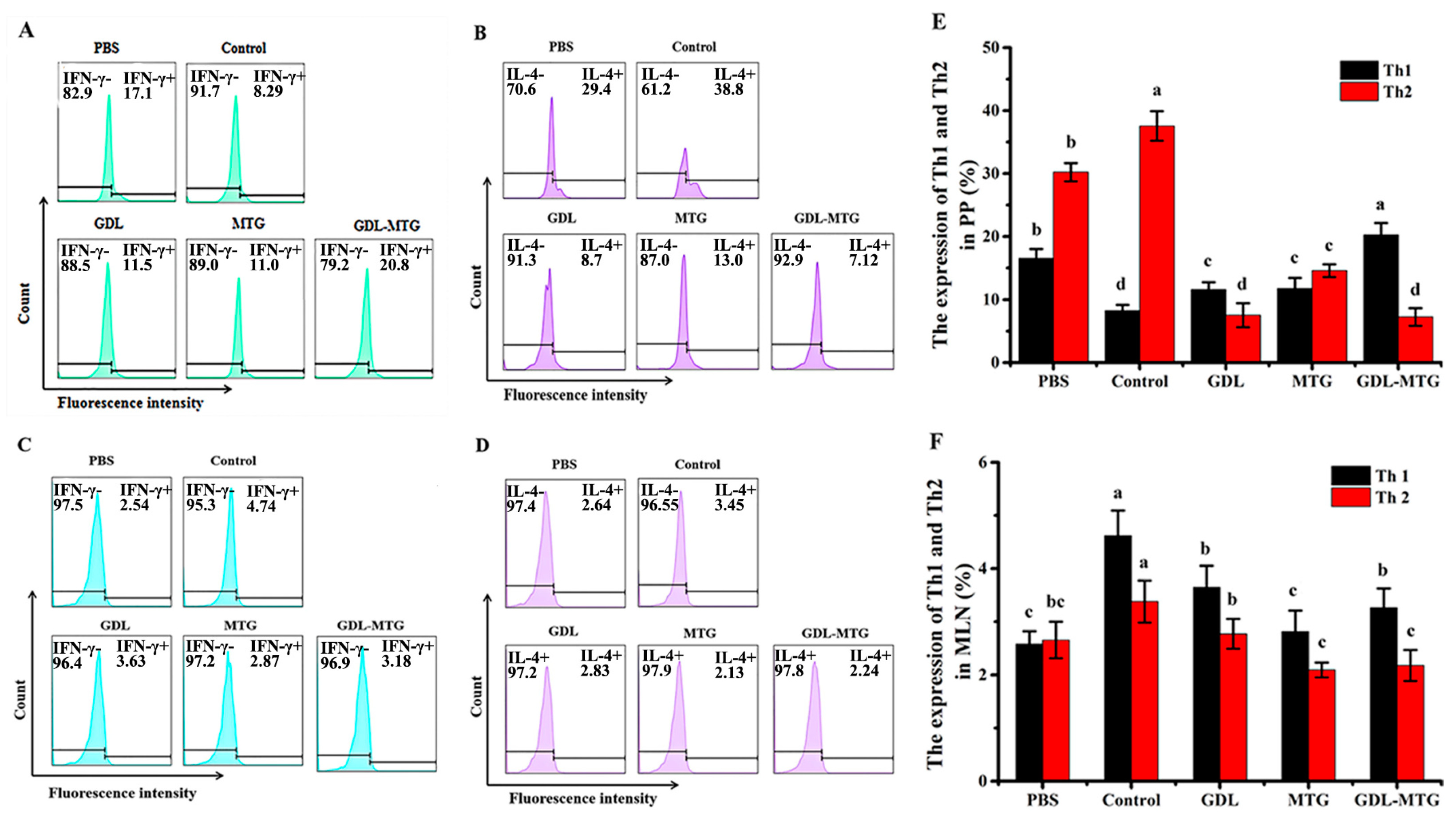
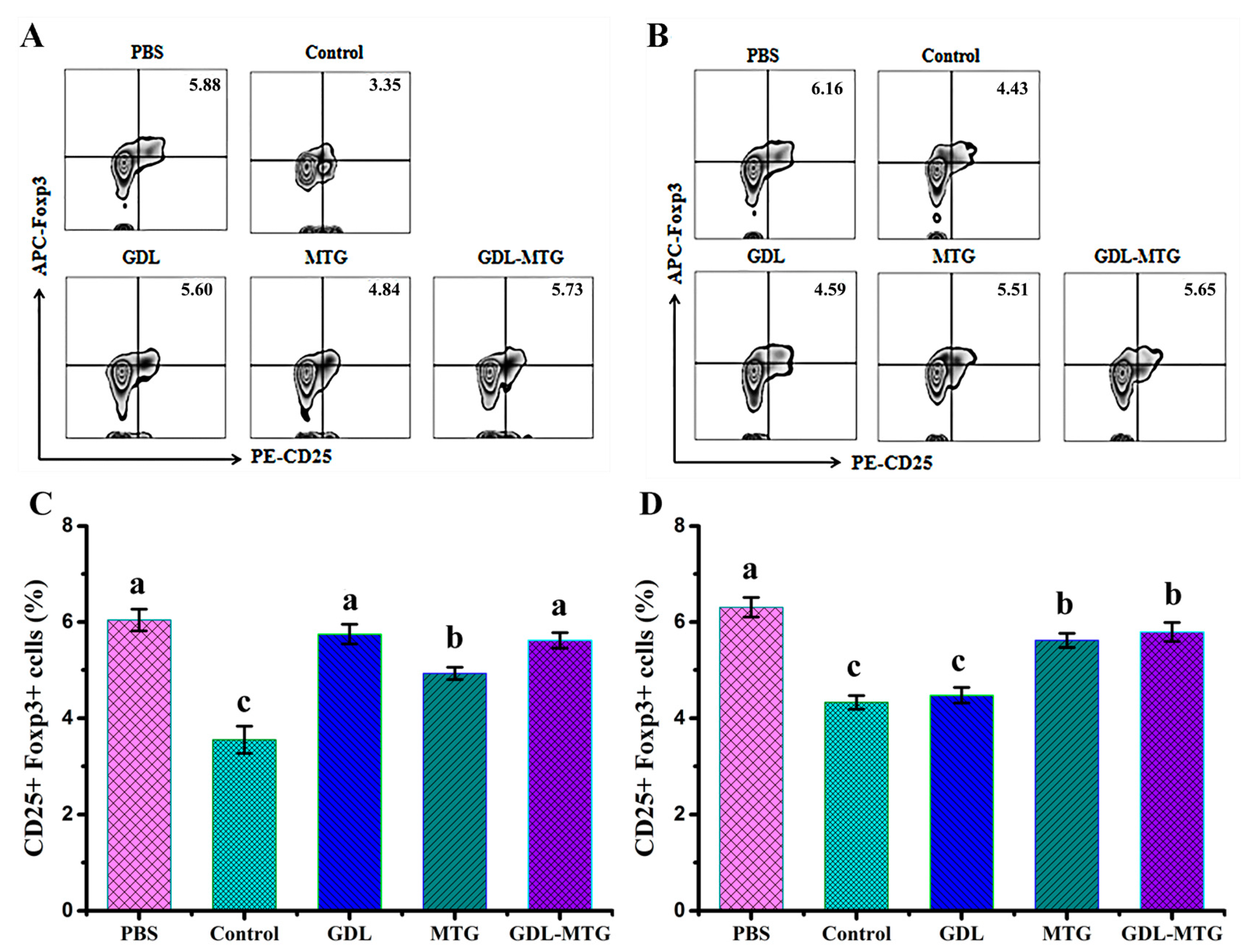
3.4. Identification of T Lymphocyte Subsets
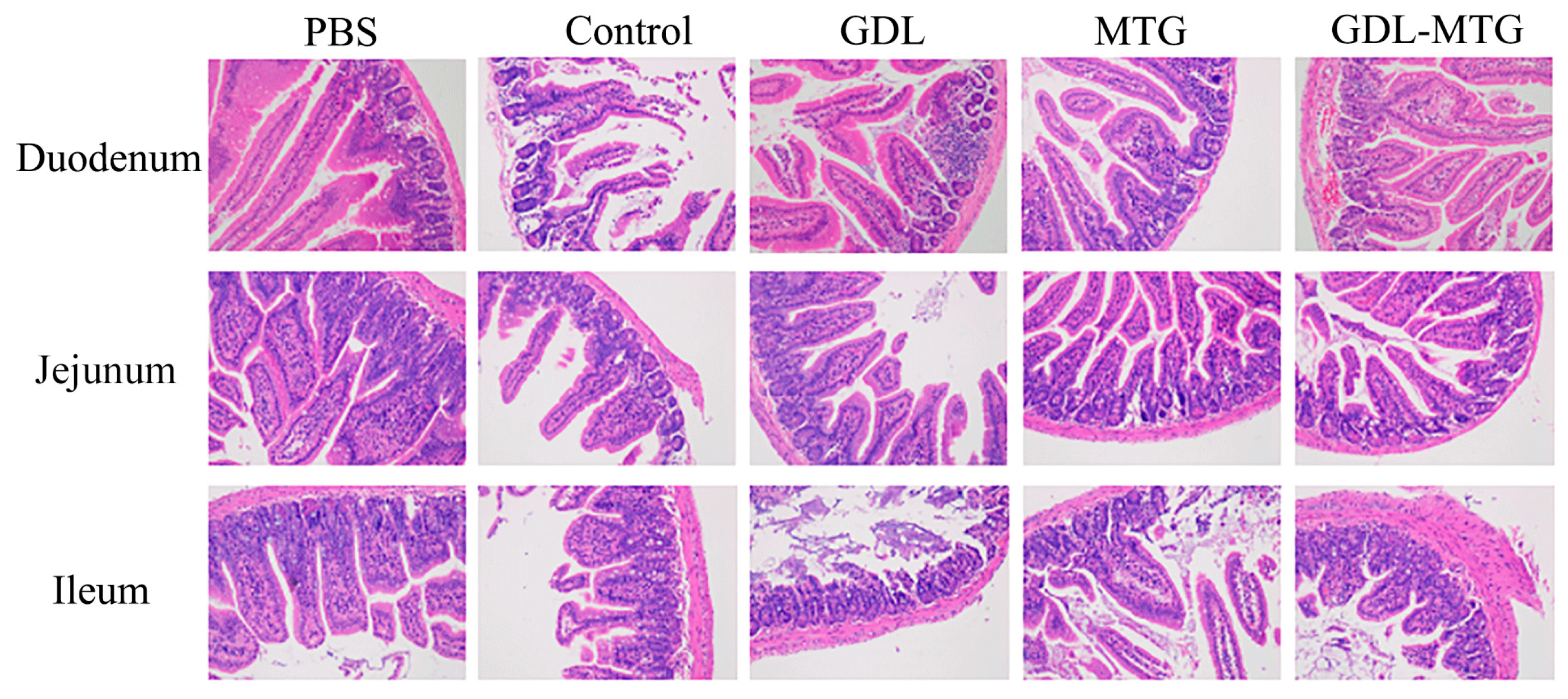
3.5. Histopathological Section of the Intestine
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Rossi, F.; Felis, G.E.; Martinelli, A.; Calcavecchia, B.; Torriani, S. Microbiological characteristics of fresh tofu produced in small industrial scale and identification of specific spoiling microorganisms (SSO). LWT-Food Sci. Technol. 2016, 70, 280–285. [Google Scholar] [CrossRef]
- Chen, X.; Zhao, X.; Hu, Y.; Bowei, Z.; Zhang, Y.; Wang, S. Lactobacillus rhamnosus GG alleviates β-conglycinin-induced allergy by regulating the T cell receptor signaling pathway. Food Funct. 2020, 11, 10554–10567. [Google Scholar] [CrossRef] [PubMed]
- Fiocchi, A.; Burks, W.; Bahna, S.L.; Bielory, L.; Boyle, R.J.; Cocco, R.; Dreborg, S.; Goodman, R.; Kuitunen, M.; Haahtela, T.; et al. Clinical Use of Probiotics in Pediatric Allergy (CUPPA): A World Allergy Organization Position Paper. World Allergy Organ J. 2012, 5, 148–167. [Google Scholar] [CrossRef] [PubMed]
- Onodera, Y.; Ono, T.; Nakasato, K.; Toda, K. Homogeneity and microstructure of tofu depends on 11S/7S globulin ratio in soymilk and coagulant concentration. Food Sci. Technol. Res. 2009, 15, 265–274. [Google Scholar] [CrossRef]
- James, A.T.; Yang, A. Interactions of protein content and globulin subunit composition of soybean proteins in relation to tofu gel properties. Food Chem. 2016, 194, 284–289. [Google Scholar] [CrossRef] [PubMed]
- Tang, C.; Luo, L.; Liu, F.; Chen, Z. Transglutaminase-set soy globulin-stabilized emulsion gels: Influence of soy β-conglycinin/glycinin ratio on properties, microstructure and gelling mechanism. Food Res. Int. 2013, 51, 804–812. [Google Scholar] [CrossRef]
- Chang, Y.; Shiau, S.; Chen, F.; Lin, F. Effect of microbial transglutaminase on the rheological and textural characteristics of black soybean packed tofu coagulating with Agar. LWT-Food Sci. Technol. 2011, 44, 1107–1112. [Google Scholar] [CrossRef]
- Yasir, S.; Sutton, K.; Newberry, M.; Andrews, N.; Gerrard, J. The impact of transglutaminase on soy proteins and tofu texture. Food Chem. 2007, 104, 1491–1501. [Google Scholar] [CrossRef]
- Xing, G.; Giosafatto, C.V.L.; Rui, X.; Dong, M.; Mariniello, L. Microbial transglutaminase-mediated polymerization in the presence of lactic acid bacteria affects antigenicity of soy protein component present in bio-tofu. J. Funct. Foods 2019, 53, 292–298. [Google Scholar] [CrossRef]
- Adachi, A.; Horikawa, T.; Shimizu, H.; Sarayama, Y.; Ogawa, T.; Sjolander, S.; Tanaka, A.; Moriyama, T. Soybean β-conglycinin as the main allergen in a patient with food dependent exerciseinduced anaphylaxis bytofu: Food processing alters pepsin resistance. Clin. Exp. Allergy 2009, 39, 167–173. [Google Scholar] [CrossRef]
- Bai, J.; Zhao, X.; Zhang, M.; Xia, X.; Yang, A.; Chen, H. Gut microbiota: A target for prebiotics and probiotics in the intervention and therapy of food allergy. Crit. Rev. Food Sci. 2022, 64, 3623–3637. [Google Scholar] [CrossRef] [PubMed]
- Cerutti, A.; Chen, K.; Chorny, A.A. Immunoglobulin responses at the mucosal interface. Annu. Rev. Immunol. 2011, 29, 273–293. [Google Scholar] [CrossRef] [PubMed]
- Caubet, J.; Szajewska, H.; Shamir, R.; Nowak-Węgrzyn, A. Non-IgE-mediated gastrointestinal food allergies in children. Pediat. Allerg. Imm. 2017, 28, 6–17. [Google Scholar] [CrossRef]
- Tsuda, M.; Arakawa, H.; Ishii, N.; Ubukata, C.; Michimori, M.; Noda, M.; Takahashi, K.; Kaminogawa, S.; Hosono, A. Dietary Fructo-Oligosaccharides Attenuate Early Activation of CD4+ T Cells Which Produce both Th1 and Th2 Cytokines in the Intestinal Lymphoid Tissues of a Murine Food Allergy Model. Int. Arch. Allergy Imm. 2017, 174, 121–132. [Google Scholar] [CrossRef] [PubMed]
- Tordesillas, L.; Mondoulet, L.; Blazquez, A.B.; Benhamou, P.H.; Sampson, H.A.; Berin, C. Epicutaneous immunotherapy induces gastrointestinal LAP+ Tregs and prevents food-induced anaphylaxis. J. Allergy Clin Immun. 2017, 139, 189–201. [Google Scholar] [CrossRef] [PubMed]
- Meng, X.; Wu, Y.; Wen, X.; Gao, J.; Xie, Y.; Zhao, X.; Yuan, J.; Yang, H.; Zeng, Z.; Li, X.; et al. Dietary Linolenic Acid Increases Sensitizing and Eliciting Capacities of Cow’s Milk Whey Proteins in BALB/c Mice. Nutrients 2022, 14, 822. [Google Scholar] [CrossRef] [PubMed]
- Villas-Boas, M.B.; Vieira, K.P.; Trevizan, G.; de Lima Zollner, R.; Netto, F.M. The effect of transglutaminase-induced polymerization in the presence of cysteine on β-lactoglobulin antigenicity. Int. Dairy J. 2010, 20, 386–392. [Google Scholar] [CrossRef]
- Porta, R.; Giosafatto, C.V.L.; di Pierro, P.; Sorrentino, A.; Mariniello, L. Transglutaminase-mediated modification of ovomucoid: Effects on its trypsin inhibitory activity and antigenic properties. Amino Acids 2013, 44, 285–292. [Google Scholar] [CrossRef]
- Yang, M.; Liu, F.; Tang, C. Properties and microstructure of transglutaminase-set soy protein-stabilized emulsion gels. Food Res. Int. 2013, 52, 409–418. [Google Scholar] [CrossRef]
- Meng, S.; Tan, Y.; Chang, S.; Li, J.; Maleki, S.; Puppala, N. Peanut allergen reduction and functional property improvement by means of enzymatic hydrolysis and transglutaminase crosslinking. Food Chem. 2020, 302, 125186. [Google Scholar] [CrossRef]
- Benedé, S.; Martínez-Blanco, M.; López-Fandiño, R.; Molina, E. IgE-Binding and Immunostimulating Properties of Enzymatic Crosslinked Milk Proteins as Influenced by Food Matrix and Digestibility. Nutrients 2022, 14, 4584. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, I.; Lin, H.; Li, Z.; Xu, L.; Qazi, I.M.; Luo, C.; Gao, X.; Khan, M.U.; Iqbal, A.; Guo, Y.; et al. Tyrosinase/caffeic acid cross-linking alleviated shrimp (Metapenaeus ensis) tropomyosin-induced allergic responses by modulating the Th1/Th2 immunobalance. Food Chem. 2021, 340, 127948. [Google Scholar] [CrossRef] [PubMed]
- Liu, G.; Hu, M.; Sun, L.; Han, X.; Liu, Q.; Alcocer, M.; Fei, D.; Cao, M.; Liu, G. Allergenicity and Oral Tolerance of Enzymatic Cross-Linked Tropomyosin Evaluated Using Cell and Mouse Models. J. Agr. Food Chem. 2017, 65, 2205–2213. [Google Scholar] [CrossRef] [PubMed]
- Bai, J.; Hui, J.; Lu, Q.; Yang, A.; Yuan, J.; Gao, J.; Wu, Z.; Li, X.; Tong, P.; Chen, H. Effect of transglutaminase cross-linking on the allergenicity of tofu based on a BALB/c mouse model. Food Funct. 2020, 11, 404–413. [Google Scholar] [CrossRef]
- Zhu, J.; Deng, H.; Yang, A.; Wu, Z.; Li, X.; Tong, P.; Chen, H. Effect of microbial transglutaminase cross-linking on the quality characteristics and potential allergenicity of tofu. Food Funct. 2019, 10, 5485–5497. [Google Scholar] [CrossRef] [PubMed]
- Yang, A.; Liao, Y.; Zhu, J.; Zhang, J.; Wu, Z.; Li, X.; Tong, P.; Chen, H.; Wang, S.; Liu, Z. Screening of anti-allergy Lactobacillus and its effect on allergic reactions in BALB/c mice sensitized by soybean protein. J. Funct. Foods 2021, 87, 104858. [Google Scholar] [CrossRef]
- Perez-Mazliah, D.; Langhorne, J. CD4 T-Cell Subsets in Malaria: TH1/TH2 Revisited. Front. Immunol. 2015, 5, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Talaat, R.M.; Mohamed, S.F.; Bassyouni, I.H.; Raouf, A.A. Th1/Th2/Th17/Treg cytokine imbalance in systemic lupus erythematosus (SLE) patients: Correlation with disease activity. Cytokine 2015, 72, 146–153. [Google Scholar] [CrossRef] [PubMed]
- Graham, M.T.; Nadeau, K.C. Lessons learned from mice and man: Mimicking human allergy through mouse models. Clin. Immunol. 2014, 155, 1–16. [Google Scholar] [CrossRef]
- Kanagaratham, C.; Sallis, B.F.; Fiebiger, E. Experimental Models for Studying Food Allergy. Cell. Mol. Gastroenter. 2018, 6, 356–369. [Google Scholar] [CrossRef]
- Schülke, S.; Albrecht, M. Mouse Models for Food Allergies: Where Do We Stand? Cells 2019, 8, 546. [Google Scholar] [CrossRef] [PubMed]
- Castan, L.; Bøgh, K.L.; Maryniak, N.Z.; Epstein, M.M.; Kazemi, S.; O’Mahony, L.; Bodinier, M.; Smit, J.J.; Bilsen, J.H.M.; Blanchard, C.; et al. Overview of in vivo and ex vivo endpoints in murine food allergy models: Suitable for evaluation of the sensitizing capacity of novel proteins? Allergy 2020, 75, 289–301. [Google Scholar] [CrossRef] [PubMed]
- Van Gramberg, J.L.; de Veer, M.J.; O’Hehir, R.E.; Meeusen, E.N.T.; Bischof, R.J. Use of Animal Models to Investigate Major Allergens Associated with Food Allergy. J. Allergy 2013, 2013, 635695. [Google Scholar] [CrossRef] [PubMed]
- Gao, J.; Li, T.; Liang, D.; Gong, H.; Zhao, L.; Mao, X. Maternal obesity exacerbates the responsiveness of offspring BALB/c mice to cow’s milk protein-induced food allergy. Food Sci. Hum. Well. 2023, 12, 920–928. [Google Scholar] [CrossRef]
- Krempski, J.W.; Lama, J.K.; Iijima, K.; Kobayashi, T.; Matsunaga, M.; Kita, H. A mouse model of the LEAP study reveals a role for CTLA-4 in preventing peanut allergy induced by environmental peanut exposure. J. Allergy Clin. Immun. 2022, 150, 425–439. [Google Scholar] [CrossRef] [PubMed]
- Deng, Y.; Chen, H.; Wu, Y.; Yuan, J.; Shi, Q.; Tong, P.; Gao, J. Aflatoxin B1 can aggravate BALB/c mice allergy to ovalbumin through changing their Th2 cells immune responses. Toxicon 2023, 228, 107121. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.; Chen, C.; Yan, X.; Gu, S.; Jia, X.; Fu, W.; Meng, X.; Xue, W. Assessment of immune responses and intestinal flora in BALB/c mice model of wheat food allergy via different sensitization methods. Food Sci. Hum. Well. 2023, 12, 871–881. [Google Scholar] [CrossRef]
- Wu, Y.; Lu, Y.; Huang, Y.; Lin, H.; Dang, X.; Jing, Y.; Meng, Z.; Wang, X.; Li, Z. Insight analysis of the cross-sensitization of multiple fish parvalbuminsvia the Th1/Th2 immunological balance and cytokine release from the perspective of safe consumption of fish. Food Qual. Saf. 2022, 6, 690–699. [Google Scholar]
- Smit, J.J.; Bol-Schoenmakers, M.; Hassing, I.; Fiechter, D.; Boon, L.; Bleumink, R.; Pieters, R.H.H. The role of intestinal dendritic cells subsets in the establishment of food allergy. Clin. Exp. Allergy 2011, 41, 890–898. [Google Scholar] [CrossRef]
- Blázquez, A.B.; Berin, M.C. Gastrointestinal Dendritic Cells Promote Th2 Skewing via OX40L. J. Immunol. 2008, 180, 4441–4450. [Google Scholar] [CrossRef]
- Samadi, N.; Klems, M.; Heiden, D.; Bauer, R.; Kitzmüller, C.; Weidmann, E.; Ret, D.; Ondracek, A.S.; Duschl, A.; Horejs Hoeck, J.; et al. Nitrated food proteins induce a regulatory immune response associated with allergy prevention after oral exposure in a Balb/c mouse food allergy model. Allergy 2020, 75, 412–422. [Google Scholar] [CrossRef]
- de Kivit, S.; Kostadinova, A.I.; Kerperien, J.; Morgan, M.E.; Muruzabal, V.A.; Hofman, G.A.; Knippels, L.M.J.; Kraneveld, A.D.; Garssen, J.; Willemsen, L.E.M. Dietary, nondigestible oligosaccharides and Bifidobacterium breve M-16V suppress allergic inflammation in intestine via targeting dendritic cell maturation. J. Leukoc. Biol. 2017, 102, 105–115. [Google Scholar] [CrossRef]
- Forchielli, M.L.; Walker, W.A. The role of gut-associated lymphoid tissues and mucosal defence. Brit. J. Nutr. 2005, 93, S41–S48. [Google Scholar] [CrossRef] [PubMed]
- Constantinescu, C.S.; Arsenescu, R.I.; Arsenescu, V. Targeting Immunomodulatory Agents to the Gut-Associated Lymphoid Tissue; Springer: Cham, Switzerland, 2016; pp. 237–261. [Google Scholar]
- Liu, P.; Kang, C.; Zhang, J.; Liu, Y.; Liu, J.; Hu, T.; Zeng, X.; Qiu, S. The role of dendritic cells in allergic diseases. Int. Immunopharmacol. 2022, 113, 109449. [Google Scholar] [CrossRef]
- Kumar, P.; Saini, S.; Khan, S.; Lele, S.S.; Prabhakar, B.S. Restoring Self-tolerance in Autoimmune Diseases by Enhancing Regulatory Tcell. Cell. Immunol. 2018, 339, 41–49. [Google Scholar] [CrossRef]
- Lee, K.; Park, M.; Ji, K.; Lee, H.; Jang, J.; Yoon, I.; Oh, S.; Kim, S.; Jeong, Y.; Yun, C.; et al. Bacterial β-(1,3)-glucan prevents DSS-induced IBD by restoring the reduced population of regulatory T cells. Immunobiology 2014, 219, 802–812. [Google Scholar] [CrossRef] [PubMed]
- Rivas, M.N.; Chatila, T.A. Regulatory T cells in allergic diseases. J. Allergy Clin. Immunol. 2017, 138, 639–652. [Google Scholar] [CrossRef] [PubMed]
- Niewiem, M.; Grzybowska-Chlebowczyk, U. Intestinal Barrier Permeability in Allergic Diseases. Nutrients 2022, 14, 1893. [Google Scholar] [CrossRef]
- Yang, A.; Zuo, L.; Cheng, Y.; Wu, Z.; Li, X.; Tong, P.; Chen, H. Degradation of major allergens and allergenicity reduction of soybean meal through solid-state fermentation with microorganisms. Food Funct. 2018, 9, 1899–1909. [Google Scholar] [CrossRef]
- Yang, H.; Gao, J.; Yang, A.; Lu, J.; Chen, H. Allergenicity characteristics of germinated soybean proteins in a BALB/c mouse model. Regul. Toxicol. Pharm. 2015, 72, 249–255. [Google Scholar] [CrossRef]
- Samadi, N.; Klems, M.; Untersmayr, E. The role of gastrointestinal permeability in food allergy. Ann. Allergy Asthma Immunol. 2018, 121, 168–173. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Score | Symptoms |
|---|---|
| 0 | No symptoms |
| 1 | Scratching nose and mouth |
| 2 | Swelling around the eyes and mouth; pillar erection; reduced activity; higher breathing rate |
| 3 | Shortness of breath; blue rash around the mouth and tail; higher breathing rate |
| 4 | No activity after stimulation; shivering and muscle contractions |
| 5 | Death by shock |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bai, J.; Zhou, Y.; Xia, X.; Wu, Z.; Li, X.; Tong, P.; Yang, A.; Chen, H. Transglutaminase-Cross-Linked Tofu Suppressed Soybean-Induced Allergic Reactions by Enhancing Intestinal Mucosa Immune Tolerance. Foods 2024, 13, 1206. https://doi.org/10.3390/foods13081206
Bai J, Zhou Y, Xia X, Wu Z, Li X, Tong P, Yang A, Chen H. Transglutaminase-Cross-Linked Tofu Suppressed Soybean-Induced Allergic Reactions by Enhancing Intestinal Mucosa Immune Tolerance. Foods. 2024; 13(8):1206. https://doi.org/10.3390/foods13081206
Chicago/Turabian StyleBai, Jing, Yiling Zhou, Xinlei Xia, Zhihua Wu, Xin Li, Ping Tong, Anshu Yang, and Hongbing Chen. 2024. "Transglutaminase-Cross-Linked Tofu Suppressed Soybean-Induced Allergic Reactions by Enhancing Intestinal Mucosa Immune Tolerance" Foods 13, no. 8: 1206. https://doi.org/10.3390/foods13081206