
Kahweol Inhibits Pro-Inflammatory Cytokines and Chemokines in Tumor Necrosis Factor-α/Interferon-γ-Stimulated Human Keratinocyte HaCaT Cells


Abstract
:1. Introduction
2. Materials and Methods
2.1. Cell Culture
2.2. Cell Viability Assay
2.3. Quantitative Real-Time Polymerase Chain Reaction (qPCR)
2.4. Enzyme-Linked Immunosorbent Assay (ELISA)
2.5. Western Blot Analysis
2.6. Electrophoretic Mobility Shift Assay (EMSA)
2.7. Immunofluorescence (IF) Staining
2.8. Statistical Analysis
3. Results
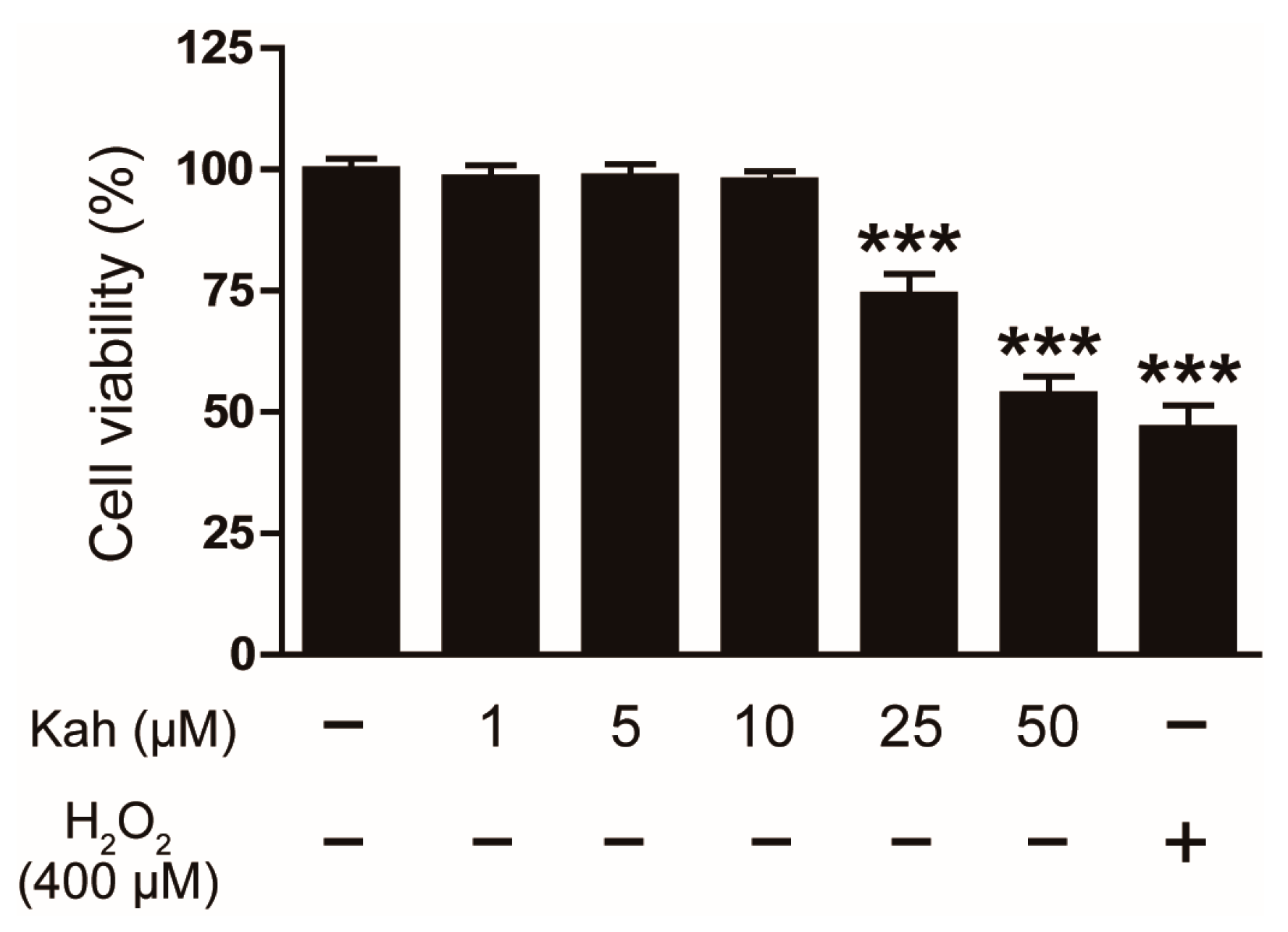
3.1. Impact of Kahweol on the Viability of Human Keratinocyte HaCaT Cells
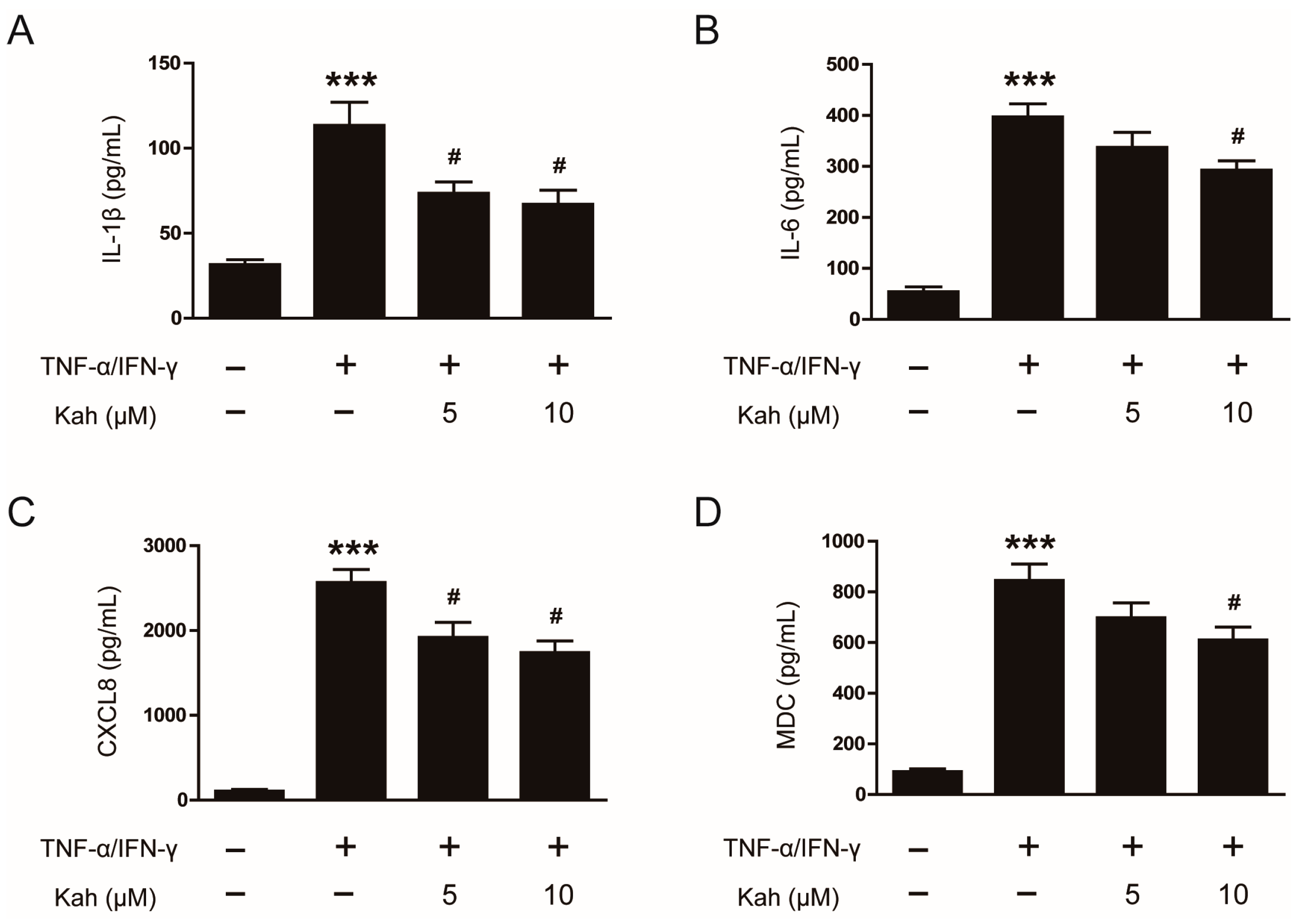
3.2. Impact of Kahweol on Cytokine and Chemokine Production in TNF-α/IFN-γ-Activated Keratinocytes
3.3. Impact of Kahweol on Mitogen-Activated Protein Kinase (MAPK) Signaling Cascades in TNF-α/IFN-γ-Activated Keratinocytes
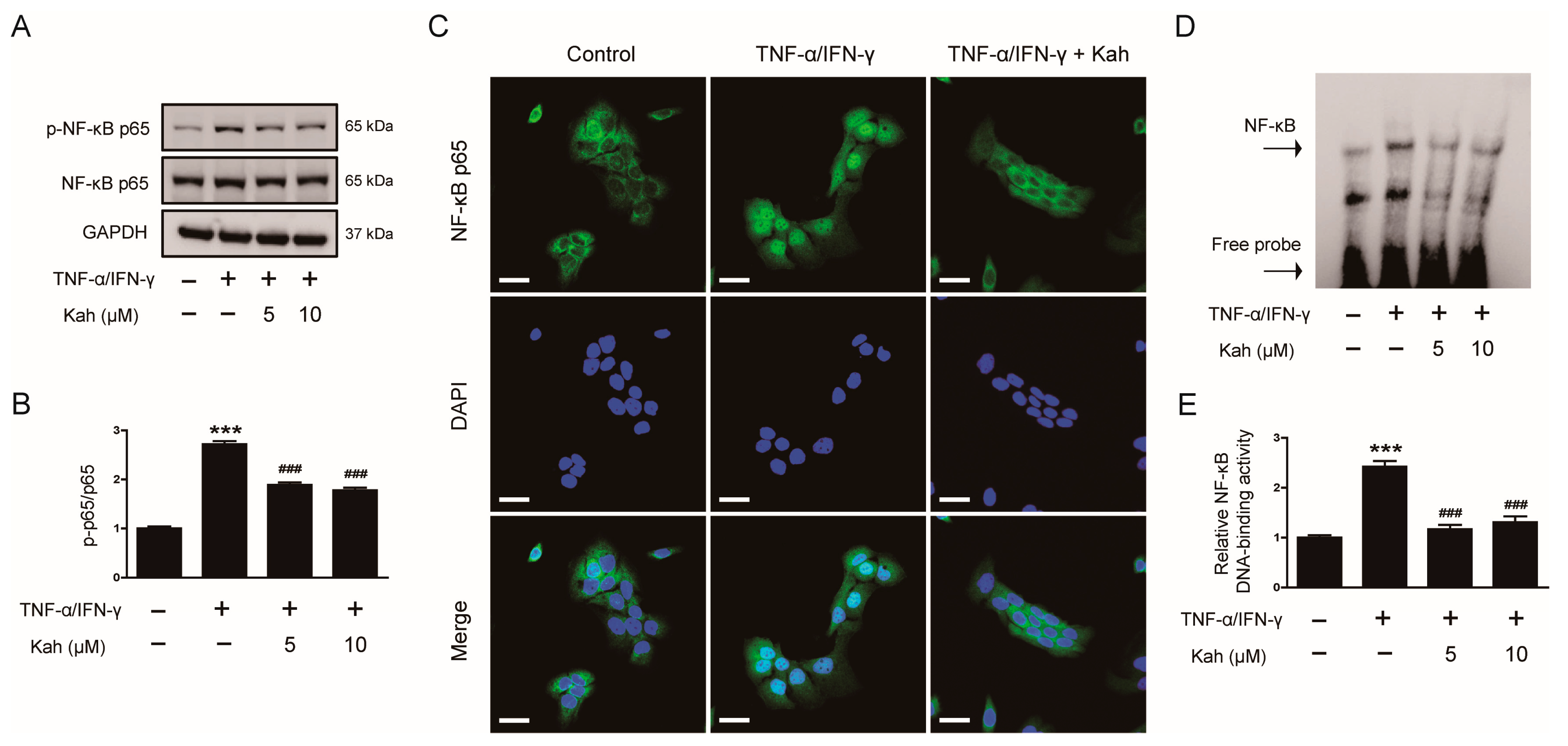
3.4. Impact of Kahweol on NF-κB Activation in TNF-α/IFN-γ-Activated Keratinocytes
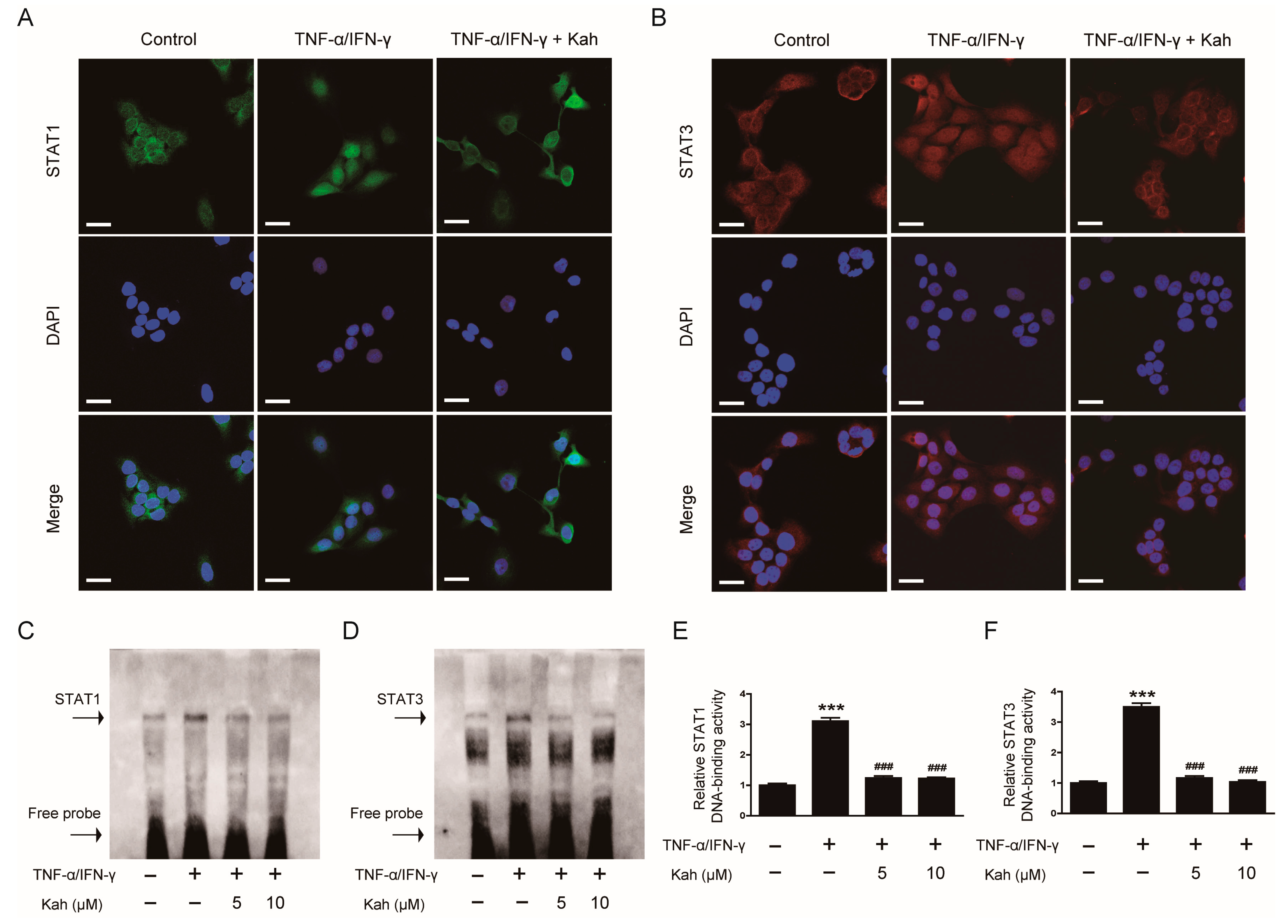
3.5. Impact of Kahweol on STAT1 and STAT3 Activation in TNF-α/IFN-γ-Activated Keratinocytes
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Schuler, C.F., 4th; Billi, A.C.; Maverakis, E.; Tsoi, L.C.; Gudjonsson, J.E. Novel insights into atopic dermatitis. J. Allergy Clin. Immunol. 2023, 151, 1145–1154. [Google Scholar] [CrossRef]
- Weidinger, S.; Novak, N. Atopic dermatitis. Lancet 2016, 387, 1109–1122. [Google Scholar] [CrossRef]
- Eichenfield, L.F.; Ahluwalia, J.; Waldman, A.; Borok, J.; Udkoff, J.; Boguniewicz, M. Current guidelines for the evaluation and management of atopic dermatitis: A comparison of the Joint Task Force Practice Parameter and American Academy of Dermatology guidelines. J. Allergy Clin. Immunol. 2017, 139, S49–S57. [Google Scholar] [CrossRef]
- Bieber, T. Atopic dermatitis: An expanding therapeutic pipeline for a complex disease. Nat. Rev. Drug Discov. 2022, 21, 21–40. [Google Scholar] [CrossRef]
- Butala, S.; Castelo-Soccio, L.; Seshadri, R.; Simpson, E.L.; O’Shea, J.J.; Bieber, T.; Paller, A.S. Biologic Versus Small Molecule Therapy for Treating Moderate to Severe Atopic Dermatitis: Clinical Considerations. J. Allergy Clin. Immunol. Pract. 2023, 11, 1361–1373. [Google Scholar] [CrossRef]
- Malik, K.; Heitmiller, K.D.; Czarnowicki, T. An Update on the Pathophysiology of Atopic Dermatitis. Dermatol. Clin. 2017, 35, 317–326. [Google Scholar] [CrossRef]
- Yang, G.; Seok, J.K.; Kang, H.C.; Cho, Y.-Y.; Lee, H.S.; Lee, J.Y. Skin Barrier Abnormalities and Immune Dysfunction in Atopic Dermatitis. Int. J. Mol. Sci. 2020, 21, 2867. [Google Scholar] [CrossRef]
- Elias, P.M.; Steinhoff, M. “Outside-to-inside” (and now back to “outside”) pathogenic mechanisms in atopic dermatitis. J. Investig. Dermatol. 2008, 128, 1067–1070. [Google Scholar] [CrossRef]
- Lefèvre-Utile, A.; Braun, C.; Haftek, M.; Aubin, F. Five Functional Aspects of the Epidermal Barrier. Int. J. Mol. Sci. 2021, 22, 11676. [Google Scholar] [CrossRef]
- Das, P.; Mounika, P.; Yellurkar, M.L.; Prasanna, V.S.; Sarkar, S.; Velayutham, R.; Arumugam, S. Keratinocytes: An Enigmatic Factor in Atopic Dermatitis. Cells 2022, 11, 1683. [Google Scholar] [CrossRef]
- Chieosilapatham, P.; Kiatsurayanon, C.; Umehara, Y.; Trujillo-Paez, J.V.; Peng, G.; Yue, H.; Nguyen, L.T.H.; Niyonsaba, F. Keratinocytes: Innate immune cells in atopic dermatitis. Clin. Exp. Immunol. 2021, 204, 296–309. [Google Scholar] [CrossRef]
- Gwon, M.G.; Leem, J.; An, H.J.; Gu, H.; Bae, S.; Kim, J.H.; Park, K.K. The decoy oligodeoxynucleotide against HIF-1α and STAT5 ameliorates atopic dermatitis-like mouse model. Mol. Ther. Nucleic Acids 2023, 34, 102036. [Google Scholar] [CrossRef]
- Najmi, A.; Javed, S.A.; Al Bratty, M.; Alhazmi, H.A. Modern Approaches in the Discovery and Development of Plant-Based Natural Products and Their Analogues as Potential Therapeutic Agents. Molecules 2022, 27, 349. [Google Scholar] [CrossRef]
- Čižmárová, B.; Hubková, B.; Tomečková, V.; Birková, A. Flavonoids as Promising Natural Compounds in the Prevention and Treatment of Selected Skin Diseases. Int. J. Mol. Sci. 2023, 24, 6324. [Google Scholar] [CrossRef]
- Karuppagounder, V.; Arumugam, S.; Thandavarayan, R.A.; Pitchaimani, V.; Sreedhar, R.; Afrin, R.; Harima, M.; Suzuki, H.; Nomoto, M.; Miyashita, S.; et al. Modulation of HMGB1 translocation and RAGE/NFκB cascade by quercetin treatment mitigates atopic dermatitis in NC/Nga transgenic mice. Exp. Dermatol. 2015, 24, 418–423. [Google Scholar] [CrossRef]
- Carlucci, C.D.; Hui, Y.; Chumanevich, A.P.; Robida, P.A.; Fuseler, J.W.; Sajish, M.; Nagarkatti, P.; Nagarkatti, M.; Oskeritzian, C.A. Resveratrol Protects against Skin Inflammation through Inhibition of Mast Cell, Sphingosine Kinase-1, Stat3 and NF-κB p65 Signaling Activation in Mice. Int. J. Mol. Sci. 2023, 24, 6707. [Google Scholar] [CrossRef]
- Sharma, S.; Sethi, G.S.; Naura, A.S. Curcumin Ameliorates Ovalbumin-Induced Atopic Dermatitis and Blocks the Progression of Atopic March in Mice. Inflammation 2020, 43, 358–369. [Google Scholar] [CrossRef]
- Socała, K.; Szopa, A.; Serefko, A.; Poleszak, E.; Wlaź, P. Neuroprotective Effects of Coffee Bioactive Compounds: A Review. Int. J. Mol. Sci. 2021, 22, 107. [Google Scholar] [CrossRef]
- Bravi, F.; Tavani, A.; Bosetti, C.; Boffetta, P.; La Vecchia, C. Coffee and the risk of hepatocellular carcinoma and chronic liver disease: A systematic review and meta-analysis of prospective studies. Eur. J. Cancer Prev. 2017, 26, 368–377. [Google Scholar] [CrossRef]
- Barrea, L.; Pugliese, G.; Frias-Toral, E.; El Ghoch, M.; Castellucci, B.; Chapela, S.P.; Carignano, M.L.A.; Laudisio, D.; Savastano, S.; Colao, A.; et al. Coffee consumption, health benefits and side effects: A narrative review and update for dietitians and nutritionists. Crit. Rev. Food Sci. Nutr. 2023, 63, 1238–1261. [Google Scholar] [CrossRef]
- Chieng, D.; Kistler, P.M. Coffee and tea on cardiovascular disease (CVD) prevention. Trends Cardiovasc. Med. 2022, 32, 399–405. [Google Scholar] [CrossRef]
- Park, S.; Choi, H.S.; Bae, J.H. Instant noodles, processed food intake, and dietary pattern are associated with atopic dermatitis in an adult population (KNHANES 2009–2011). Asia Pac. J. Clin. Nutr. 2016, 25, 602–613. [Google Scholar]
- Barrea, L.; Muscogiuri, G.; Di Somma, C.; Annunziata, G.; Megna, M.; Falco, A.; Balato, A.; Colao, A.; Savastano, S. Coffee consumption, metabolic syndrome and clinical severity of psoriasis: Good or bad stuff? Arch. Toxicol. 2018, 92, 1831–1845. [Google Scholar] [CrossRef]
- Ren, Y.; Wang, C.; Xu, J.; Wang, S. Cafestol and Kahweol: A Review on Their Bioactivities and Pharmacological Properties. Int. J. Mol. Sci. 2019, 20, 4238. [Google Scholar] [CrossRef]
- Eldesouki, S.; Qadri, R.; Abu Helwa, R.; Barqawi, H.; Bustanji, Y.; Abu-Gharbieh, E.; El-Huneidi, W. Recent Updates on the Functional Impact of Kahweol and Cafestol on Cancer. Molecules 2022, 27, 7332. [Google Scholar] [CrossRef]
- Kim, J.Y.; Kim, D.H.; Jeong, H.G. Inhibitory effect of the coffee diterpene kahweol on carrageenan-induced inflammation in rats. Biofactors 2006, 26, 17–28. [Google Scholar] [CrossRef]
- Lee, K.J.; Choi, J.H.; Jeong, H.G. Hepatoprotective and antioxidant effects of the coffee diterpenes kahweol and cafestol on carbon tetrachloride-induced liver damage in mice. Food Chem. Toxicol. 2007, 45, 2118–2125. [Google Scholar]
- Seo, H.-Y.; Jung, Y.-A.; Lee, S.-H.; Hwang, J.S.; Park, K.-G.; Kim, M.-K.; Jang, B.K. Kahweol decreases hepatic fibrosis by inhibiting the expression of connective tissue growth factor via the transforming growth factor-beta signaling pathway. Oncotarget 2017, 8, 87086–87094. [Google Scholar] [CrossRef]
- Lee, H.-F.; Lin, J.S.; Chang, C.-F. Acute Kahweol Treatment Attenuates Traumatic Brain Injury Neuroinflammation and Functional Deficits. Nutrients 2019, 11, 2301. [Google Scholar] [CrossRef]
- Kim, J.-Y.; Jo, J.; Leem, J.; Park, K.-K. Kahweol Ameliorates Cisplatin-Induced Acute Kidney Injury through Pleiotropic Effects in Mice. Biomedicines 2020, 8, 572. [Google Scholar] [CrossRef]
- Kim, J.-Y.; Leem, J.; Kim, G.M. Kahweol Protects against Acetaminophen-Induced Hepatotoxicity in Mice through Inhibiting Oxidative Stress, Hepatocyte Death, and Inflammation. BioMed Res. Int. 2022, 2022, 8121124. [Google Scholar] [CrossRef]
- An, H.-J.; Gwon, M.-G.; Gu, H.; Bae, S.; Leem, J.; Lee, J.B.; Park, K.-K. STAT3/NF-κB decoy oligodeoxynucleotides inhibit atherosclerosis through regulation of the STAT/NF-κB signaling pathway in a mouse model of atherosclerosis. Int. J. Mol. Med. 2023, 51, 37. [Google Scholar] [CrossRef]
- Kjellerup, R.B.; Kragballe, K.; Iversen, L.; Johansen, C. Pro-inflammatory cytokine release in keratinocytes is mediated through the MAPK signal-integrating kinases. Exp. Dermatol. 2008, 17, 498–504. [Google Scholar] [CrossRef]
- Kim, S.Y.; Sohn, E.J.; Kim, D.W.; Jeong, H.J.; Kim, M.J.; Kang, H.W.; Shin, M.J.; Ahn, E.H.; Kwon, S.W.; Kim, Y.N.; et al. Transduced PEP-1-FK506BP ameliorates atopic dermatitis in NC/Nga mice. J. Investig. Dermatol. 2011, 131, 1477–1485. [Google Scholar] [CrossRef]
- Kim, T.-Y.; Park, N.-J.; Jo, B.-G.; Lee, B.S.; Keem, M.-J.; Kwon, T.-H.; Kim, K.H.; Kim, S.-N.; Yang, M.H. Anti-Wrinkling Effect of 3,4,5-tri-O-caffeoylquinic Acid from the Roots of Nymphoides peltata through MAPK/AP-1, NF-κB, and Nrf2 Signaling in UVB-Irradiated HaCaT Cells. Antioxidants 2023, 12, 1899. [Google Scholar] [CrossRef]
- Huang, J.C.; Yue, Z.P.; Yu, H.F.; Yang, Z.Q.; Wang, Y.S.; Guo, B. TAZ ameliorates the microglia-mediated inflammatory response via the Nrf2-ROS-NF-κB pathway. Mol. Ther. Nucleic Acids 2022, 28, 435–449. [Google Scholar] [CrossRef]
- Tsiogka, A.; Kyriazopoulou, M.; Kontochristopoulos, G.; Nicolaidou, E.; Stratigos, A.; Rigopoulos, D.; Gregoriou, S. The JAK/STAT Pathway and Its Selective Inhibition in the Treatment of Atopic Dermatitis: A Systematic Review. J. Clin. Med. 2022, 11, 4431. [Google Scholar] [CrossRef]
- Huang, I.-H.; Chung, W.H.; Wu, P.C.; Chen, C.B. JAK-STAT signaling pathway in the pathogenesis of atopic dermatitis: An updated review. Front. Immunol. 2022, 13, 1068260. [Google Scholar] [CrossRef]
- Tsai, Y.-C.; Chang, H.-H.; Chou, S.-C.; Chu, T.W.; Hsu, Y.-J.; Hsiao, C.-Y.; Lo, Y.-H.; Wu, N.-L.; Chang, D.-C.; Hung, C.-F. Evaluation of the Anti-Atopic Dermatitis Effects of α-Boswellic Acid on Tnf-α/Ifn-γ-Stimulated HaCat Cells and DNCB-Induced BALB/c Mice. Int. J. Mol. Sci. 2022, 23, 9863. [Google Scholar] [CrossRef]
- Dong, L.; Lee, H.; Liu, Z.; Lee, D.-S. Anti-Skin Inflammatory and Anti-Oxidative Effects of the Neoflavonoid Latifolin Isolated from Dalbergia odorifera in HaCaT and BJ-5ta Cells. Int. J. Mol. Sci. 2023, 24, 7371. [Google Scholar] [CrossRef]
- Oh, J.-H.; Kim, S.-H.; Kwon, O.-K.; Kim, J.-H.; Oh, S.-R.; Han, S.-B.; Park, J.-W.; Ahn, K.-S. Purpurin suppresses atopic dermatitis via TNF-α/IFN-γ-induced inflammation in HaCaT cells. Int. J. Immunopathol. Pharmacol. 2022, 36, 3946320221111135. [Google Scholar] [CrossRef]
- Humeau, M.; Boniface, K.; Bodet, C. Cytokine-Mediated Crosstalk between Keratinocytes and T Cells in Atopic Dermatitis. Front. Immunol. 2022, 13, 801579. [Google Scholar] [CrossRef]
- Arthur, J.S.; Ley, S.C. Mitogen-activated protein kinases in innate immunity. Nat. Rev. Immunol. 2013, 13, 679–692. [Google Scholar] [CrossRef]
- Álvarez, S.; Contreras-Kallens, P.; Aguayo, S.; Ramírez, O.; Vallejos, C.; Ruiz, J.; Carrasco-Gallardo, E.; Troncoso-Vera, S.; Morales, B.; Schuh, C.M.A.P. Royal jelly extracellular vesicles promote wound healing by modulating underlying cellular responses. Mol. Ther. Nucleic Acids 2023, 31, 541–552. [Google Scholar] [CrossRef]
- Kim, M.-J.; Hwang, B.S.; Hwang, Y.; Jeong, Y.T.; Jeong, D.W.; Oh, Y.T. Anti-Inflammatory and Antiatopic Effects of Rorippa cantoniensis (Lour.) Ohwi in RAW 264.7 and HaCaT Cells. Molecules 2023, 28, 5463. [Google Scholar] [CrossRef]
- Hong, S.; Kim, E.-Y.; Lim, S.-E.; Kim, J.-H.; Sohn, Y.; Jung, H.-S. Dendrobium nobile Lindley Administration Attenuates Atopic Dermatitis-like Lesions by Modulating Immune Cells. Int. J. Mol. Sci. 2022, 23, 4470. [Google Scholar] [CrossRef]
- Kong, L.; Liu, J.; Wang, J.; Luo, Q.; Zhang, H.; Liu, B.; Xu, F.; Pang, Q.; Liu, Y.; Dong, J. Icariin inhibits TNF-α/IFN-γ induced inflammatory response via inhibition of the substance P and p38-MAPK signaling pathway in human keratinocytes. Int. Immunopharmacol. 2015, 29, 401–407. [Google Scholar] [CrossRef]
- Hammouda, M.B.; Ford, A.E.; Liu, Y.; Zhang, J.Y. The JNK Signaling Pathway in Inflammatory Skin Disorders and Cancer. Cells 2020, 9, 857. [Google Scholar] [CrossRef]
- Shibata, S.; Tada, Y.; Asano, Y.; Hau, C.S.; Kato, T.; Saeki, H.; Yamauchi, T.; Kubota, N.; Kadowaki, T.; Sato, S. Adiponectin regulates cutaneous wound healing by promoting keratinocyte proliferation and migration via the ERK signaling pathway. J. Immunol. 2012, 189, 3231–3241. [Google Scholar]
- Wang, H.; Lei, L.; Hu, J.; Li, Y. Oncostatin M upregulates Livin to promote keratinocyte proliferation and survival via ERK and STAT3 signalling pathways. Exp. Physiol. 2020, 105, 1151–1158. [Google Scholar] [CrossRef]
- Zeze, N.; Kido-Nakahara, M.; Tsuji, G.; Maehara, E.; Sato, Y.; Sakai, S.; Fujishima, K.; Hashimoto-Hachiya, A.; Furue, M.; Nakahara, T. Role of ERK Pathway in the Pathogenesis of Atopic Dermatitis and Its Potential as a Therapeutic Target. Int. J. Mol. Sci. 2022, 23, 3467. [Google Scholar] [CrossRef]
- Meng, X.; Qiu, L.; Song, H.; Dang, N. MAPK Pathway Involved in Epidermal Terminal Differentiation of Normal Human Epidermal Keratinocytes. Open Med. 2018, 13, 189–195. [Google Scholar] [CrossRef]
- Ren, X.; Shi, Y.; Zhao, D.; Xu, M.; Li, X.; Dang, Y.; Ye, X. Naringin protects ultraviolet B-induced skin damage by regulating p38 MAPK signal pathway. J. Dermatol. Sci. 2016, 82, 106–114. [Google Scholar] [CrossRef]
- Liu, T.; Zhang, L.; Joo, D.; Sun, S.C. NF-κB signaling in inflammation. Signal Transduct. Target. Ther. 2017, 2, 17023. [Google Scholar] [CrossRef]
- Shen, T.; Park, Y.C.; Kim, S.H.; Lee, J.; Cho, J.Y. Nuclear factor-kappaB/signal transducers and activators of transcription-1-mediated inflammatory responses in lipopolysaccharide-activated macrophages are a major inhibitory target of kahweol, a coffee diterpene. Biol. Pharm. Bull. 2010, 33, 1159–1164. [Google Scholar] [CrossRef]
- Kim, J.Y.; Jung, K.S.; Lee, K.J.; Na, H.K.; Chun, H.-K.; Kho, Y.-H.; Jeong, H.G. The coffee diterpene kahweol suppress the inducible nitric oxide synthase expression in macrophages. Cancer Lett. 2004, 213, 147–154. [Google Scholar] [CrossRef]
- Kim, H.G.; Kim, J.Y.; Hwang, Y.P.; Lee, K.J.; Lee, K.Y.; Kim, D.H.; Kim, D.H.; Jeong, H.G. The coffee diterpene kahweol inhibits tumor necrosis factor-alpha-induced expression of cell adhesion molecules in human endothelial cells. Toxicol. Appl. Pharmacol. 2006, 217, 332–341. [Google Scholar] [CrossRef]
- Seo, H.-Y.; Kim, M.-K.; Lee, S.-H.; Hwang, J.S.; Park, K.-G.; Jang, B.K. Kahweol Ameliorates the Liver Inflammation through the Inhibition of NF-κB and STAT3 Activation in Primary Kupffer Cells and Primary Hepatocytes. Nutrients 2018, 10, 863. [Google Scholar] [CrossRef]
- El-Huneidi, W.; Anjum, S.; Bajbouj, K.; Abu-Gharbieh, E.; Taneera, J. The Coffee Diterpene, Kahweol, Ameliorates Pancreatic β-Cell Function in Streptozotocin (STZ)-Treated Rat INS-1 Cells through NF-kB and p-AKT/Bcl-2 Pathways. Molecules 2021, 26, 5167. [Google Scholar] [CrossRef]
- Amano, W.; Nakajima, S.; Kunugi, H.; Numata, Y.; Kitoh, A.; Egawa, G.; Dainichi, T.; Honda, T.; Otsuka, A.; Kimoto, Y.; et al. The Janus kinase inhibitor JTE-052 improves skin barrier function through suppressing signal transducer and activator of transcription 3 signaling. J. Allergy Clin. Immunol. 2015, 136, 667–677.e7. [Google Scholar] [CrossRef]
- Kim, J.; Kim, M.G.; Jeong, S.H.; Kim, H.J.; Son, S.W. STAT3 maintains skin barrier integrity by modulating SPINK5 and KLK5 expression in keratinocytes. Exp. Dermatol. 2022, 31, 223–232. [Google Scholar] [CrossRef]
- Sano, S.; Chan, K.S.; DiGiovanni, J. Impact of Stat3 activation upon skin biology: A dichotomy of its role between homeostasis and diseases. J. Dermatol. Sci. 2008, 50, 1–14. [Google Scholar] [CrossRef]
- Gil, T.-Y.; Hong, C.-H.; An, H.-J. Anti-Inflammatory Effects of Ellagic Acid on Keratinocytes via MAPK and STAT Pathways. Int. J. Mol. Sci. 2021, 22, 1277. [Google Scholar] [CrossRef]
- Kim, W.H.; An, H.J.; Kim, J.Y.; Gwon, M.G.; Gu, H.; Lee, S.J.; Park, J.Y.; Park, K.D.; Han, S.M.; Kim, M.K.; et al. Apamin inhibits TNF-α- and IFN-γ-induced inflammatory cytokines and chemokines via suppressions of NF-κB signaling pathway and STAT in human keratinocytes. Pharmacol. Rep. 2017, 69, 1030–1035. [Google Scholar] [CrossRef]
- Ito, M.; Morita, T.; Okazaki, S.; Koto, M.; Ichikawa, Y.; Takayama, R.; Hoashi, T.; Saeki, H.; Kanda, N. Dietary habits in adult Japanese patients with atopic dermatitis. J. Dermatol. 2019, 46, 515–521. [Google Scholar] [CrossRef]
- Chiricozzi, A.; Maurelli, M.; Calabrese, L.; Peris, K.; Girolomoni, G. Overview of Atopic Dermatitis in Different Ethnic Groups. J. Clin. Med. 2023, 12, 2701. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Primer Sequence (5′→3′) | Accession No. |
|---|---|---|
| IL-1β | F: ATGCACCTGTACGATCACTG R: ACAAAGGACATGGAGAACACC | NM_000576 |
| IL-6 | F: CCACTCACCTCTTCAGAACG R: CATCTTTGGAAGGTTCAGGTTG | NM_000600 |
| CXCL8 | F: CTGCGCCAACACAGAAATTA R: ATTGCATCTGGCAACCCTAC | NM_000584 |
| MDC | F: AGGACAGAGCATGGCTCGCCTACAGA R: TAATGGCAGGGAGGTAGGGCTCCTGA | NM_002990 |
| GAPDH | F: GTCTCCTCTGACTTCAACAGCG R: ACCACCCTGTTGCTGTAGCCAA | NM_002046 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kwon, Y.J.; Kwon, H.H.; Leem, J.; Jang, Y.Y. Kahweol Inhibits Pro-Inflammatory Cytokines and Chemokines in Tumor Necrosis Factor-α/Interferon-γ-Stimulated Human Keratinocyte HaCaT Cells. Curr. Issues Mol. Biol. 2024, 46, 3470-3483. https://doi.org/10.3390/cimb46040218
Kwon YJ, Kwon HH, Leem J, Jang YY. Kahweol Inhibits Pro-Inflammatory Cytokines and Chemokines in Tumor Necrosis Factor-α/Interferon-γ-Stimulated Human Keratinocyte HaCaT Cells. Current Issues in Molecular Biology. 2024; 46(4):3470-3483. https://doi.org/10.3390/cimb46040218
Chicago/Turabian StyleKwon, Ye Jin, Hyun Hee Kwon, Jaechan Leem, and Yoon Young Jang. 2024. "Kahweol Inhibits Pro-Inflammatory Cytokines and Chemokines in Tumor Necrosis Factor-α/Interferon-γ-Stimulated Human Keratinocyte HaCaT Cells" Current Issues in Molecular Biology 46, no. 4: 3470-3483. https://doi.org/10.3390/cimb46040218