
CCN1-Mediated Signaling in Placental Villous Tissues after SARS-CoV-2 Infection in Term Pregnant Women: Implications for Dysregulated Angiogenesis

 and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Population
2.2. Immunoblotting
2.3. Immunofluorescence and Immunohistochemical Examination
2.4. Statistical Analysis
3. Results
3.1. Characteristics of Pregnant Women and Neonatal Outcome with and without Maternal SARS-CoV-2 Infection in Term Pregnancies
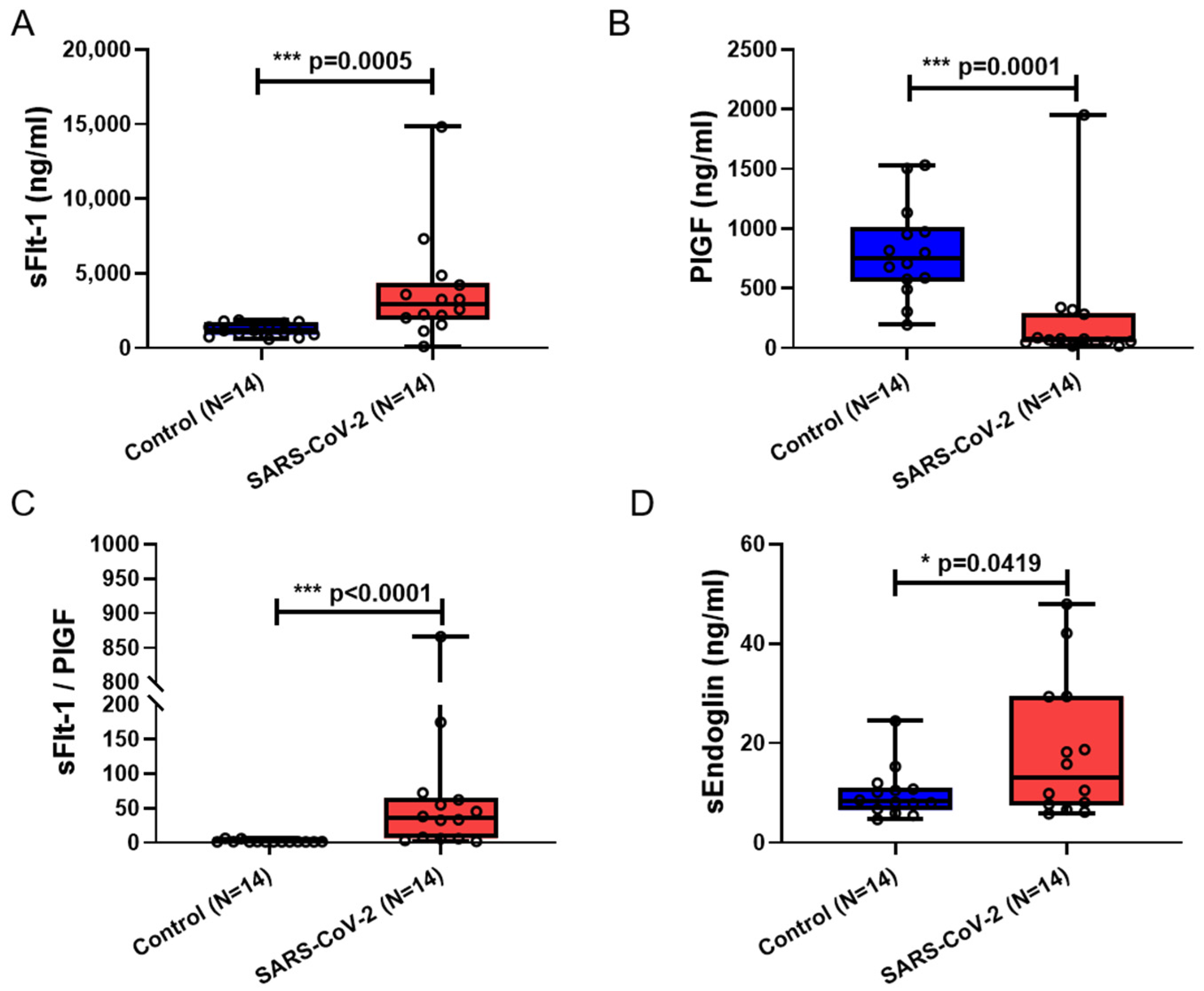
3.2. Serum sFlt-1, sFlt-1/PlGF, and sEng Were Significantly Elevated in Maternal Blood from Patients Infected with SARS-CoV-2
3.3. Decreased Vascular Remodeling and Increased ROS Level in Placental Villi from Patients Infected with SARS-CoV-2
3.4. CCN1 and pNF-κB Expression Was Increased in Placental Villi from SARS-CoV-2-Infected Pregnancies
3.5. Flt-1 and Endoglin (Eng) Expression Was Increased in Placental Villi from SARS-CoV-2-Infected Pregnancies
3.6. TGFβR1 and eNOS Expression Are Counterregulated in Placental Villi from SARS-CoV-2-Infected Pregnancies
3.7. Recombinant CCN1 Induced Flt-1 and Endoglin Protein Expression in SGHPL-5 Trophoblast Cells, along with the Release of sFlt-1 and sEng
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
References
- Hoen, B.; Schaub, B.; Funk, A.L.; Ardillon, V.; Boullard, M.; Cabie, A.; Callier, C.; Carles, G.; Cassadou, S.; Cesaire, R.; et al. Pregnancy Outcomes after ZIKV Infection in French Territories in the Americas. N. Engl. J. Med. 2018, 378, 985–994. [Google Scholar] [CrossRef] [PubMed]
- Leruez-Ville, M.; Magny, J.F.; Couderc, S.; Pichon, C.; Parodi, M.; Bussieres, L.; Guilleminot, T.; Ghout, I.; Ville, Y. Risk Factors for Congenital Cytomegalovirus Infection Following Primary and Nonprimary Maternal Infection: A Prospective Neonatal Screening Study Using Polymerase Chain Reaction in Saliva. Clin. Infect. Dis. 2017, 65, 398–404. [Google Scholar] [CrossRef] [PubMed]
- Megli, C.J.; Coyne, C.B. Infections at the maternal-fetal interface: An overview of pathogenesis and defence. Nat. Rev. Microbiol. 2022, 20, 67–82. [Google Scholar] [CrossRef] [PubMed]
- Lu, R.; Zhao, X.; Li, J.; Niu, P.; Yang, B.; Wu, H.; Wang, W.; Song, H.; Huang, B.; Zhu, N.; et al. Genomic characterisation and epidemiology of 2019 novel coronavirus: Implications for virus origins and receptor binding. Lancet 2020, 395, 565–574. [Google Scholar] [CrossRef] [PubMed]
- Fan, C.; Lei, D.; Fang, C.; Li, C.; Wang, M.; Liu, Y.; Bao, Y.; Sun, Y.; Huang, J.; Guo, Y.; et al. Perinatal Transmission of 2019 Coronavirus Disease-Associated Severe Acute Respiratory Syndrome Coronavirus 2: Should We Worry? Clin. Infect. Dis. 2021, 72, 862–864. [Google Scholar] [CrossRef] [PubMed]
- Juan, J.; Gil, M.M.; Rong, Z.; Zhang, Y.; Yang, H.; Poon, L.C. Effect of coronavirus disease 2019 (COVID-19) on maternal, perinatal and neonatal outcome: Systematic review. Ultrasound Obs. Gynecol. 2020, 56, 15–27. [Google Scholar] [CrossRef] [PubMed]
- Villar, J.; Ariff, S.; Gunier, R.B.; Thiruvengadam, R.; Rauch, S.; Kholin, A.; Roggero, P.; Prefumo, F.; do Vale, M.S.; Cardona-Perez, J.A.; et al. Maternal and Neonatal Morbidity and Mortality Among Pregnant Women With and Without COVID-19 Infection: The INTERCOVID Multinational Cohort Study. JAMA Pediatr. 2021, 175, 817–826. [Google Scholar] [CrossRef] [PubMed]
- Hsu, A.L.; Guan, M.; Johannesen, E.; Stephens, A.J.; Khaleel, N.; Kagan, N.; Tuhlei, B.C.; Wan, X.F. Placental SARS-CoV-2 in a pregnant woman with mild COVID-19 disease. J. Med. Virol. 2021, 93, 1038–1044. [Google Scholar] [CrossRef] [PubMed]
- Reagan-Steiner, S.; Bhatnagar, J.; Martines, R.B.; Milligan, N.S.; Gisondo, C.; Williams, F.B.; Lee, E.; Estetter, L.; Bullock, H.; Goldsmith, C.S.; et al. Detection of SARS-CoV-2 in Neonatal Autopsy Tissues and Placenta. Emerg. Infect. Dis. 2022, 28, 510–517. [Google Scholar] [CrossRef] [PubMed]
- McMahon, C.L.; Castro, J.; Silvas, J.; Muniz Perez, A.; Estrada, M.; Carrion, R., Jr.; Hsieh, J. Fetal brain vulnerability to SARS-CoV-2 infection. Brain. Behav. Immun. 2023, 112, 188–205. [Google Scholar] [CrossRef] [PubMed]
- Torge, D.; Bernardi, S.; Arcangeli, M.; Bianchi, S. Histopathological Features of SARS-CoV-2 in Extrapulmonary Organ Infection: A Systematic Review of Literature. Pathogens 2022, 11, 867. [Google Scholar] [CrossRef] [PubMed]
- Kubota, S.; Takigawa, M. CCN family proteins and angiogenesis: From embryo to adulthood. Angiogenesis 2007, 10, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Schober, J.M.; Lau, L.F.; Ugarova, T.P.; Lam, S.C. Identification of a novel integrin alphaMbeta2 binding site in CCN1 (CYR61), a matricellular protein expressed in healing wounds and atherosclerotic lesions. J. Biol. Chem. 2003, 278, 25808–25815. [Google Scholar] [CrossRef] [PubMed]
- Lobel, M.; Bauer, S.; Meisel, C.; Eisenreich, A.; Kudernatsch, R.; Tank, J.; Rauch, U.; Kuhl, U.; Schultheiss, H.P.; Volk, H.D.; et al. CCN1: A novel inflammation-regulated biphasic immune cell migration modulator. Cell Mol. Life Sci. 2012, 69, 3101–3113. [Google Scholar] [CrossRef] [PubMed]
- Imhof, B.A.; Jemelin, S.; Ballet, R.; Vesin, C.; Schapira, M.; Karaca, M.; Emre, Y. CCN1/CYR61-mediated meticulous patrolling by Ly6Clow monocytes fuels vascular inflammation. Proc. Natl. Acad. Sci. USA 2016, 113, E4847–E4856. [Google Scholar] [CrossRef]
- Sun, J.; Zhang, W.; Tan, Z.; Zheng, C.; Tang, Y.; Ke, X.; Zhang, Y.; Liu, Y.; Li, P.; Hu, Q.; et al. Zika virus promotes CCN1 expression via the CaMKIIalpha-CREB pathway in astrocytes. Virulence 2020, 11, 113–131. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.M.; Park, J.H.; Chung, S.K.; Kim, J.Y.; Hwang, H.Y.; Chung, K.C.; Jo, I.; Park, S.I.; Nam, J.H. Coxsackievirus B3 infection induces cyr61 activation via JNK to mediate cell death. J. Virol. 2004, 78, 13479–13488. [Google Scholar] [CrossRef] [PubMed]
- Kase, Y.; Okano, H. Expression of ACE2 and a viral virulence-regulating factor CCN family member 1 in human iPSC-derived neural cells: Implications for COVID-19-related CNS disorders. Inflamm. Regen. 2020, 40, 32. [Google Scholar] [CrossRef]
- Forsyth, C.B.; Zhang, L.; Bhushan, A.; Swanson, B.; Zhang, L.; Mamede, J.I.; Voigt, R.M.; Shaikh, M.; Engen, P.A.; Keshavarzian, A. The SARS-CoV-2 S1 Spike Protein Promotes MAPK and NF-kB Activation in Human Lung Cells and Inflammatory Cytokine Production in Human Lung and Intestinal Epithelial Cells. Microorganisms 2022, 10, 1996. [Google Scholar] [CrossRef] [PubMed]
- Jiang, R.; Tang, J.; Zhang, X.; He, Y.; Yu, Z.; Chen, S.; Xia, J.; Lin, J.; Ou, Q. CCN1 Promotes Inflammation by Inducing IL-6 Production via alpha6beta1/PI3K/Akt/NF-kappaB Pathway in Autoimmune Hepatitis. Front. Immunol. 2022, 13, 810671. [Google Scholar] [CrossRef] [PubMed]
- Bai, T.; Chen, C.C.; Lau, L.F. Matricellular protein CCN1 activates a proinflammatory genetic program in murine macrophages. J. Immunol. 2010, 184, 3223–3232. [Google Scholar] [CrossRef] [PubMed]
- Umapathy, A.; Chamley, L.W.; James, J.L. Reconciling the distinct roles of angiogenic/anti-angiogenic factors in the placenta and maternal circulation of normal and pathological pregnancies. Angiogenesis 2020, 23, 105–117. [Google Scholar] [CrossRef] [PubMed]
- Maynard, S.E.; Min, J.Y.; Merchan, J.; Lim, K.H.; Li, J.; Mondal, S.; Libermann, T.A.; Morgan, J.P.; Sellke, F.W.; Stillman, I.E.; et al. Excess placental soluble fms-like tyrosine kinase 1 (sFlt1) may contribute to endothelial dysfunction, hypertension, and proteinuria in preeclampsia. J. Clin. Investig. 2003, 111, 649–658. [Google Scholar] [CrossRef] [PubMed]
- Venkatesha, S.; Toporsian, M.; Lam, C.; Hanai, J.; Mammoto, T.; Kim, Y.M.; Bdolah, Y.; Lim, K.H.; Yuan, H.T.; Libermann, T.A.; et al. Soluble endoglin contributes to the pathogenesis of preeclampsia. Nat. Med. 2006, 12, 642–649. [Google Scholar] [CrossRef] [PubMed]
- Lecarpentier, E.; Tsatsaris, V. Angiogenic balance (sFlt-1/PlGF) and preeclampsia. Ann. Endocrinol. 2016, 77, 97–100. [Google Scholar] [CrossRef] [PubMed]
- Perry, H.; Binder, J.; Kalafat, E.; Jones, S.; Thilaganathan, B.; Khalil, A. Angiogenic Marker Prognostic Models in Pregnant Women With Hypertension. Hypertension 2020, 75, 755–761. [Google Scholar] [CrossRef] [PubMed]
- Hosier, H.; Farhadian, S.F.; Morotti, R.A.; Deshmukh, U.; Lu-Culligan, A.; Campbell, K.H.; Yasumoto, Y.; Vogels, C.B.; Casanovas-Massana, A.; Vijayakumar, P.; et al. SARS-CoV-2 infection of the placenta. J. Clin. Investig. 2020, 130, 4947–4953. [Google Scholar] [CrossRef] [PubMed]
- Gychka, S.G.; Brelidze, T.I.; Kuchyn, I.L.; Savchuk, T.V.; Nikolaienko, S.I.; Zhezhera, V.M.; Chermak, I.I.; Suzuki, Y.J. Placental vascular remodeling in pregnant women with COVID-19. PLoS ONE 2022, 17, e0268591. [Google Scholar] [CrossRef] [PubMed]
- Schwartz, D.A.; Baldewijns, M.; Benachi, A.; Bugatti, M.; Collins, R.R.J.; De Luca, D.; Facchetti, F.; Linn, R.L.; Marcelis, L.; Morotti, D.; et al. Chronic Histiocytic Intervillositis With Trophoblast Necrosis Is a Risk Factor Associated With Placental Infection From Coronavirus Disease 2019 (COVID-19) and Intrauterine Maternal-Fetal Severe Acute Respiratory Syndrome Coronavirus 2 (SARS-CoV-2) Transmission in Live-Born and Stillborn Infants. Arch. Pathol. Lab. Med. 2021, 145, 517–528. [Google Scholar] [PubMed]
- Watkins, J.C.; Torous, V.F.; Roberts, D.J. Defining Severe Acute Respiratory Syndrome Coronavirus 2 (SARS-CoV-2) Placentitis. Arch. Pathol. Lab. Med. 2021, 145, 1341–1349. [Google Scholar] [CrossRef] [PubMed]
- Shanes, E.D.; Mithal, L.B.; Otero, S.; Azad, H.A.; Miller, E.S.; Goldstein, J.A. Placental Pathology in COVID-19. Am. J. Clin. Pathol. 2020, 154, 23–32. [Google Scholar] [CrossRef] [PubMed]
- Arts, N.; Schiffer, V.; Severens-Rijvers, C.; Bons, J.; Spaanderman, M.; Al-Nasiry, S. Cumulative effect of maternal vascular malperfusion types in the placenta on adverse pregnancy outcomes. Placenta 2022, 129, 43–50. [Google Scholar] [CrossRef] [PubMed]
- Carvajal, J.; Casanello, P.; Toso, A.; Farias, M.; Carrasco-Negue, K.; Araujo, K.; Valero, P.; Fuenzalida, J.; Solari, C.; Sobrevia, L. Functional consequences of SARS-CoV-2 infection in pregnant women, fetoplacental unit, and neonate. Biochim. Biophys. Acta Mol. Basis Dis. 2023, 1869, 166582. [Google Scholar] [CrossRef] [PubMed]
- Therapeutics and COVID-19: Living Guideline; World Health Organization: Geneva, Switzerland, 2022.
- Gellhaus, A.; Schmidt, M.; Dunk, C.; Lye, S.J.; Kimmig, R.; Winterhager, E. Decreased expression of the angiogenic regulators CYR61 (CCN1) and NOV (CCN3) in human placenta is associated with pre-eclampsia. Mol. Hum. Reprod. 2006, 12, 389–399. [Google Scholar] [CrossRef] [PubMed]
- Duan, L.; Reisch, B.; Iannaccone, A.; Hadrovic, E.; Wu, Y.; Vogtmann, R.; Winterhager, E.; Kimmig, R.; Koninger, A.; Mach, P.; et al. Abnormal expression of the costimulatory molecule B7-H4 in placental chorionic villous and decidual basalis tissues of patients with preeclampsia and HELLP syndrome. Am. J. Reprod. Immunol. 2021, 86, e13430. [Google Scholar] [CrossRef] [PubMed]
- Goud, P.T.; Bai, D.; Abu-Soud, H.M. A Multiple-Hit Hypothesis Involving Reactive Oxygen Species and Myeloperoxidase Explains Clinical Deterioration and Fatality in COVID-19. Int. J. Biol. Sci. 2021, 17, 62–72. [Google Scholar] [CrossRef] [PubMed]
- Khomich, O.A.; Kochetkov, S.N.; Bartosch, B.; Ivanov, A.V. Redox Biology of Respiratory Viral Infections. Viruses 2018, 10, 392. [Google Scholar] [CrossRef] [PubMed]
- Ryter, S.W.; Alam, J.; Choi, A.M. Heme oxygenase-1/carbon monoxide: From basic science to therapeutic applications. Physiol. Rev. 2006, 86, 583–650. [Google Scholar] [CrossRef] [PubMed]
- Seiwert, N.; Wecklein, S.; Demuth, P.; Hasselwander, S.; Kemper, T.A.; Schwerdtle, T.; Brunner, T.; Fahrer, J. Heme oxygenase 1 protects human colonocytes against ROS formation, oxidative DNA damage and cytotoxicity induced by heme iron, but not inorganic iron. Cell Death Dis. 2020, 11, 787. [Google Scholar] [CrossRef] [PubMed]
- Duan, L.; Reisch, B.; Mach, P.; Kimmig, R.; Gellhaus, A.; Iannaccone, A. The immunological role of B7-H4 in pregnant women with SARS-CoV-2 infection. Am. J. Reprod. Immunol. 2022, 88, e13626. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.Y.; Su, C.M.; Huang, Y.L.; Tsai, C.H.; Fuh, L.J.; Tang, C.H. CCN1 induces oncostatin M production in osteoblasts via integrin-dependent signal pathways. PLoS ONE 2014, 9, e106632. [Google Scholar] [CrossRef] [PubMed]
- Vicen, M.; Igreja Sa, I.C.; Tripska, K.; Vitverova, B.; Najmanova, I.; Eissazadeh, S.; Micuda, S.; Nachtigal, P. Membrane and soluble endoglin role in cardiovascular and metabolic disorders related to metabolic syndrome. Cell Mol. Life Sci. 2021, 78, 2405–2418. [Google Scholar] [CrossRef] [PubMed]
- ElMonier, A.A.; El-Boghdady, N.A.; Abdelaziz, M.A.; Shaheen, A.A. Association between endoglin/transforming growth factor beta receptors 1, 2 gene polymorphisms and the level of soluble endoglin with preeclampsia in Egyptian women. Arch. Biochem. Biophys. 2019, 662, 7–14. [Google Scholar] [CrossRef] [PubMed]
- Varejckova, M.; Gallardo-Vara, E.; Vicen, M.; Vitverova, B.; Fikrova, P.; Dolezelova, E.; Rathouska, J.; Prasnicka, A.; Blazickova, K.; Micuda, S.; et al. Soluble endoglin modulates the pro-inflammatory mediators NF-kappaB and IL-6 in cultured human endothelial cells. Life Sci. 2017, 175, 52–60. [Google Scholar] [CrossRef] [PubMed]
- Kipkeew, F.; Kirsch, M.; Klein, D.; Wuelling, M.; Winterhager, E.; Gellhaus, A. CCN1 (CYR61) and CCN3 (NOV) signaling drives human trophoblast cells into senescence and stimulates migration properties. Cell Adh. Migr. 2016, 10, 163–178. [Google Scholar] [CrossRef] [PubMed]
- Giardini, V.; Carrer, A.; Casati, M.; Contro, E.; Vergani, P.; Gambacorti-Passerini, C. Increased sFLT-1/PlGF ratio in COVID-19: A novel link to angiotensin II-mediated endothelial dysfunction. Am. J. Hematol. 2020, 95, E188–E191. [Google Scholar] [CrossRef] [PubMed]
- Negro, A.; Fama, A.; Penna, D.; Belloni, L.; Zerbini, A.; Giuri, P.G. SFLT-1 levels in COVID-19 patients: Association with outcome and thrombosis. Am. J. Hematol. 2021, 96, E41–E43. [Google Scholar] [CrossRef] [PubMed]
- Shibuya, M. Vascular Endothelial Growth Factor (VEGF) and Its Receptor (VEGFR) Signaling in Angiogenesis: A Crucial Target for Anti- and Pro-Angiogenic Therapies. Genes. Cancer 2011, 2, 1097–1105. [Google Scholar] [CrossRef] [PubMed]
- Margioula-Siarkou, G.; Margioula-Siarkou, C.; Petousis, S.; Margaritis, K.; Vavoulidis, E.; Gullo, G.; Alexandratou, M.; Dinas, K.; Sotiriadis, A.; Mavromatidis, G. The role of endoglin and its soluble form in pathogenesis of preeclampsia. Mol. Cell Biochem. 2022, 477, 479–491. [Google Scholar] [CrossRef] [PubMed]
- Krause, B.J.; Hanson, M.A.; Casanello, P. Role of nitric oxide in placental vascular development and function. Placenta 2011, 32, 797–805. [Google Scholar] [CrossRef] [PubMed]
- Kosinska-Kaczynska, K.; Malicka, E.; Szymusik, I.; Dera, N.; Pruc, M.; Feduniw, S.; Rafique, Z.; Szarpak, L. The sFlt-1/PlGF Ratio in Pregnant Patients Affected by COVID-19. J. Clin. Med. 2023, 12, 1059. [Google Scholar] [CrossRef] [PubMed]
- Facchetti, F.; Bugatti, M.; Drera, E.; Tripodo, C.; Sartori, E.; Cancila, V.; Papaccio, M.; Castellani, R.; Casola, S.; Boniotti, M.B.; et al. SARS-CoV-2 vertical transmission with adverse effects on the newborn revealed through integrated immunohistochemical, electron microscopy and molecular analyses of Placenta. EBioMedicine 2020, 59, 102951. [Google Scholar] [CrossRef] [PubMed]
- Winterhager, E.; Gellhaus, A. The role of the CCN family of proteins in female reproduction. Cell Mol. Life Sci. 2014, 71, 2299–2311. [Google Scholar] [CrossRef] [PubMed]
- Li, X.C.; Jin, F.; Wang, B.Y.; Yin, X.J.; Hong, W.; Tian, F.J. The m6A demethylase ALKBH5 controls trophoblast invasion at the maternal-fetal interface by regulating the stability of CYR61 mRNA. Theranostics 2019, 9, 3853–3865. [Google Scholar] [CrossRef] [PubMed]
- Shook, L.L.; Sullivan, E.L.; Lo, J.O.; Perlis, R.H.; Edlow, A.G. COVID-19 in pregnancy: Implications for fetal brain development. Trends Mol. Med. 2022, 28, 319–330. [Google Scholar] [CrossRef] [PubMed]
- Emre, Y.; Imhof, B.A. Matricellular protein CCN1/CYR61: A new player in inflammation and leukocyte trafficking. Semin. Immunopathol. 2014, 36, 253–259. [Google Scholar] [CrossRef] [PubMed]
- Dri, E.; Lampas, E.; Lazaros, G.; Lazarou, E.; Theofilis, P.; Tsioufis, C.; Tousoulis, D. Inflammatory Mediators of Endothelial Dysfunction. Life 2023, 13, 1420. [Google Scholar] [CrossRef] [PubMed]
- Iannaccone, A.; Gellhaus, A.; Reisch, B.; Dzietko, M.; Schmidt, B.; Mavarani, L.; Kraft, K.; Andresen, K.; Kimmig, R.; Pecks, U.; et al. The Importance of Vaccination, Variants and Time Point of SARS-CoV-2 Infection in Pregnancy for Stillbirth and Preterm Birth Risk: An Analysis of the CRONOS Register Study. J. Clin. Med. 2024, 13, 1522. [Google Scholar] [CrossRef]
- Kaiser, S.; Koch, Y.; Kuhnel, E.; Sharma, N.; Gellhaus, A.; Kuckenberg, P.; Schorle, H.; Winterhager, E. Reduced Gene Dosage of Tfap2c Impairs Trophoblast Lineage Differentiation and Alters Maternal Blood Spaces in the Mouse Placenta. Biol. Reprod. 2015, 93, 31. [Google Scholar] [CrossRef] [PubMed]
- Vogtmann, R.; Bao, M.; Dewan, M.V.; Riedel, A.; Kimmig, R.; Felderhoff-Muser, U.; Bendix, I.; Plosch, T.; Gellhaus, A. Growth-Restricted Fetuses and Offspring Reveal Adverse Sex-Specific Metabolic Responses in Preeclamptic Mice Expressing Human sFLT1. Int. J. Mol. Sci. 2023, 24, 6885. [Google Scholar] [CrossRef] [PubMed]
- Iannaccone, A.; Reisch, B.; Kimmig, R.; Schmidt, B.; Mavarani, L.; Darkwah Oppong, M.; Tyczynski, B.; Dzietko, M.; Jahn, M.; Gellhaus, A.; et al. Therapeutic Plasma Exchange in Early-Onset Preeclampsia: A 7-Year Monocentric Experience. J. Clin. Med. 2023, 12, 4289. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variable | Control (n = 23) | SARS-CoV-2 (n = 24) | p-Value |
|---|---|---|---|
| Maternal age at delivery, years, median (IQR) | 34.00 (30.00–37.00) | 32.50 (29.25–36.50) | 0.4257 |
| Gestational age at delivery, weeks, mean (min and max) | 38 + 6 (36 + 2–41 + 0) | 39 + 0 (36 + 5–41 + 1) | 0.8336 |
| Pregnancy BMI before birth, median (IQR) | 31.00 (27.00–35.00) | 28.50 (26.25–32.75) | 0.3159 |
| Cesarean section, no. (%) | 14 (60.87) | 11 (45.83) | - |
| Leukocyte, 103/mm3, mean ± SD | 10.98 ± 2.96 | 9.07 ± 3.44 | 0.0363 |
| CRP, mg/dL, mean ± SD | 0.39 ± 0.42 | 2.54 ± 3.61 | 0.0197 |
| Asymptomatic infection, no. (%) | - | 15 (62.50) | - |
| Mild infection, no. (%) | - | 9 (37.50) | - |
| Birth weight, g, mean ± SD | 3387 ± 392.70 | 3570 ± 529.90 | 0.3853 |
| 5-min Apgar score, mean (min and max) | 9.87 (9–10) | 9.58 (8–10) | 0.0686 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ma, Y.; Duan, L.; Reisch, B.; Kimmig, R.; Iannaccone, A.; Gellhaus, A. CCN1-Mediated Signaling in Placental Villous Tissues after SARS-CoV-2 Infection in Term Pregnant Women: Implications for Dysregulated Angiogenesis. Curr. Issues Mol. Biol. 2024, 46, 3533-3550. https://doi.org/10.3390/cimb46040221
Ma Y, Duan L, Reisch B, Kimmig R, Iannaccone A, Gellhaus A. CCN1-Mediated Signaling in Placental Villous Tissues after SARS-CoV-2 Infection in Term Pregnant Women: Implications for Dysregulated Angiogenesis. Current Issues in Molecular Biology. 2024; 46(4):3533-3550. https://doi.org/10.3390/cimb46040221
Chicago/Turabian StyleMa, Yuyang, Liyan Duan, Beatrix Reisch, Rainer Kimmig, Antonella Iannaccone, and Alexandra Gellhaus. 2024. "CCN1-Mediated Signaling in Placental Villous Tissues after SARS-CoV-2 Infection in Term Pregnant Women: Implications for Dysregulated Angiogenesis" Current Issues in Molecular Biology 46, no. 4: 3533-3550. https://doi.org/10.3390/cimb46040221