
Rhoa/ROCK, mTOR and Secretome-Based Treatments for Ischemic Stroke: New Perspectives

 , and
, and
Abstract
:1. Introduction
2. Search Strategy
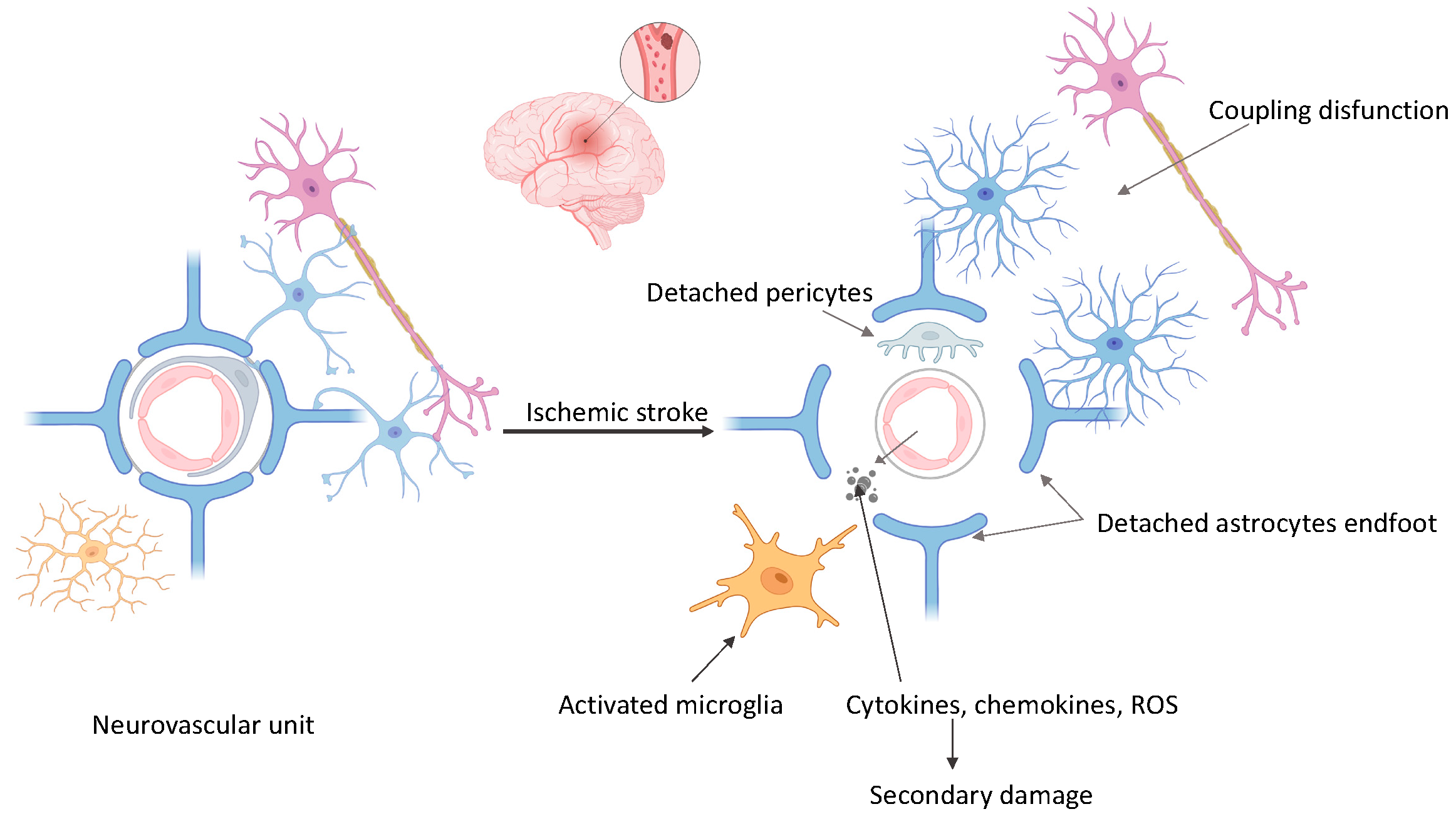
3. The Neurovascular Unit and Ischemic Cascade
4. Targeting Cellular Pathways in Ischemic Stroke Treatment
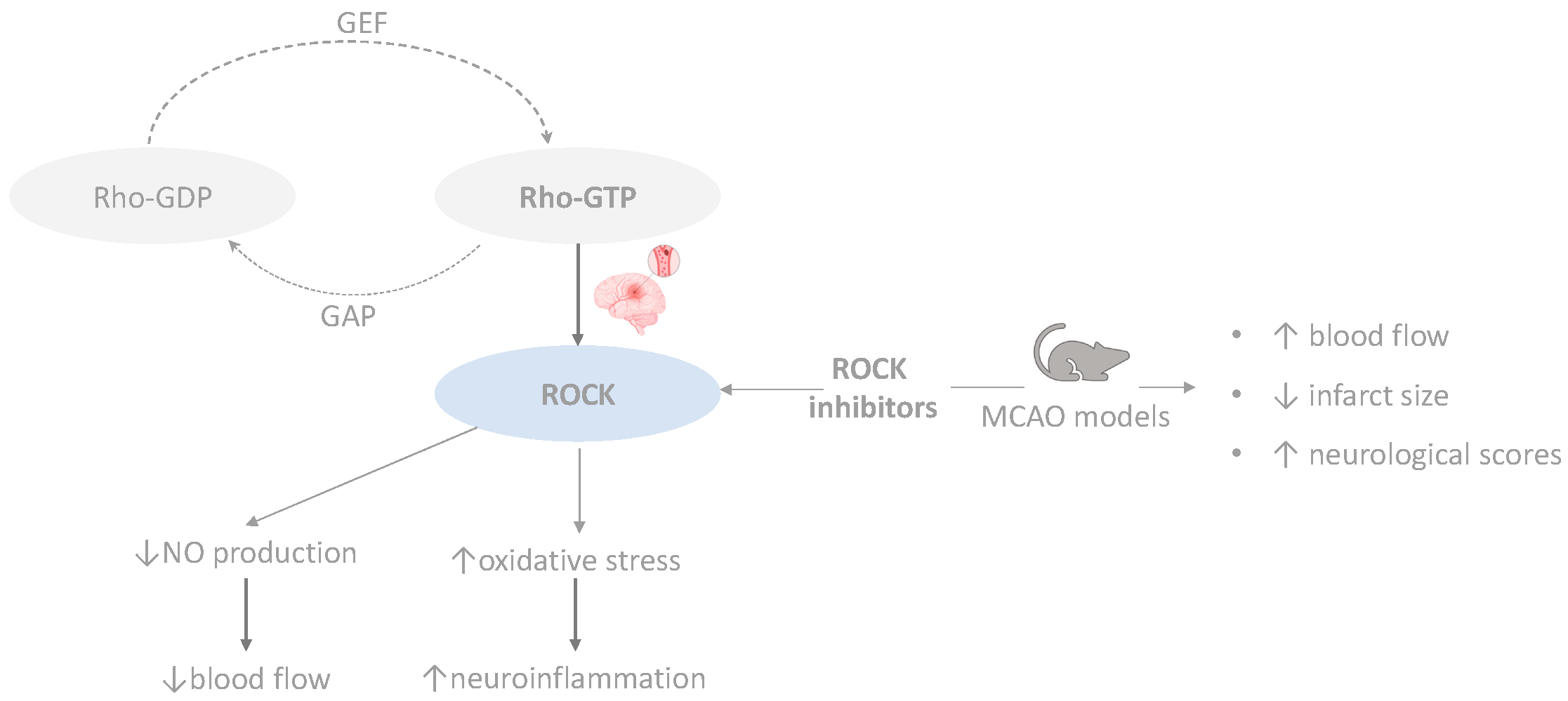
4.1. RhoA/ROCK Pathway
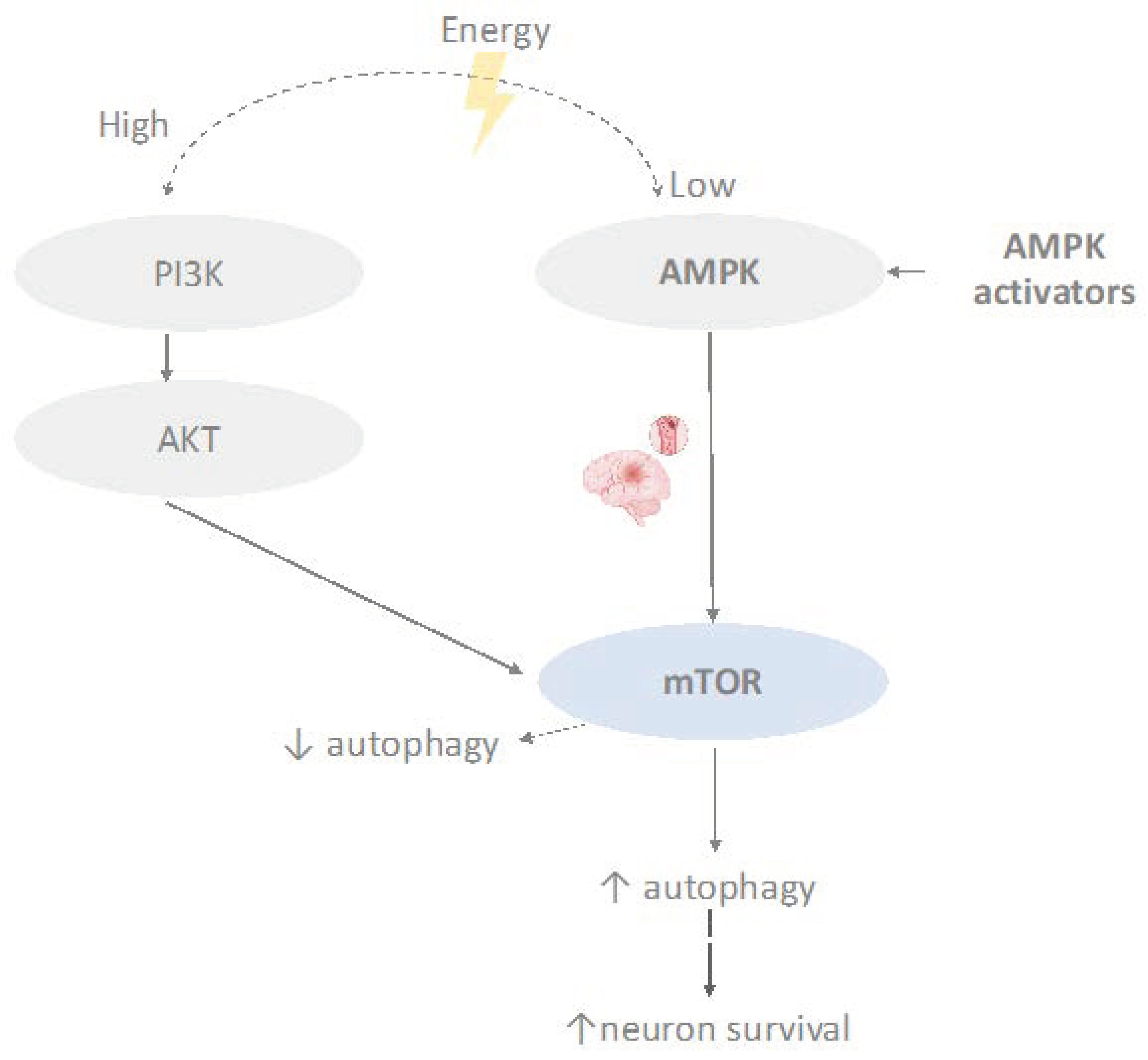
4.2. mTOR Signaling Pathway
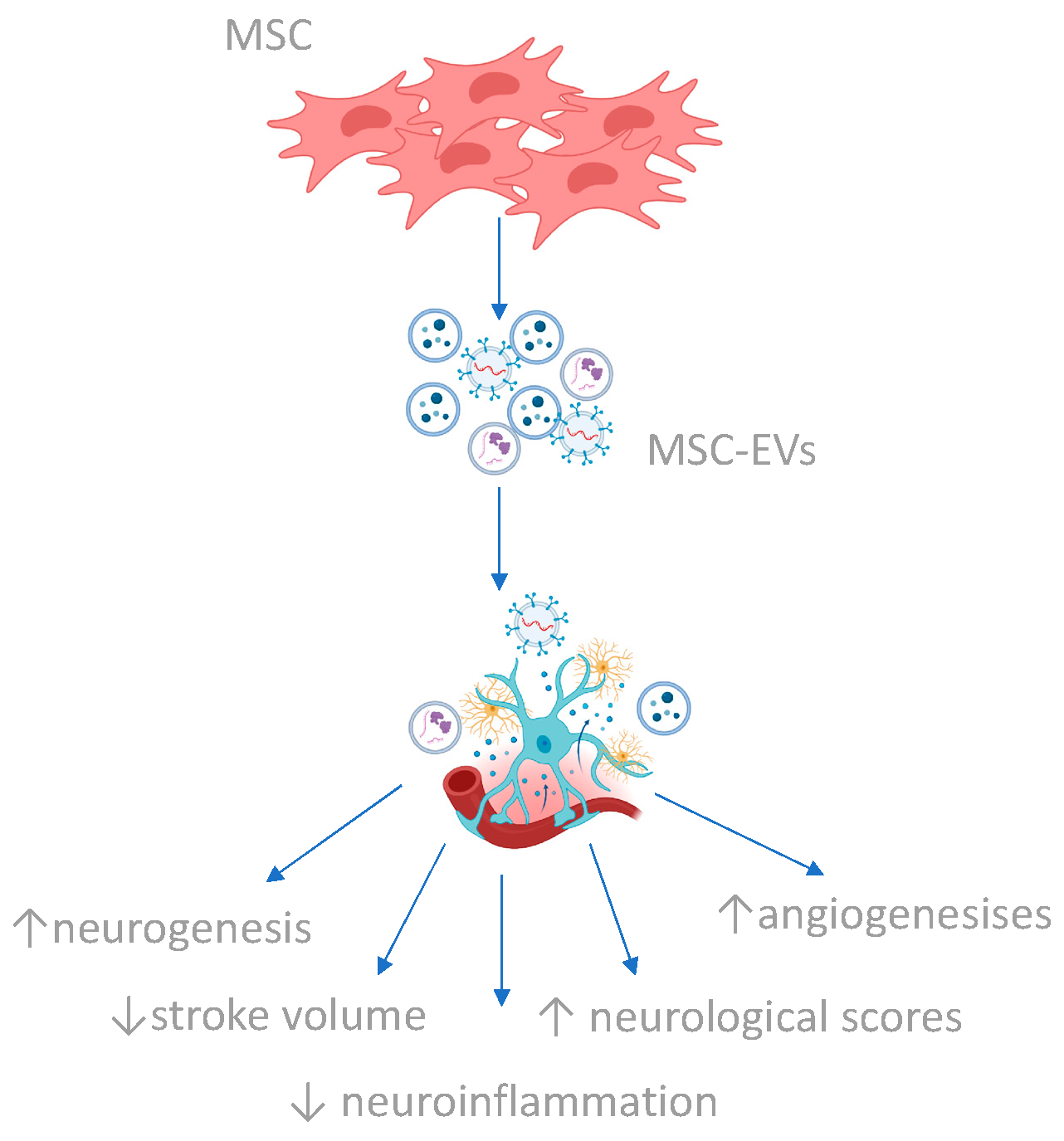
4.3. Secretome
5. Discussion
6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- GBD 2019 Stroke Collaborators. Global, regional, and national burden of stroke and its risk factors, 1990–2019: A systematic analysis for the Global Burden of Disease Study 2019. Lancet Neurol. 2021, 20, 795–820. [Google Scholar] [CrossRef]
- Feigin, V.L.; Brainin, M.; Norrving, B.; Martins, S.; Sacco, R.L.; Hacke, W.; Fisher, M.; Pandian, J.; Lindsay, P. World Stroke Organization (WSO): Global Stroke Fact Sheet 2022. Int. J. Stroke 2022, 17, 18–29, Erratum in Int. J. Stroke 2022, 17, 478. [Google Scholar] [CrossRef]
- Tsivgoulis, G.; Zand, R.; Katsanos, A.H.; Goyal, N.; Uchino, K.; Chang, J. Safety of intravenous thrombolysis in stroke mimics: Prospective 5-year study and comprehensive meta-analysis. Stroke 2015, 46, 1281–1287. [Google Scholar] [CrossRef] [PubMed]
- Ramadan, A.R.; Denny, M.C.; Vahidy, F.; Yamal, J.M.; Wu, T.C.; Sarraj, A. Agreement among Stroke Faculty and Fellows in Treating Ischemic Stroke Patients with Tissue-Type Plasminogen Activator and Thrombectomy. Stroke 2017, 48, 222–224. [Google Scholar] [CrossRef]
- Powers, W.J.; Rabinstein, A.A.; Ackerson, T.; Adeoye, O.M.; Bambakidis, N.C.; Becker, K.; Biller, J.; Brown, M.; Demaerschalk, B.M.; Hoh, B.; et al. 2018 Guidelines for the Early Management of Patients with Acute Ischemic Stroke: A Guideline for Healthcare Professionals from the American Heart Association/American Stroke Association. Stroke 2018, 49, e46–e99. [Google Scholar] [CrossRef] [PubMed]
- Moskowitz, M.A.; Lo, E.H.; Iadecola, C. The science of stroke: Mechanisms in search of treatments. Neuron 2010, 67, 181–198, Erratum in Neuron 2010, 68, 161. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.; Chen, S.; Luo, Y.; Han, Z. Crosstalk between Inflammation and the BBB in Stroke. Curr. Neuropharmacol. 2020, 18, 1227–1236. [Google Scholar] [CrossRef] [PubMed]
- Candelario-Jalil, E.; Dijkhuizen, R.M.; Magnus, T. Neuroinflammation, Stroke, Blood-Brain Barrier Dysfunction, and Imaging Modalities. Stroke 2022, 53, 1473–1486. [Google Scholar] [CrossRef]
- Ermine, C.M.; Bivard, A.; Parsons, M.W.; Baron, J.C. The ischemic penumbra: From concept to reality. Int. J. Stroke 2021, 16, 497–509. [Google Scholar] [CrossRef]
- Wang, L.; Xiong, X.; Zhang, L.; Shen, J. Neurovascular Unit: A critical role in ischemic stroke. CNS Neurosci. Ther. 2021, 27, 7–16. [Google Scholar] [CrossRef]
- Przykaza, Ł. Understanding the Connection between Common Stroke Comorbidities, Their Associated Inflammation, and the Course of the Cerebral Ischemia/Reperfusion Cascade. Front. Immunol. 2021, 12, 782569. [Google Scholar] [CrossRef] [PubMed]
- Wardlaw, J.M.; Murray, V.; Berge, E.; del Zoppo, G.J. Thrombolysis for acute ischaemic stroke. Cochrane Database Syst. Rev. 2014, 2014, CD000213. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Li, Q.; Wang, J.; Zhuang, Q.K.; Zhang, Y.Y. Combination of thrombolytic therapy and neuroprotective therapy in acute ischemic stroke: Is it important? Eur. Rev. Med. Pharmacol. Sci. 2015, 19, 416–422. [Google Scholar] [PubMed]
- Vesterinen, H.M.; Currie, G.L.; Carter, S.; Mee, S.; Watzlawick, R.; Egan, K.J.; Macleod, M.R.; Sena, E.S. Systematic review and stratified meta-analysis of the efficacy of RhoA and Rho kinase inhibitors in animal models of ischaemic stroke. Syst. Rev. 2013, 2, 33. [Google Scholar] [CrossRef] [PubMed]
- Kimura, T.; Horikoshi, Y.; Kuriyagawa, C.; Niiyama, Y. Rho/ROCK Pathway and Noncoding RNAs: Implications in Ischemic Stroke and Spinal Cord Injury. Int. J. Mol. Sci. 2021, 22, 11573. [Google Scholar] [CrossRef] [PubMed]
- Karalis, V.; Bateup, H.S. Current Approaches and Future Directions for the Treatment of mTORopathies. Dev. Neurosci. 2021, 43, 143–158. [Google Scholar] [CrossRef]
- Villa-González, M.; Martín-López, G.; Pérez-Álvarez, M.J. Dysregulation of mTOR Signaling after Brain Ischemia. Int. J. Mol. Sci. 2022, 23, 2814. [Google Scholar] [CrossRef] [PubMed]
- Kuriakose, D.; Xiao, Z. Pathophysiology and Treatment of Stroke: Raluca and Future Perspectives. Int. J. Mol. Sci. 2020, 21, 7609. [Google Scholar] [CrossRef] [PubMed]
- Haupt, M.; Gerner, S.T.; Bähr, M.; Doeppner, T.R. Neuroprotective Strategies for Ischemic Stroke-Future Perspectives. Int. J. Mol. Sci. 2023, 24, 4334. [Google Scholar] [CrossRef]
- Lakhan, S.E.; Kirchgessner, A.; Hofer, M. Inflammatory mechanisms in ischemic stroke: Therapeutic approaches. J. Transl. Med. 2009, 7, 97. [Google Scholar] [CrossRef]
- Shen, Z.; Xiang, M.; Chen, C.; Ding, F.; Wang, Y.; Shang, C.; Xin, L.; Zhang, Y.; Cui, X. Glutamate excitotoxicity: Potential therapeutic target for ischemic stroke. Biomed. Pharmacother. 2022, 151, 113125. [Google Scholar] [CrossRef] [PubMed]
- Rose, C.R.; Ziemens, D.; Verkhratsky, A. On the special role of NCX in astrocytes: Translating Na+-transients into intracellular Ca2+ signals. Cell Calcium 2020, 86, 102154. [Google Scholar] [CrossRef] [PubMed]
- Yan, J.; Bengtson, C.P.; Buchthal, B.; Hagenston, A.M.; Bading, H. Coupling of NMDA receptors and TRPM4 guides discovery of unconventional neuroprotectants. Science 2020, 370, eaay3302. [Google Scholar] [CrossRef] [PubMed]
- Stanika, R.I.; Villanueva, I.; Kazanina, G.; Andrews, S.B.; Pivovarova, N.B. Comparative impact of voltage-gated calcium channels and NMDA receptors on mitochondria-mediated neuronal injury. J. Neurosci. 2012, 32, 6642–6650. [Google Scholar] [CrossRef] [PubMed]
- Martel, M.A.; Ryan, T.J.; Bell, K.F.; Fowler, J.H.; McMahon, A.; Al-Mubarak, B.; Komiyama, N.H.; Horsburgh, K.; Kind, P.C.; Grant, S.G.; et al. The subtype of GluN2 C-terminal domain determines the response to excitotoxic insults. Neuron 2012, 74, 543–556. [Google Scholar] [CrossRef] [PubMed]
- He, J.; Liu, J.; Huang, Y.; Tang, X.; Xiao, H.; Hu, Z. Oxidative Stress, Inflammation, and Autophagy: Potential Targets of Mesenchymal Stem Cells-Based Therapies in Ischemic Stroke. Front. Neurosci. 2021, 15, 641157. [Google Scholar] [CrossRef] [PubMed]
- Nakka, V.P.; Prakash-Babu, P.; Vemuganti, R. Crosstalk between Endoplasmic Reticulum Stress, Oxidative Stress, and Autophagy: Potential Therapeutic Targets for Acute CNS Injuries. Mol. Neurobiol. 2016, 53, 532–544. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Zhang, X.; Chen, X.; Wei, Y. Neuronal injuries in cerebral infarction and ischemic stroke: From mechanisms to treatment (Review). Int. J. Mol. Med. 2022, 49, 15. [Google Scholar] [CrossRef] [PubMed]
- Pinoșanu, E.A.; Surugiu, R.; Burada, E.; Pîrșcoveanu, D.; Stănciulescu, C.E.; Sandu, R.E.; Pisoschi, C.; Albu, C.V. Oxidative Stress and Antioxidant Defense Mechanisms in Acute Ischemic Stroke Patients with Concurrent COVID-19 Infection. Int. J. Mol. Sci. 2023, 24, 16790. [Google Scholar] [CrossRef]
- Xiong, X.Y.; Liu, L.; Yang, Q.W. Functions and mechanisms of microglia/macrophages in neuroinflammation and neurogenesis after stroke. Prog. Neurobiol. 2016, 142, 23–44. [Google Scholar] [CrossRef]
- Meng, H.L.; Li, X.X.; Chen, Y.T.; Yu, L.J.; Zhang, H.; Lao, J.M.; Zhang, X.; Xu, Y. Neuronal Soluble Fas Ligand Drives M1-Microglia Polarization after Cerebral Ischemia. CNS Neurosci. Ther. 2016, 22, 771–781. [Google Scholar] [CrossRef] [PubMed]
- Jiang, C.T.; Wu, W.F.; Deng, Y.H.; Ge, J.W. Modulators of microglia activation and polarization in ischemic stroke (Review). Mol. Med. Rep. 2020, 21, 2006–2018. [Google Scholar] [CrossRef] [PubMed]
- Pekny, M.; Pekna, M. Astrocyte reactivity and reactive astrogliosis: Costs and benefits. Physiol. Rev. 2014, 94, 1077–1098. [Google Scholar] [CrossRef] [PubMed]
- Choudhury, G.R.; Ding, S. Reactive astrocytes and therapeutic potential in focal ischemic stroke. Neurobiol. Dis. 2016, 85, 234–244. [Google Scholar] [CrossRef]
- Ni, X.C.; Wang, H.F.; Cai, Y.Y.; Yang, D.; Alolga, R.N.; Liu, B.; Li, J.; Huang, F.Q. Ginsenoside Rb1 inhibits astrocyte activation and promotes transfer of astrocytic mitochondria to neurons against ischemic stroke. Redox Biol. 2022, 54, 102363. [Google Scholar] [CrossRef]
- Wang, Y.; Liu, W.; Geng, P.; Du, W.; Guo, C.; Wang, Q.; Zheng, G.Q.; Jin, X. Role of Crosstalk between Glial Cells and Immune Cells in Blood-Brain Barrier Damage and Protection after Acute Ischemic Stroke. Aging Dis. 2023. Epub ahead of print. [Google Scholar] [CrossRef]
- Qin, X.; Akter, F.; Qin, L.; Cheng, J.; Guo, M.; Yao, S.; Jian, Z.; Liu, R.; Wu, S. Adaptive Immunity Regulation and Cerebral Ischemia. Front. Immunol. 2020, 11, 689. [Google Scholar] [CrossRef]
- Surugiu, R.; Glavan, D.; Popescu, M.; Margaritescu, O.; Eugen, R.; Popa-Wagner, A. Vasculature Remodeling in a Rat Model of Cerebral Ischemia. The Fate of the BrdU-Labeled Cells Prior to Stroke. Front. Neurol. 2018, 9, 1014. [Google Scholar] [CrossRef]
- Iadecola, C.; Buckwalter, M.S.; Anrather, J. Immune responses to stroke: Mechanisms, modulation, and therapeutic potential. J. Clin. Investig. 2020, 130, 2777–2788. [Google Scholar] [CrossRef]
- Mosaddeghzadeh, N.; Ahmadian, M.R. The RHO Family GTPases: Mechanisms of Regulation and Signaling. Cells 2021, 10, 1831. [Google Scholar] [CrossRef]
- Jaffe, A.B.; Hall, A. Rho GTPases: Biochemistry and biology. Annu. Rev. Cell Dev. Biol. 2005, 21, 247–269. [Google Scholar] [CrossRef]
- Dvorsky, R.; Ahmadian, M.R. Always look on the bright site of Rho: Structural implications for a conserved intermolecular interface. EMBO Rep. 2004, 5, 1130–1136. [Google Scholar] [CrossRef]
- Sajib, M.S.; Zahra, F.T.; Akwii, R.G.; Mikelis, C.M. Identification of Rho GEF and RhoA Activation by Pull-Down Assays. Methods Mol. Biol. 2021, 2193, 97–109. [Google Scholar] [CrossRef]
- Chang, M.; Yi, L.; Zhou, Z.; Yi, X.; Chen, H.; Liang, X.; Jin, R.; Huang, X. GEF-H1/RhoA signaling pathway mediates pro-inflammatory effects of NF-κB on CD40L-induced pulmonary endothelial cells. Mol. Immunol. 2023, 157, 42–52. [Google Scholar] [CrossRef] [PubMed]
- Mason, F.M.; Xie, S.; Vasquez, C.G.; Tworoger, M.; Martin, A.C. RhoA GTPase inhibition organizes contraction during epithelial morphogenesis. J. Cell Biol. 2016, 214, 603–617. [Google Scholar] [CrossRef]
- Feng, Y.; LoGrasso, P.V.; Defert, O.; Li, R. Rho Kinase (ROCK) Inhibitors and Their Therapeutic Potential. J. Med. Chem. 2016, 59, 2269–2300. [Google Scholar] [CrossRef] [PubMed]
- Guan, G.; Cannon, R.D.; Coates, D.E.; Mei, L. Effect of the Rho-Kinase/ROCK Signaling Pathway on Cytoskeleton Components. Genes 2023, 14, 272. [Google Scholar] [CrossRef]
- Lee, S.R.; Xu, Y.N.; Jo, Y.J.; Namgoong, S.; Kim, N.H. The Rho-GTPase effector ROCK regulates meiotic maturation of the bovine oocyte via myosin light chain phosphorylation and cofilin phosphorylation. Mol. Reprod. Dev. 2015, 82, 849–858. [Google Scholar] [CrossRef] [PubMed]
- Hodge, R.G.; Ridley, A.J. Regulating Rho GTPases and their regulators. Nat. Rev. Mol. Cell Biol. 2016, 17, 496–510. [Google Scholar] [CrossRef] [PubMed]
- Wen, J.Y.; Gao, S.S.; Chen, F.L.; Chen, S.; Wang, M.; Chen, Z.W. Role of CSE-Produced H2S on Cerebrovascular Relaxation via RhoA-ROCK Inhibition and Cerebral Ischemia-Reperfusion Injury in Mice. ACS Chem. Neurosci. 2019, 10, 1565–1574. [Google Scholar] [CrossRef]
- Maldonado, H.; Calderon, C.; Burgos-Bravo, F.; Kobler, O.; Zuschratter, W.; Ramirez, O.; Härtel, S.; Schneider, P.; Quest, A.F.; Herrera-Molina, R.; et al. Astrocyte-to-neuron communication through integrin-engaged Thy-1/CBP/Csk/Src complex triggers neurite retraction via the RhoA/ROCK pathway. Biochim. Biophys. Acta Mol. Cell Res. 2017, 1864, 243–254. [Google Scholar] [CrossRef]
- Gao, J.; Liu, J.; Li, Y.; Liu, J.; Wang, H.; Chai, M.; Dong, Y.; Zhang, Z.; Su, G.; Wang, M. Targeting p53 for neuroinflammation: New therapeutic strategies in ischemic stroke. J. Neurosci. Res. 2023, 101, 1393–1408. [Google Scholar] [CrossRef] [PubMed]
- Sladojevic, N.; Yu, B.; Liao, J.K. ROCK as a therapeutic target for ischemic stroke. Expert Rev. Neurother. 2017, 17, 1167–1177. [Google Scholar] [CrossRef] [PubMed]
- Shin, H.K.; Salomone, S.; Potts, E.M.; Lee, S.W.; Millican, E.; Noma, K.; Huang, P.L.; Boas, D.A.; Liao, J.K.; Moskowitz, M.A.; et al. Rho-kinase inhibition acutely augments blood flow in focal cerebral ischemia via endothelial mechanisms. J. Cereb. Blood Flow Metab. 2007, 27, 998–1009. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Miao, L.; Peng, Q.; Fan, X.; Song, W.; Yang, B.; Zhang, P.; Liu, G.; Liu, J. Parthenolide modulates cerebral ischemia-induced microglial polarization and alleviates neuroinflammatory injury via the RhoA/ROCK pathway. Phytomedicine 2022, 105, 154373. [Google Scholar] [CrossRef] [PubMed]
- Rikitake, Y.; Kim, H.H.; Huang, Z.; Seto, M.; Yano, K.; Asano, T.; Moskowitz, M.A.; Liao, J.K. Inhibition of Rho kinase (ROCK) leads to increased cerebral blood flow and stroke protection. Stroke 2005, 36, 2251–2257. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.M.; Stalker, T.J.; Gong, Y.; Rikitake, Y.; Scalia, R.; Liao, J.K. Inhibition of Rho-kinase attenuates endothelial-leukocyte interaction during ischemia-reperfusion injury. Vasc. Med. 2012, 17, 379–385. [Google Scholar] [CrossRef]
- Shibuya, M.; Hirai, S.; Seto, M.; Satoh, S.; Ohtomo, E.; Fasudil Ischemic Stroke Study Group. Effects of fasudil in acute ischemic stroke: Results of a prospective placebo-controlled double-blind trial. J. Neurol. Sci. 2005, 238, 31–39. [Google Scholar] [CrossRef] [PubMed]
- Nowosad, A.; Jeannot, P.; Callot, C.; Creff, J.; Perchey, R.T.; Joffre, C.; Codogno, P.; Manenti, S.; Besson, A. p27 controls Ragulator and mTOR activity in amino acid-deprived cells to regulate the autophagy-lysosomal pathway and coordinate cell cycle and cell growth. Nat. Cell Biol. 2020, 22, 1076–1090, Erratum in Nat. Cell Biol. 2021, 23, 1048. [Google Scholar] [CrossRef]
- Yu, J.S.; Cui, W. Proliferation, survival and metabolism: The role of PI3K/AKT/mTOR signalling in pluripotency and cell fate determination. Development 2016, 143, 3050–3060. [Google Scholar] [CrossRef]
- Hung, C.M.; Garcia-Haro, L.; Sparks, C.A.; Guertin, D.A. mTOR-dependent cell survival mechanisms. Cold Spring Harb. Perspect. Biol. 2012, 4, a008771. [Google Scholar] [CrossRef]
- Luo, Y.; Xu, W.; Li, G.; Cui, W. Weighing In on mTOR Complex 2 Signaling: The Expanding Role in Cell Metabolism. Oxidative Med. Cell. Longev. 2018, 2018, 7838647. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.C.; Guan, K.L. mTOR: A pharmacologic target for autophagy regulation. J. Clin. Investig. 2015, 125, 25–32. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Zhang, H. Regulation of Autophagy by mTOR Signaling Pathway. Adv. Exp. Med. Biol. 2019, 1206, 67–83. [Google Scholar] [CrossRef] [PubMed]
- Yang, H.; Rudge, D.G.; Koos, J.D.; Vaidialingam, B.; Yang, H.J.; Pavletich, N.P. mTOR kinase structure, mechanism and regulation. Nature 2013, 497, 217–223. [Google Scholar] [CrossRef] [PubMed]
- Wang, P.; Shao, B.Z.; Deng, Z.; Chen, S.; Yue, Z.; Miao, C.Y. Autophagy in ischemic stroke. Prog. Neurobiol. 2018, 163–164, 98–117. [Google Scholar] [CrossRef] [PubMed]
- Sudarsanam, S.; Johnson, D.E. Functional consequences of mTOR inhibition. Curr. Opin. Drug Discov. Devel. 2010, 13, 31–40. [Google Scholar] [PubMed]
- Kim, J.; Kundu, M.; Viollet, B.; Guan, K.L. AMPK and mTOR regulate autophagy through direct phosphorylation of Ulk1. Nat. Cell Biol. 2011, 13, 132–141. [Google Scholar] [CrossRef] [PubMed]
- Cespedes, A.; Villa, M.; Benito-Cuesta, I.; Perez-Alvarez, M.J.; Ordoñez, L.; Wandosell, F. Energy-Sensing Pathways in Ischemia: The Counterbalance between AMPK and mTORC. Curr. Pharm. Des. 2019, 25, 4763–4770. [Google Scholar] [CrossRef]
- Hardie, D.G. AMP-activated/SNF1 protein kinases: Conserved guardians of cellular energy. Nat. Rev. Mol. Cell Biol. 2007, 8, 774–785. [Google Scholar] [CrossRef]
- Qin, C.; Yang, S.; Chu, Y.H.; Zhang, H.; Pang, X.W.; Chen, L.; Zhou, L.Q.; Chen, M.; Tian, D.S.; Wang, W. Signaling pathways involved in ischemic stroke: Molecular mechanisms and therapeutic interventions. Signal Transduct. Target. Ther. 2022, 7, 215, Erratum in Signal Transduct. Target. Ther. 2022, 7, 278. [Google Scholar] [CrossRef]
- Jiang, S.; Li, T.; Ji, T.; Yi, W.; Yang, Z.; Wang, S.; Yang, Y.; Gu, C. AMPK: Potential Therapeutic Target for Ischemic Stroke. Theranostics 2018, 8, 4535–4551. [Google Scholar] [CrossRef] [PubMed]
- Jiang, T.; Yu, J.T.; Zhu, X.C.; Zhang, Q.Q.; Tan, M.S.; Cao, L.; Wang, H.F.; Shi, J.Q.; Gao, L.; Qin, H.; et al. Ischemic preconditioning provides neuroprotection by induction of AMP-activated protein kinase-dependent autophagy in a rat model of ischemic stroke. Mol. Neurobiol. 2015, 51, 220–229. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Xue, X.; Zhang, H.; Che, X.; Luo, J.; Wang, P.; Xu, J.; Xing, Z.; Yuan, L.; Liu, Y.; et al. Neuronal-targeted TFEB rescues dysfunction of the autophagy-lysosomal pathway and alleviates ischemic injury in permanent cerebral ischemia. Autophagy 2019, 15, 493–509. [Google Scholar] [CrossRef]
- Pineda-Ramírez, N.; Gutiérrez Aguilar, G.F.; Espinoza-Rojo, M.; Aguilera, P. Current evidence for AMPK activation involvement on resveratrol-induced neuroprotection in cerebral ischemia. Nutr. Neurosci. 2018, 21, 229–247. [Google Scholar] [CrossRef]
- Mo, Y.; Sun, Y.Y.; Liu, K.Y. Autophagy and inflammation in ischemic stroke. Neural Regen. Res. 2020, 15, 1388–1396. [Google Scholar] [CrossRef]
- Kim, K.A.; Shin, D.; Kim, J.H.; Shin, Y.J.; Rajanikant, G.K.; Majid, A.; Baek, S.H.; Bae, O.N. Role of Autophagy in Endothelial Damage and Blood-Brain Barrier Disruption in Ischemic Stroke. Stroke 2018, 49, 1571–1579. [Google Scholar] [CrossRef] [PubMed]
- Zhang, D.M.; Zhang, T.; Wang, M.M.; Wang, X.X.; Qin, Y.Y.; Wu, J.; Han, R.; Sheng, R.; Wang, Y.; Chen, Z.; et al. TIGAR alleviates ischemia/reperfusion-induced autophagy and ischemic brain injury. Free. Radic. Biol. Med. 2019, 137, 13–23. [Google Scholar] [CrossRef]
- Fu, B.; Zeng, Q.; Zhang, Z.; Qian, M.; Chen, J.; Dong, W.; Li, M. Epicatechin Gallate Protects HBMVECs from Ischemia/Reperfusion Injury through Ameliorating Apoptosis and Autophagy and Promoting Neovascularization. Oxidative Med. Cell. Longev. 2019, 2019, 7824684. [Google Scholar] [CrossRef]
- Li, J.; McCullough, L.D. Effects of AMP-activated protein kinase in cerebral ischemia. J. Cereb. Blood Flow Metab. 2010, 30, 480–492. [Google Scholar] [CrossRef]
- Ashabi, G.; Khodagholi, F.; Khalaj, L.; Goudarzvand, M.; Nasiri, M. Activation of AMP-activated protein kinase by metformin protects against global cerebral ischemia in male rats: Interference of AMPK/PGC-1α pathway. Metab. Brain Dis. 2014, 29, 47–58. [Google Scholar] [CrossRef]
- Zhao, M.; Li, X.W.; Chen, Z.; Hao, F.; Tao, S.X.; Yu, H.Y.; Cheng, R.; Liu, H. Neuro-Protective Role of Metformin in Patients with Acute Stroke and Type 2 Diabetes Mellitus via AMPK/Mammalian Target of Rapamycin (mTOR) Signaling Pathway and Oxidative Stress. Med. Sci. Monit. 2019, 25, 2186–2194. [Google Scholar] [CrossRef] [PubMed]
- Wang, P.F.; Fang, H.; Chen, J.; Lin, S.; Liu, Y.; Xiong, X.Y.; Wang, Y.C.; Xiong, R.P.; Lv, F.L.; Wang, J.; et al. Polyinosinic-polycytidylic acid has therapeutic effects against cerebral ischemia/reperfusion injury through the downregulation of TLR4 signaling via TLR3. J. Immunol. 2014, 192, 4783–4794. [Google Scholar] [CrossRef] [PubMed]
- Ma, X.; Xiao, W.; Li, H.; Pang, P.; Xue, F.; Wan, L.; Pei, L.; Yan, H. Metformin restores hippocampal neurogenesis and learning and memory via regulating gut microbiota in the obese mouse model. Brain Behav. Immun. 2021, 95, 68–83. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Tang, G.; Zhang, Z.; Wang, Y.; Yang, G.Y. Metformin promotes focal angiogenesis and neurogenesis in mice following middle cerebral artery occlusion. Neurosci. Lett. 2014, 579, 46–51. [Google Scholar] [CrossRef] [PubMed]
- Venna, V.R.; Li, J.; Hammond, M.D.; Mancini, N.S.; McCullough, L.D. Chronic metformin treatment improves post-stroke angiogenesis and recovery after experimental stroke. Eur. J. Neurosci. 2014, 39, 2129–2138. [Google Scholar] [CrossRef] [PubMed]
- Yuan, R.; Wang, Y.; Li, Q.; Zhen, F.; Li, X.; Lai, Q.; Hu, P.; Wang, X.; Zhu, Y.; Fan, H.; et al. Metformin reduces neuronal damage and promotes neuroblast proliferation and differentiation in a cerebral ischemia/reperfusion rat model. Neuroreport 2019, 30, 232–240. [Google Scholar] [CrossRef] [PubMed]
- Farbood, Y.; Sarkaki, A.; Khalaj, L.; Khodagholi, F.; Badavi, M.; Ashabi, G. Targeting Adenosine Monophosphate-Activated Protein Kinase by Metformin Adjusts Post-Ischemic Hyperemia and Extracellular Neuronal Discharge in Transient Global Cerebral Ischemia. Microcirculation 2015, 22, 534–541. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Zhang, J.; Zhu, X.; Jiao, W.; Yang, Y.; Wu, Y.; Yang, L.; Wang, Y. Metformin enhances neural precursor cells migration and functional recovery after ischemic stroke in mice. Exp. Brain Res. 2023, 241, 505–515. [Google Scholar] [CrossRef] [PubMed]
- Zhu, X.C.; Jiang, T.; Zhang, Q.Q.; Cao, L.; Tan, M.S.; Wang, H.F.; Ding, Z.Z.; Tan, L.; Yu, J.T. Chronic Metformin Preconditioning Provides Neuroprotection via Suppression of NF-κB-Mediated Inflammatory Pathway in Rats with Permanent Cerebral Ischemia. Mol. Neurobiol. 2015, 52, 375–385. [Google Scholar] [CrossRef]
- Ghadernezhad, N.; Khalaj, L.; Pazoki-Toroudi, H.; Mirmasoumi, M.; Ashabi, G. Metformin pretreatment enhanced learning and memory in cerebral forebrain ischaemia: The role of the AMPK/BDNF/P70SK signalling pathway. Pharm. Biol. 2016, 54, 2211–2219. [Google Scholar] [CrossRef]
- Li, J.; Benashski, S.E.; Venna, V.R.; McCullough, L.D. Effects of metformin in experimental stroke. Stroke 2010, 41, 2645–2652. [Google Scholar] [CrossRef]
- Sharma, S.; Nozohouri, S.; Vaidya, B.; Abbruscato, T. Repurposing metformin to treat age-related neurodegenerative disorders and ischemic stroke. Life Sci. 2021, 274, 119343. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Z.; Xiao, J.; Wang, J.; Dong, W.; Peng, Z.; An, D. Anti-inflammatory effects of novel sinomenine derivatives. Int. Immunopharmacol. 2015, 29, 354–360. [Google Scholar] [CrossRef]
- Qiu, J.; Wang, M.; Zhang, J.; Cai, Q.; Lu, D.; Li, Y.; Dong, Y.; Zhao, T.; Chen, H. The neuroprotection of Sinomenine against ischemic stroke in mice by suppressing NLRP3 inflammasome via AMPK signaling. Int. Immunopharmacol. 2016, 40, 492–500. [Google Scholar] [CrossRef] [PubMed]
- Qiu, J.; Yan, Z.; Tao, K.; Li, Y.; Li, Y.; Li, J.; Dong, Y.; Feng, D.; Chen, H. Sinomenine activates astrocytic dopamine D2 receptors and alleviates neuroinflammatory injury via the CRYAB/STAT3 pathway after ischemic stroke in mice. J. Neuroinflamm. 2016, 13, 263. [Google Scholar] [CrossRef]
- Bi, F.; Zhang, Y.; Liu, W.; Xie, K. Sinomenine activation of Nrf2 signaling prevents inflammation and cerebral injury in a mouse model of ischemic stroke. Exp. Ther. Med. 2021, 21, 647. [Google Scholar] [CrossRef]
- Walle, T. Bioavailability of resveratrol. Ann. N. Y. Acad. Sci. 2011, 1215, 9–15. [Google Scholar] [CrossRef] [PubMed]
- Gutiérrez Aguilar, G.F.; Alquisiras-Burgos, I.; Franco-Pérez, J.; Pineda-Ramírez, N.; Ortiz-Plata, A.; Torres, I.; Pedraza-Chaverri, J.; Aguilera, P. Resveratrol Prevents GLUT3 Up-Regulation Induced by Middle Cerebral Artery Occlusion. Brain Sci. 2020, 10, 651. [Google Scholar] [CrossRef]
- Park, D.J.; Kang, J.B.; Shah, F.A.; Koh, P.O. Resveratrol modulates the Akt/GSK-3β signaling pathway in a middle cerebral artery occlusion animal model. Lab. Anim. Res. 2019, 35, 18. [Google Scholar] [CrossRef]
- Ashafaq, M.; Intakhab Alam, M.; Khan, A.; Islam, F.; Khuwaja, G.; Hussain, S.; Ali, R.; Alshahrani, S.; Antar Makeen, H.; Alhazmi, H.A.; et al. Nanoparticles of resveratrol attenuates oxidative stress and inflammation after ischemic stroke in rats. Int. Immunopharmacol. 2021, 94, 107494. [Google Scholar] [CrossRef]
- Liao, H.; Huang, J.; Liu, J.; Chen, Y.; Zhu, H.; Li, X.; Wen, J.; Xiang, Q.; Yang, Q. Resveratrol Inhibits Activation of Microglia after Stroke through Triggering Translocation of Smo to Primary Cilia. J. Pers. Med. 2023, 13, 268. [Google Scholar] [CrossRef]
- Wan, D.; Zhou, Y.; Wang, K.; Hou, Y.; Hou, R.; Ye, X. Resveratrol provides neuroprotection by inhibiting phosphodiesterases and regulating the cAMP/AMPK/SIRT1 pathway after stroke in rats. Brain Res. Bull. 2016, 121, 255–262. [Google Scholar] [CrossRef]
- Pineda-Ramírez, N.; Alquisiras-Burgos, I.; Ortiz-Plata, A.; Ruiz-Tachiquín, M.E.; Espinoza-Rojo, M.; Aguilera, P. Resveratrol Activates Neuronal Autophagy Through AMPK in the Ischemic Brain. Mol. Neurobiol. 2020, 57, 1055–1069. [Google Scholar] [CrossRef]
- Hou, Y.; Wang, K.; Wan, W.; Cheng, Y.; Pu, X.; Ye, X. Resveratrol provides neuroprotection by regulating the JAK2/STAT3/PI3K/AKT/mTOR pathway after stroke in rats. Genes Dis. 2018, 5, 245–255. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; He, J.; Huang, Y.; Hu, Z. Resveratrol has an Overall Neuroprotective Role in Ischemic Stroke: A Meta-Analysis in Rodents. Front. Pharmacol. 2021, 12, 795409. [Google Scholar] [CrossRef] [PubMed]
- Fodor, K.; Tit, D.M.; Pasca, B.; Bustea, C.; Uivarosan, D.; Endres, L.; Iovan, C.; Abdel-Daim, M.M.; Bungau, S. Long-Term Resveratrol Supplementation as a Secondary Prophylaxis for Stroke. Oxidative Med. Cell. Longev. 2018, 2018, 4147320. [Google Scholar] [CrossRef]
- Chen, J.; Bai, Q.; Zhao, Z.; Sui, H.; Xie, X. Resveratrol improves delayed r-tPA treatment outcome by reducing MMPs. Acta Neurol. Scand. 2016, 134, 54–60. [Google Scholar] [CrossRef]
- Papadia, S.; Hardingham, G.E. The dichotomy of NMDA receptor signaling. Neuroscientist 2007, 13, 572–579. [Google Scholar] [CrossRef] [PubMed]
- Hardingham, G.E. Coupling of the NMDA receptor to neuroprotective and neurodestructive events. Biochem. Soc. Trans. 2009, 37, 1147–1160. [Google Scholar] [CrossRef]
- Wu, Q.J.; Tymianski, M. Targeting NMDA receptors in stroke: New hope in neuroprotection. Mol. Brain 2018, 11, 15. [Google Scholar] [CrossRef]
- Lai, T.W.; Zhang, S.; Wang, Y.T. Excitotoxicity and stroke: Identifying novel targets for neuroprotection. Prog. Neurobiol. 2014, 115, 157–188. [Google Scholar] [CrossRef] [PubMed]
- Cook, D.J.; Teves, L.; Tymianski, M. Treatment of stroke with a PSD-95 inhibitor in the gyrencephalic primate brain. Nature 2012, 483, 213–217. [Google Scholar] [CrossRef] [PubMed]
- Hill, M.D.; Martin, R.H.; Mikulis, D.; Wong, J.H.; Silver, F.L.; Terbrugge, K.G.; Milot, G.; Clark, W.M.; Macdonald, R.L.; Kelly, M.E.; et al. Safety and efficacy of NA-1 in patients with iatrogenic stroke after endovascular aneurysm repair (ENACT): A phase 2, randomised, double-blind, placebo-controlled trial. Lancet Neurol. 2012, 11, 942–950. [Google Scholar] [CrossRef] [PubMed]
- Basto, F.M.; Lyden, P. Hypothermia in acute ischemic stroke therapy. Handb. Clin. Neurol. 2018, 157, 823–837. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Benashski, S.; McCullough, L.D. Post-stroke hypothermia provides neuroprotection through inhibition of AMP-activated protein kinase. J. Neurotrauma 2011, 28, 1281–1288. [Google Scholar] [CrossRef] [PubMed]
- Sandu, R.E.; Dumbrava, D.; Surugiu, R.; Glavan, D.G.; Gresita, A.; Petcu, E.B. Cellular and Molecular Mechanisms Underlying Non-Pharmaceutical Ischemic Stroke Therapy in Aged Subjects. Int. J. Mol. Sci. 2017, 19, 99. [Google Scholar] [CrossRef] [PubMed]
- Sandu, R.E.; Uzoni, A.; Ciobanu, O.; Moldovan, M.; Anghel, A.; Radu, E.; Coogan, A.N.; Popa-Wagner, A. Post-stroke gaseous hypothermia increases vascular density but not neurogenesis in the ischemic penumbra of aged rats. Restor. Neurol. Neurosci. 2016, 34, 401–414. [Google Scholar] [CrossRef] [PubMed]
- Guan, L.; Lee, H.; Geng, X.; Li, F.; Shen, J.; Ji, Y.; Peng, C.; Du, H.; Ding, Y. Neuroprotective Effects of Pharmacological Hypothermia on Hyperglycolysis and Gluconeogenesis in Rats after Ischemic Stroke. Biomolecules 2022, 12, 851. [Google Scholar] [CrossRef] [PubMed]
- Han, Y.; Geng, X.K.; Lee, H.; Li, F.; Ding, Y. Neuroprotective Effects of Early Hypothermia Induced by Phenothiazines and DHC in Ischemic Stroke. Evid.-Based Complement. Altern. Med. 2021, 2021, 1207092. [Google Scholar] [CrossRef]
- Sandu, R.E.; Buga, A.M.; Balseanu, A.T.; Moldovan, M.; Popa-Wagner, A. Twenty-four hours hypothermia has temporary efficacy in reducing brain infarction and inflammation in aged rats. Neurobiol. Aging 2016, 38, 127–140. [Google Scholar] [CrossRef]
- Zeng, Y.; Hao, L.; Chen, Y.; Liu, S.; Fan, Y.; Zhao, Z.; Wang, Y.; Chen, Q.; Li, Y. Optimizing intra-arterial hypothermia scheme for acute ischemic stroke in an MCAO/R rat model. Sci. Rep. 2023, 13, 9566. [Google Scholar] [CrossRef] [PubMed]
- Kuczynski, A.M.; Marzoughi, S.; Al Sultan, A.S.; Colbourne, F.; Menon, B.K.; van Es, A.C.G.M.; Berez, A.L.; Goyal, M.; Demchuk, A.M.; Almekhlafi, M.A. Therapeutic Hypothermia in Acute Ischemic Stroke—A Systematic Review and Meta-Analysis. Curr. Neurol. Neurosci. Rep. 2020, 20, 13. [Google Scholar] [CrossRef] [PubMed]
- Lyden, P.; Hemmen, T.; Grotta, J.; Rapp, K.; Ernstrom, K.; Rzesiewicz, T.; Parker, S.; Concha, M.; Hussain, S.; Agarwal, S.; et al. Results of the ICTuS 2 Trial (Intravascular Cooling in the Treatment of Stroke 2). Stroke 2016, 47, 2888–2895. [Google Scholar] [CrossRef]
- van der Worp, H.B.; Macleod, M.R.; Bath, P.M.; Bathula, R.; Christensen, H.; Colam, B.; Cordonnier, C.; Demotes-Mainard, J.; Durand-Zaleski, I.; Gluud, C.; et al. Therapeutic hypothermia for acute ischaemic stroke. Results of a European multicentre, randomised, phase III clinical trial. Eur. Stroke J. 2019, 4, 254–262. [Google Scholar] [CrossRef] [PubMed]
- Palà, E.; Penalba, A.; Bustamante, A.; García-Berrocoso, T.; Lamana-Vallverdú, M.; Meisel, C.; Meisel, A.; van der Worp, H.B.; RMacleod, M.; Kallmünzer, B.; et al. Blood biomarker changes following therapeutic hypothermia in ischemic stroke. Brain Behav. 2023, 13, e3230. [Google Scholar] [CrossRef]
- Jiao, C.; Liu, C.; Yang, Z.; Jin, C.; Chen, X.; Xue, J.; Zhang, G.; Pan, C.; Jia, J.; Hou, X. Brain Protection Effects of Mild Hypothermia Combined with Distant Ischemic Postconditioning and Thrombolysis in Patients with Acute Ischemic Stroke. Ther. Hypothermia Temp. Manag. 2023. Epub ahead of print. [Google Scholar] [CrossRef]
- Ferreira, J.R.; Teixeira, G.Q.; Santos, S.G.; Barbosa, M.A.; Almeida-Porada, G.; Gonçalves, R.M. Mesenchymal Stromal Cell Secretome: Influencing Therapeutic Potential by Cellular Pre-conditioning. Front. Immunol. 2018, 9, 2837. [Google Scholar] [CrossRef]
- Műzes, G.; Sipos, F. Mesenchymal Stem Cell-Derived Secretome: A Potential Therapeutic Option for Autoimmune and Immune-Mediated Inflammatory Diseases. Cells 2022, 11, 2300. [Google Scholar] [CrossRef]
- Ben-David, U.; Mayshar, Y.; Benvenisty, N. Large-scale analysis reveals acquisition of lineage-specific chromosomal aberrations in human adult stem cells. Cell Stem Cell 2011, 9, 97–102. [Google Scholar] [CrossRef] [PubMed]
- Squillaro, T.; Peluso, G.; Galderisi, U. Clinical Trials with Mesenchymal Stem Cells: An Update. Cell Transplant. 2016, 25, 829–848. [Google Scholar] [CrossRef]
- Kumar, P.; Kandoi, S.; Misra, R.; Vijayalakshmi, S.; Rajagopal, K.; Verma, R.S. The mesenchymal stem cell secretome: A new paradigm towards cell-free therapeutic mode in regenerative medicine. Cytokine Growth Factor Rev. 2019, 46, 1–9. [Google Scholar] [CrossRef]
- Zagrean, A.M.; Hermann, D.M.; Opris, I.; Zagrean, L.; Popa-Wagner, A. Multicellular Crosstalk between Exosomes and the Neurovascular Unit after Cerebral Ischemia. Therapeutic Implications. Front. Neurosci. 2018, 12, 811. [Google Scholar] [CrossRef] [PubMed]
- Fu, Y.; Karbaat, L.; Wu, L.; Leijten, J.; Both, S.K.; Karperien, M. Trophic Effects of Mesenchymal Stem Cells in Tissue Regeneration. Tissue Eng. Part B Rev. 2017, 23, 515–528. [Google Scholar] [CrossRef]
- Surugiu, R.; Olaru, A.; Hermann, D.M.; Glavan, D.; Catalin, B.; Popa-Wagner, A. Recent Advances in Mono- and Combined Stem Cell Therapies of Stroke in Animal Models and Humans. Int. J. Mol. Sci. 2019, 20, 6029. [Google Scholar] [CrossRef] [PubMed]
- Chang, C.; Yan, J.; Yao, Z.; Zhang, C.; Li, X.; Mao, H.Q. Effects of Mesenchymal Stem Cell-Derived Paracrine Signals and Their Delivery Strategies. Adv. Healthc. Mater. 2021, 10, e2001689. [Google Scholar] [CrossRef] [PubMed]
- Martins, L.F.; Costa, R.O.; Pedro, J.R.; Aguiar, P.; Serra, S.C.; Teixeira, F.G.; Sousa, N.; Salgado, A.J.; Almeida, R.D. Mesenchymal stem cells secretome-induced axonal outgrowth is mediated by BDNF. Sci. Rep. 2017, 7, 4153. [Google Scholar] [CrossRef] [PubMed]
- Teixeira, F.G.; Carvalho, M.M.; Neves-Carvalho, A.; Panchalingam, K.M.; Behie, L.A.; Pinto, L.; Sousa, N.; Salgado, A.J. Secretome of mesenchymal progenitors from the umbilical cord acts as modulator of neural/glial proliferation and differentiation. Stem Cell Rev. Rep. 2015, 11, 288–297. [Google Scholar] [CrossRef]
- Kemp, K.; Hares, K.; Mallam, E.; Heesom, K.J.; Scolding, N.; Wilkins, A. Mesenchymal stem cell-secreted superoxide dismutase promotes cerebellar neuronal survival. J. Neurochem. 2010, 114, 1569–1580. [Google Scholar] [CrossRef]
- Shen, C.; Lie, P.; Miao, T.; Yu, M.; Lu, Q.; Feng, T.; Li, J.; Zu, T.; Liu, X.; Li, H. Conditioned medium from umbilical cord mesenchymal stem cells induces migration and angiogenesis. Mol. Med. Rep. 2015, 12, 20–30. [Google Scholar] [CrossRef] [PubMed]
- Bi, Y.; Qiao, X.; Liu, Q.; Song, S.; Zhu, K.; Qiu, X.; Zhang, X.; Jia, C.; Wang, H.; Yang, Z.; et al. Systemic proteomics and miRNA profile analysis of exosomes derived from human pluripotent stem cells. Stem Cell Res. Ther. 2022, 13, 449. [Google Scholar] [CrossRef]
- da Silva, A.V.; Serrenho, I.; Araújo, B.; Carvalho, A.M.; Baltazar, G. Secretome as a Tool to Treat Neurological Conditions: Are We Ready? Int. J. Mol. Sci. 2023, 24, 16544. [Google Scholar] [CrossRef]
- Hermann, D.M.; Xin, W.; Bähr, M.; Giebel, B.; Doeppner, T.R. Emerging roles of extracellular vesicle-associated non-coding RNAs in hypoxia: Insights from cancer, myocardial infarction and ischemic stroke. Theranostics 2022, 12, 5776–5802. [Google Scholar] [CrossRef] [PubMed]
- Xu, R.; Bai, Y.; Min, S.; Xu, X.; Tang, T.; Ju, S. In Vivo Monitoring and Assessment of Exogenous Mesenchymal Stem Cell-Derived Exosomes in Mice with Ischemic Stroke by Molecular Imaging. Int. J. NanoMed. 2020, 15, 9011–9023. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Zhang, M.; Liu, H.; Zhu, R.; He, H.; Zhou, Y.; Zhang, Y.; Li, C.; Liang, D.; Zeng, Q.; et al. Bone marrow mesenchymal stem cell-derived exosomes attenuate cerebral ischemia-reperfusion injury-induced neuroinflammation and pyroptosis by modulating microglia M1/M2 phenotypes. Exp. Neurol. 2021, 341, 113700. [Google Scholar] [CrossRef]
- Wang, C.; Börger, V.; Mohamud Yusuf, A.; Tertel, T.; Stambouli, O.; Murke, F.; Freund, N.; Kleinschnitz, C.; Herz, J.; Gunzer, M.; et al. Postischemic Neuroprotection Associated with Anti-Inflammatory Effects by Mesenchymal Stromal Cell-Derived Small Extracellular Vesicles in Aged Mice. Stroke 2022, 53, e14–e18. [Google Scholar] [CrossRef] [PubMed]
- Doeppner, T.R.; Herz, J.; Görgens, A.; Schlechter, J.; Ludwig, A.K.; Radtke, S.; de Miroschedji, K.; Horn, P.A.; Giebel, B.; Hermann, D.M. Extracellular Vesicles Improve Post-Stroke Neuroregeneration and Prevent Postischemic Immunosuppression. Stem Cells Transl. Med. 2015, 4, 1131–1143. [Google Scholar] [CrossRef] [PubMed]
- Dumbrava, D.A.; Surugiu, R.; Börger, V.; Ruscu, M.; Tertel, T.; Giebel, B.; Hermann, D.M.; Popa-Wagner, A. Mesenchymal stromal cell-derived small extracellular vesicles promote neurological recovery and brain remodeling after distal middle cerebral artery occlusion in aged rats. Geroscience 2022, 44, 293–310. [Google Scholar] [CrossRef] [PubMed]
- Xin, H.; Li, Y.; Liu, Z.; Wang, X.; Shang, X.; Cui, Y.; Zhang, Z.G.; Chopp, M. MiR-133b promotes neural plasticity and functional recovery after treatment of stroke with multipotent mesenchymal stromal cells in rats via transfer of exosome-enriched extracellular particles. Stem Cells 2013, 31, 2737–2746. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Zhu, T.; Li, H.; Zhao, J.; Li, X. Transplantation of Lymphocytes Co-Cultured with Human Cord Blood-Derived Multipotent Stem Cells Attenuates Inflammasome Activity in Ischemic Stroke. Clin. Interv. Aging 2019, 14, 2261–2271. [Google Scholar] [CrossRef]
- He, H.; Zeng, Q.; Huang, G.; Lin, Y.; Lin, H.; Liu, W.; Lu, P. Bone marrow mesenchymal stem cell transplantation exerts neuroprotective effects following cerebral ischemia/reperfusion injury by inhibiting autophagy via the PI3K/Akt pathway. Brain Res. 2019, 1707, 124–132. [Google Scholar] [CrossRef]
- Nazarinia, D.; Aboutaleb, N.; Gholamzadeh, R.; Nasseri Maleki, S.; Mokhtari, B.; Nikougoftar, M. Conditioned medium obtained from human amniotic mesenchymal stem cells attenuates focal cerebral ischemia/reperfusion injury in rats by targeting mTOR pathway. J. Chem. Neuroanat. 2019, 102, 101707. [Google Scholar] [CrossRef]
- Xin, H.; Liu, Z.; Buller, B.; Li, Y.; Golembieski, W.; Gan, X.; Wang, F.; Lu, M.; Ali, M.M.; Zhang, Z.G.; et al. MiR-17-92 enriched exosomes derived from multipotent mesenchymal stromal cells enhance axon-myelin remodeling and motor electrophysiological recovery after stroke. J. Cereb. Blood Flow Metab. 2021, 41, 1131–1144. [Google Scholar] [CrossRef]
- Kuang, Y.; Zheng, X.; Zhang, L.; Ai, X.; Venkataramani, V.; Kilic, E.; Hermann, D.M.; Majid, A.; Bähr, M.; Doeppner, T.R. Adipose-derived mesenchymal stem cells reduce autophagy in stroke mice by extracellular vesicle transfer of miR-25. J. Extracell. Vesicles 2020, 10, e12024. [Google Scholar] [CrossRef]
- Zeng, Q.; Zhou, Y.; Liang, D.; He, H.; Liu, X.; Zhu, R.; Zhang, M.; Luo, X.; Wang, Y.; Huang, G. Exosomes Secreted from Bone Marrow Mesenchymal Stem Cells Attenuate Oxygen-Glucose Deprivation/Reoxygenation-Induced Pyroptosis in PC12 Cells by Promoting AMPK-Dependent Autophagic Flux. Front. Cell. Neurosci. 2020, 14, 182. [Google Scholar] [CrossRef]
- Zheng, Z.; Zhang, L.; Qu, Y.; Xiao, G.; Li, S.; Bao, S.; Lu, Q.R.; Mu, D. Mesenchymal Stem Cells Protect against Hypoxia-Ischemia Brain Damage by Enhancing Autophagy through Brain Derived Neurotrophic Factor/Mammalin Target of Rapamycin Signaling Pathway. Stem Cells 2018, 36, 1109–1121. [Google Scholar] [CrossRef] [PubMed]
- Dahbour, S.; Jamali, F.; Alhattab, D.; Al-Radaideh, A.; Ababneh, O.; Al-Ryalat, N.; Al-Bdour, M.; Hourani, B.; Msallam, M.; Rasheed, M.; et al. Mesenchymal stem cells and conditioned media in the treatment of multiple sclerosis patients: Clinical, ophthalmological and radiological assessments of safety and efficacy. CNS Neurosci. Ther. 2017, 23, 866–874. [Google Scholar] [CrossRef]
- Mulherkar, S.; Tolias, K.F. RhoA-ROCK Signaling as a Therapeutic Target in Traumatic Brain Injury. Cells 2020, 9, 245. [Google Scholar] [CrossRef] [PubMed]
- Glotfelty, E.J.; Tovar-Y-Romo, L.B.; Hsueh, S.C.; Tweedie, D.; Li, Y.; Harvey, B.K.; Hoffer, B.J.; Karlsson, T.E.; Olson, L.; Greig, N.H. The RhoA-ROCK1/ROCK2 Pathway Exacerbates Inflammatory Signaling in Immortalized and Primary Microglia. Cells 2023, 12, 1367. [Google Scholar] [CrossRef]
- Lee, J.H.; Zheng, Y.; von Bornstadt, D.; Wei, Y.; Balcioglu, A.; Daneshmand, A.; Yalcin, N.; Yu, E.; Herisson, F.; Atalay, Y.B.; et al. Selective ROCK2 Inhibition in Focal Cerebral Ischemia. Ann. Clin. Transl. Neurol. 2014, 1, 2–14. [Google Scholar] [CrossRef] [PubMed]
- Guan, X.; Wei, D.; Liang, Z.; Xie, L.; Wang, Y.; Huang, Z.; Wu, J.; Pang, T. FDCA Attenuates Neuroinflammation and Brain Injury after Cerebral Ischemic Stroke. ACS Chem. Neurosci. 2023, 14, 3839–3854. [Google Scholar] [CrossRef]
- Fang, F.; Sheng, J.; Guo, Y.; Wen, J.; Chen, Z. Protection of H2S against Hypoxia/Reoxygenation Injury in Rat Hippocampal Neurons through Inhibiting Phosphorylation of ROCK2 at Thr436 and Ser575. Pharmaceuticals 2023, 16, 218. [Google Scholar] [CrossRef]
- Yang, X.; Wang, G. Fasudil mediates neuroprotection in ischemia/reperfusion by modulating the ROCK-PPARα-NOX axis. Acta Cir. Bras. 2023, 38, e387023. [Google Scholar] [CrossRef]
- Morita, M.; Gravel, S.P.; Hulea, L.; Larsson, O.; Pollak, M.; St-Pierre, J.; Topisirovic, I. mTOR coordinates protein synthesis, mitochondrial activity and proliferation. Cell Cycle 2015, 14, 473–480. [Google Scholar] [CrossRef]
- Cao, W.; Li, J.; Yang, K.; Cao, D. An overview of autophagy: Mechanism, regulation and research progress. Bull. Cancer 2021, 108, 304–322. [Google Scholar] [CrossRef]
- Gao, J.; Yao, M.; Chang, D.; Liu, J. mTOR (Mammalian Target of Rapamycin): Hitting the Bull’s Eye for Enhancing Neurogenesis after Cerebral Ischemia? Stroke 2023, 54, 279–285. [Google Scholar] [CrossRef]
- Zoungrana, L.I.; Krause-Hauch, M.; Wang, H.; Fatmi, M.K.; Bates, L.; Li, Z.; Kulkarni, P.; Ren, D.; Li, J. The Interaction of mTOR and Nrf2 in Neurogenesis and Its Implication in Neurodegenerative Diseases. Cells 2022, 11, 2048. [Google Scholar] [CrossRef]
- Pallet, N.; Legendre, C. Adverse events associated with mTOR inhibitors. Expert Opin. Drug Saf. 2013, 12, 177–186. [Google Scholar] [CrossRef]
- Li, F.; Zhang, J.; Yi, K.; Wang, H.; Wei, H.; Chan, H.F.; Tao, Y.; Li, M. Delivery of Stem Cell Secretome for Therapeutic Applications. ACS Appl. Bio Mater. 2022, 5, 2009–2030. [Google Scholar] [CrossRef]
- Hermann, D.M.; Peruzzotti-Jametti, L.; Giebel, B.; Pluchino, S. Extracellular vesicles set the stage for brain plasticity and recovery by multimodal signalling. Brain 2024, 147, 372–389. [Google Scholar] [CrossRef]
- Liu, W.; Wang, X.; O’Connor, M.; Wang, G.; Han, F. Brain-Derived Neurotrophic Factor and Its Potential Therapeutic Role in Stroke Comorbidities. Neural Plast. 2020, 2020, 1969482. [Google Scholar] [CrossRef]
- Downer, M.B.; Li, L.; Carter, S.; Beebe, S.; Rothwell, P.M. Associations of Multimorbidity with Stroke Severity, Subtype, Premorbid Disability, and Early Mortality: Oxford Vascular Study. Neurology 2023, 101, e645–e652. [Google Scholar] [CrossRef]
- Rutten-Jacobs, L.C.; Larsson, S.C.; Malik, R.; Rannikmäe, K.; Sudlow, C.L.; Dichgans, M.; Markus, H.S.; Traylor, M.; International Stroke Genetics Consortium. Genetic risk, incident stroke, and the benefits of adhering to a healthy lifestyle: Cohort study of 306 473 UK Biobank participants. BMJ 2018, 363, k4168. [Google Scholar] [CrossRef]
- Popa-Wagner, A.; Dumitrascu, D.I.; Capitanescu, B.; Petcu, E.B.; Surugiu, R.; Fang, W.H.; Dumbrava, D.A. Dietary habits, lifestyle factors and neurodegenerative diseases. Neural Regen. Res. 2020, 15, 394–400. [Google Scholar] [CrossRef]
- Albu, V.C.; Sandu, R.E.; Barbulescu, A.L.; Tartea, E.A.; Burada, E.; Taisescu, O.; Vreju, F.A.; Boldeanu, M.V.; Boldeanu, L.; Enescu, A.; et al. The Relationship between the Acute Confusing State and the Comorbidities of the Elderly Patient. Rev. Chim. 2019, 70, 2415–2419. [Google Scholar] [CrossRef]
- Popa-Wagner, A.; Glavan, D.G.; Olaru, A.; Olaru, D.G.; Margaritescu, O.; Tica, O.; Surugiu, R.; Sandu, R.E. Present Status and Future Challenges of New Therapeutic Targets in Preclinical Models of Stroke in Aged Animals with/without Comorbidities. Int. J. Mol. Sci. 2018, 19, 356. [Google Scholar] [CrossRef]
- Sandu, R.E.; Uzoni, A.; Coman, C.; Popa-Wagner, A. Cerebral ischemia in the aged. Limited anti-inflammatory efficacy of the indomethacin treatment. Rom. J. Morphol. Embryol. 2015, 56, 1111–1117. [Google Scholar]
- Popa-Wagner, A.; Sandu, R.E.; Cristin, C.; Uzoni, A.; Welle, K.A.; Hryhorenko, J.R.; Ghaemmaghami, S. Increased Degradation Rates in the Components of the Mitochondrial Oxidative Phosphorylation Chain in the Cerebellum of Old Mice. Front. Aging Neurosci. 2018, 10, 32. [Google Scholar] [CrossRef]
- Sandu, R.E.; Balseanu, A.T.; Bogdan, C.; Slevin, M.; Petcu, E.; Popa-Wagner, A. Stem cell therapies in preclinical models of stroke. Is the aged brain microenvironment refractory to cell therapy? Exp. Gerontol. 2017, 94, 73–77. [Google Scholar] [CrossRef]
- Buga, A.M.; Vintilescu, R.; Pop, O.T.; Popa-Wagner, A. Brain Aging and Regeneration after Injuries: An Organismal approach. Aging Dis. 2011, 2, 64–79. [Google Scholar]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Therapy | Targeting Pathway | Model | Species | Results | ||
|---|---|---|---|---|---|---|
| RhoA/ROCK | Fasudil | ROCK inhibition | MCAO | Mice | ↓ infarct size ↑ neurological deficit | [56] |
| Fasudil | ROCK inhibition | Hemorrhage/reinfusion | Mice | ↑ learning capacity | [57] | |
| Fasudil | ROCK inhibition | Ischemic stroke | Humans | ↑ neurological status ↑ clinical outcome | [58] | |
| mTOR | ||||||
| Metformin | AMPK activation | Global cerebral ischemia | Rats | ↑ neuroprotection | [81] | |
| Metformin | ↓ oxidative stress | Ischemic stroke and diabetes | Humans | ↓ NIHSS | [82] | |
| Sinomenine | suppresses NLRP3 inflammasomes | MCAO | Mice | ↑ neuroprotection | [95] | |
| Sinomenine | CRYAB/STAT3 pathway | MCAO | Mice | ↓ cerebral infarction ↓ neuronal apoptosis ↑ neurological deficits | [96] | |
| Sinomenine | Nrf2 pathway | MCAO | Mice | ↓ inflammation | [97] | |
| Resveratrol | cAMP/AMPK/SIRT1 pathway | MCAO | Rats | ↑ neuroprotection | [103] | |
| Resveratrol | PI3K/AKT/mTOR pathway | MCAO | Rats | ↓ neurological damage ↓ infarct volume | [105] | |
| Resveratrol | Ischemic stroke | Humans | Regulates blood pressure, glycemia and lipid profile | [107] | ||
| Resveratrol | MMP-2, MMP-9 | Ischemic stroke | Humans | ↓ NIHSS | [108] | |
| Tat-NR2B9c | NI | Macaques | ↓ infarct volume ↑ NHPSS | [113] | ||
| Tat-NR2B9c | NI | Intracranial aneurysm | Humans | fewer ischemic infarcts | [114] | |
| Hypothermia | pAMPK inhibition | MCAO | Mice | ↓ infarct volume | [116] | |
| Hypothermia | NI | Ischemic stroke | Humans | ↑ incidence of pneumonia | [124] | |
| Hypothermia | NI | Ischemic stroke | Humans | No difference between groups | [125] | |
| Hypothermia | MMP-3, FABP, IL-8 | Ischemic stroke | Humans | ↑ MMP-3, FABP, IL-8 | [126] | |
| Hypothermia + RIPC | NLRP3, MDA, SOD | Ischemic stroke | Humans | ↓ oxidative stress ↓ inflammation ↓ NIHSS | [127] | |
| Secretome | ||||||
| BMSC-Exos | NLRP3 | MCAO | Rats | ↓ infarct volume ↑ behavioral/cognitive deficit | [145] | |
| HCB-SCs | suppresses NLRP3 inflammasomes | MCAO | Rats | ↓ neuronal apoptosis | [150] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pinoșanu, E.A.; Pîrșcoveanu, D.; Albu, C.V.; Burada, E.; Pîrvu, A.; Surugiu, R.; Sandu, R.E.; Serb, A.F. Rhoa/ROCK, mTOR and Secretome-Based Treatments for Ischemic Stroke: New Perspectives. Curr. Issues Mol. Biol. 2024, 46, 3484-3501. https://doi.org/10.3390/cimb46040219
Pinoșanu EA, Pîrșcoveanu D, Albu CV, Burada E, Pîrvu A, Surugiu R, Sandu RE, Serb AF. Rhoa/ROCK, mTOR and Secretome-Based Treatments for Ischemic Stroke: New Perspectives. Current Issues in Molecular Biology. 2024; 46(4):3484-3501. https://doi.org/10.3390/cimb46040219
Chicago/Turabian StylePinoșanu, Elena Anca, Denisa Pîrșcoveanu, Carmen Valeria Albu, Emilia Burada, Andrei Pîrvu, Roxana Surugiu, Raluca Elena Sandu, and Alina Florina Serb. 2024. "Rhoa/ROCK, mTOR and Secretome-Based Treatments for Ischemic Stroke: New Perspectives" Current Issues in Molecular Biology 46, no. 4: 3484-3501. https://doi.org/10.3390/cimb46040219