Effect of Pressure on Thermal Stability of G-Quadruplex DNA and Double-Stranded DNA Structures

Abstract
:1. Introduction
2. Pressure Effect on Canonical Duplex of Nucleic Acids
2.1. Melting Analysis under High Pressure by Temperature Change

| DNA | Salt Concentration | ∆Tm/∆P (10−2 K MPa−1) | ∆Vtr (cm3 mol−1) | Ref. |
|---|---|---|---|---|
| Calf thymus | [NaCl] = 30 mM | 4.49 | −4.5 | [16] |
| [KCl] = 140 mM | 2.34 | −2.7 | [17] | |
| [KCl] = 5 mM | 0.46 | −0.51 | [18] | |
| [KCl] = 20 mM | 1.4 | −1.58 | ||
| [KCl] = 50 mM | 2.0 | −2.27 | ||
| [KCl] = 200 mM | 2.9 | −3.32 | ||
| [KCl] = 500 mM | 3.5 | −4.02 | ||
| C. perfringens | [NaCl] = 10 mM | 0.54 | [19] | |
| [NaCl] = 50 mM | 2.0 | |||
| [NaCl] = 120 mM | 2.6 | |||
| [NaCl] = 360 mM | 3.8 | |||
| [NaCl] = 1.08 M | 4.1 | |||
| [NaCl] = 3.6 M | 4.6 | |||
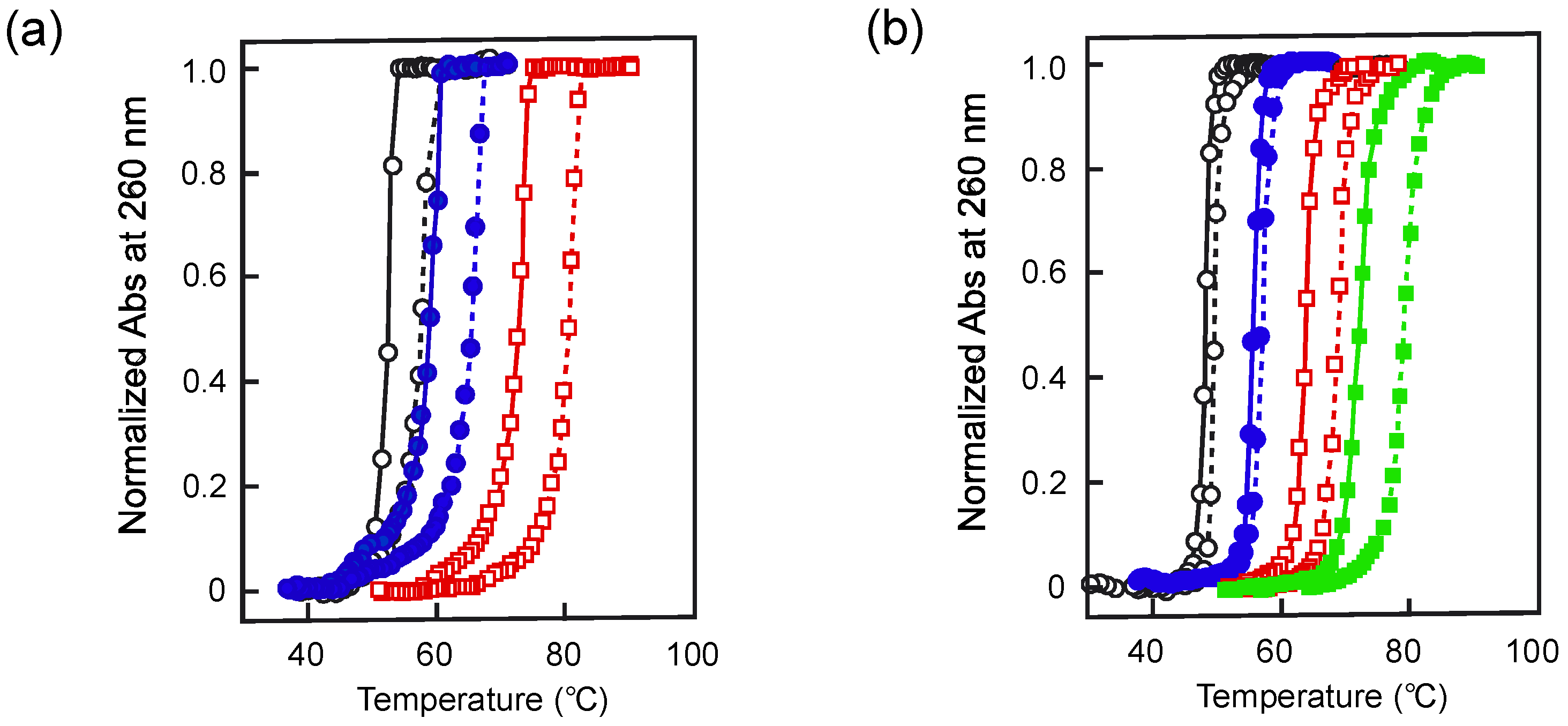
| poly[d(A-T)] | [NaCl] = 20 mM | 0.36 | −0.36 | [23] |
| [NaCl] = 50 mM | 0.93 | −0.90 | ||
| [NaCl] = 200 mM | 2.26 | −2.14 | ||
| [NaCl] = 1.0 M | 3.86 | −3.57 | ||
| poly(dA) ·poly(dT) | [NaCl] = 20 mM | 2.49 | −2.60 | [23] |
| [NaCl] = 50 mM | 3.15 | −3.44 | ||
| [NaCl] = 200 mM | 3.86 | −4.59 | ||
| poly[d(G-C)] | [NaCl] = 52 mM | 4.51 | −4.80 | [24] |
| [NaCl] = 107 mM | 4.79 | −5.16 | ||
| [NaCl] = 300 mM | 5.01 | −5.50 | ||
| [NaCl] = 1.0 M | 6.41 | −6.03 | ||
| poly(rA) ·poly(rU) | [K+] = 50 mM | −1.07 | 0.96 | [25] |
| poly[d(I-C)] | [NaCl] = 75 mM | 0.28 | −0.26 | [26] |
| [NaCl] = 270 mM | 1.36 | −1.25 | ||
| [NaCl] = 1.0 M | 2.64 | −2.39 |

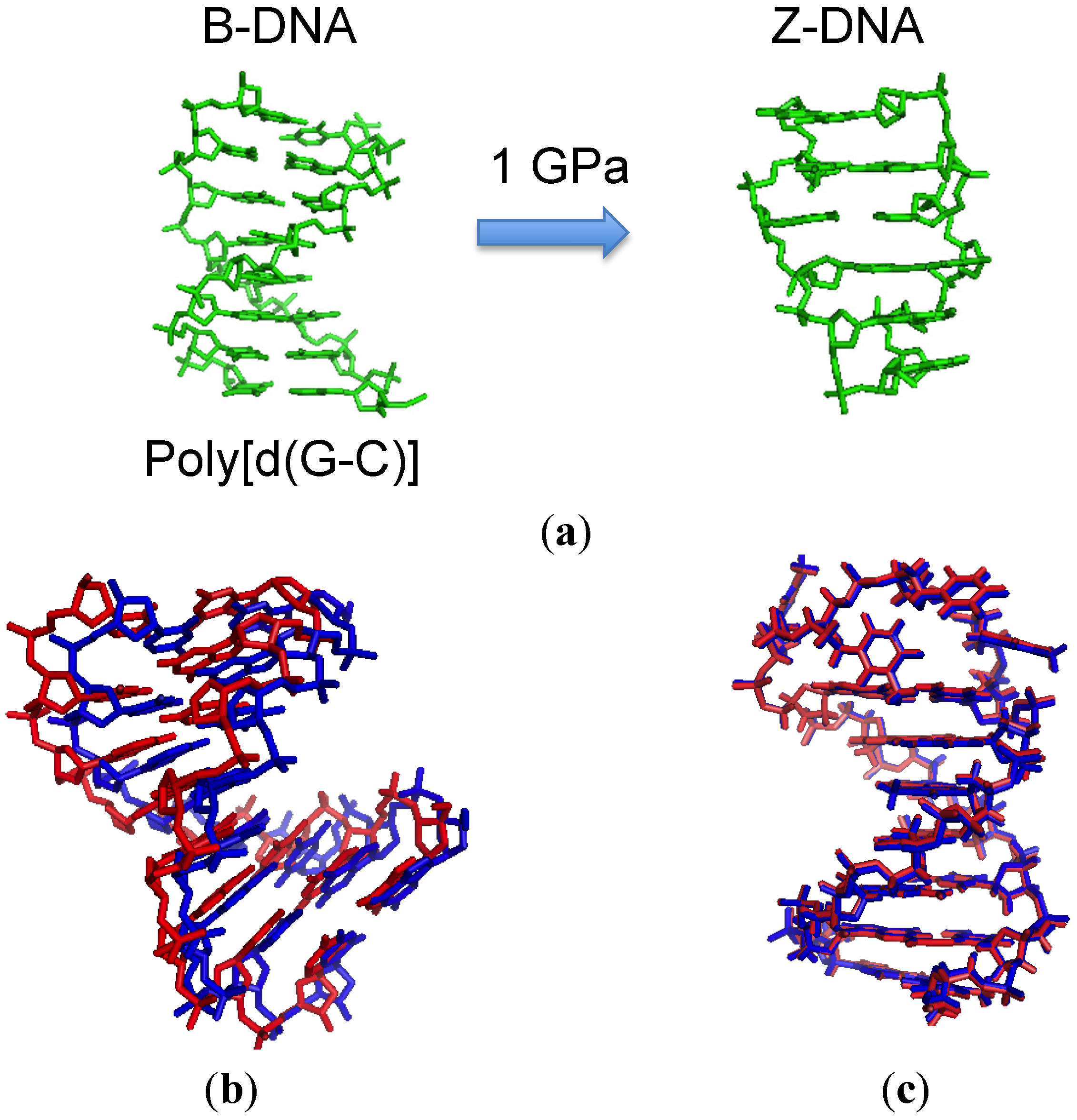
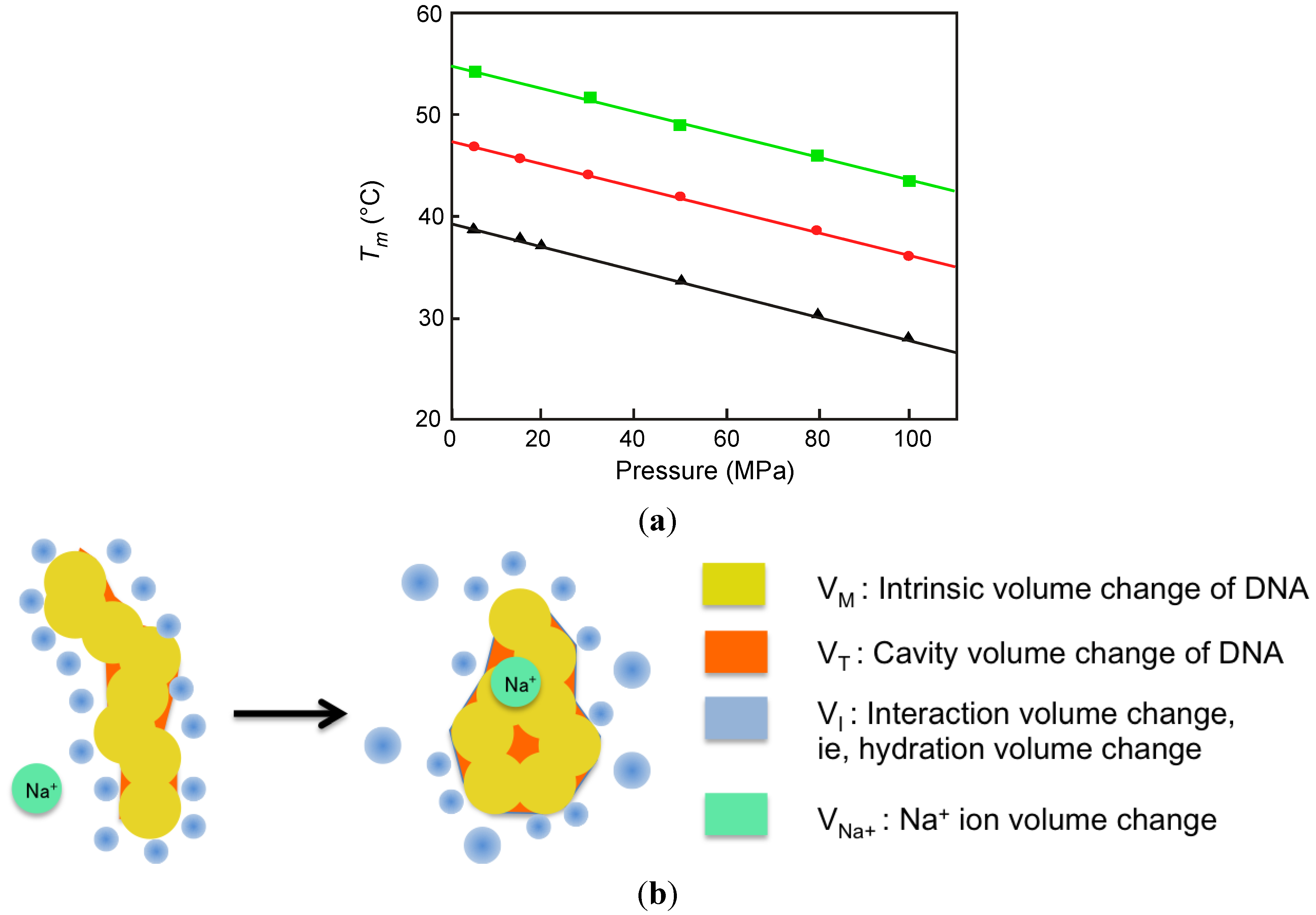
2.2. Effect of High Pressure on the Conformation of a Duplex

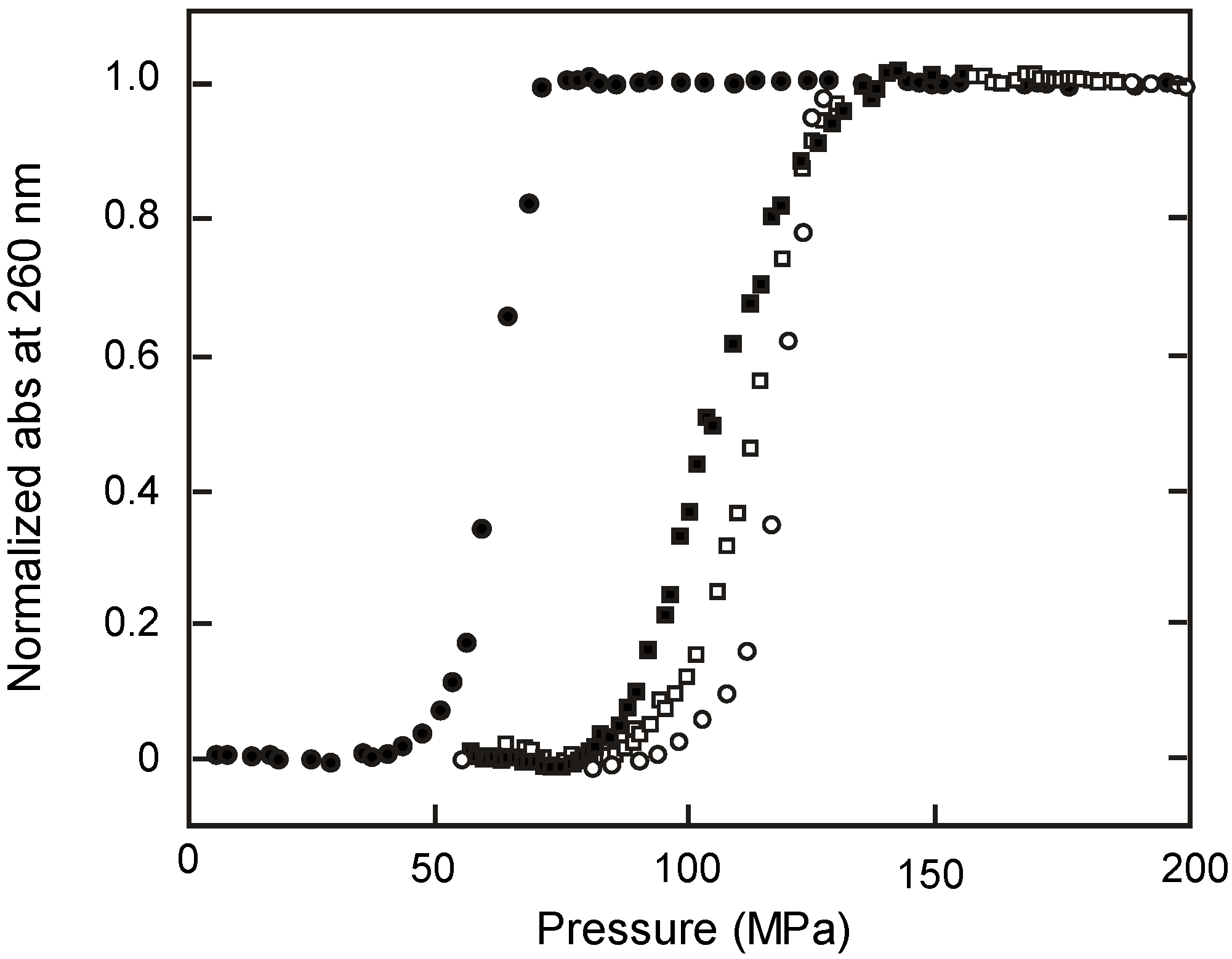
2.3. Melting of Duplex Induced by Pressure

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
2.4. Kinetic Analyses

| Fraction of GC | ∆V‡1 (cm3 mol−1) | ∆V‡−1 (cm3·mol−1) | ∆Vtr kinetic (cm3 mol−1) a | ∆Vtr (cm3 mol−1) b |
|---|---|---|---|---|
| 0.5 | −6.7 | 1.6 | −8.3 | −5.8 |
| 0.32 | −8.0 | 0.40 | −8.4 | −8.0 |
| 0.23 | −13 | 15 | −28 | −13 |
| 0.14 | −20 | 17 | −37 | −20 |
2.5. Effect of Pressure on the Interactions between DNA and Protein
3. Non-Canonical Structures of Nucleic Acids under High Pressure
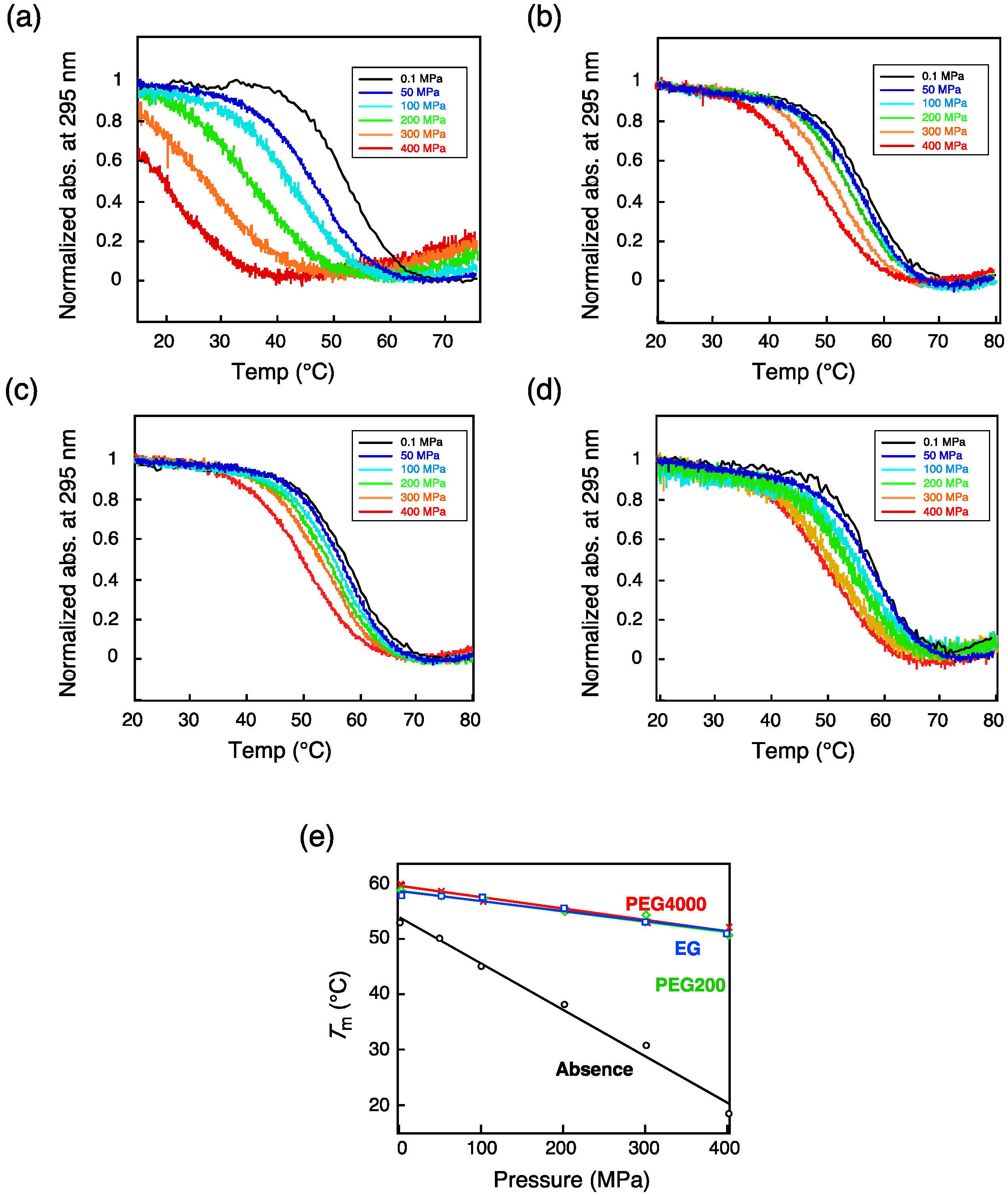
3.1. G-Quadruplex

| DNA | Salt or Cosolute | ∆Vtr (cm3 mol−1) |
|---|---|---|
| H-telo a | [NaCl] = 20 mM | 68 ± 2 |
| [NaCl] = 50 mM | 60 ± 2 | |
| [NaCl] = 100 mM | 56 ± 2 | |
| TBA b | (Absence) | 54.6 ± 4.2 |
| 40 wt% Ethylene glycol | 12.5 ± 0.8 | |
| 40 wt% PEG 200 | 12.9 ± 0.9 | |
| 40 wt% PEG 4000 | 13.1 ± 1.0 |

3.2. Triple Helix
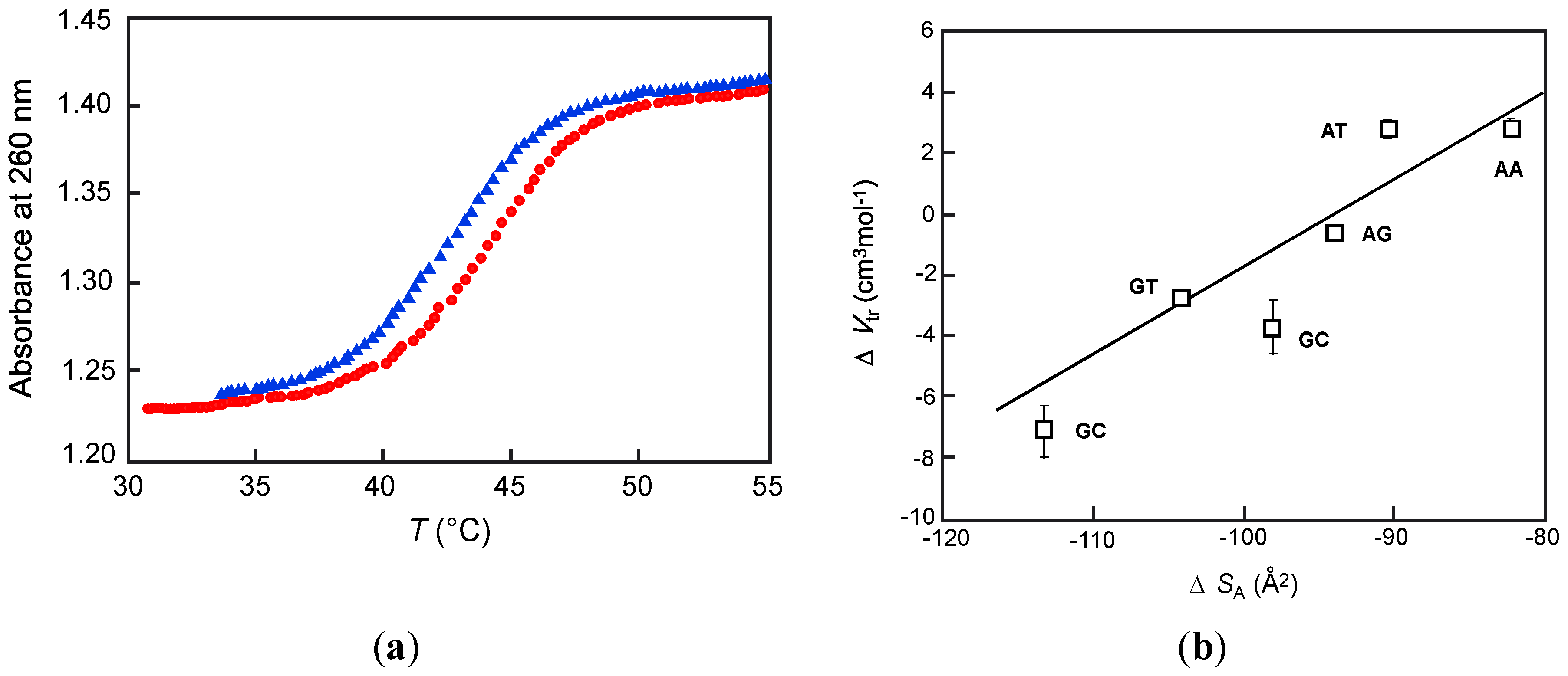
3.3. Hairpin DNA
| Loop sequence | |||||||
|---|---|---|---|---|---|---|---|
| TA2T | TG2T | TC2T | |||||
| Nucleation stack | Na+ (mM) | Tm (°C) | ∆Vtr (cm3 mol−1) a | Tm (°C) | ∆Vtr (cm3 mol−1) a | Tm (°C) | ∆Vtr (cm3 mol−1) a |
| AT/AT | 10 | 42.1 | 0.44 | 42.8 | 1.41 | 44.9 | −1.81 |
| 20 | 43.2 | −0.18 | 44.0 | 0.25 | 46.1 | −2.27 | |
| 50 | 44.6 | −0.83 | 45.5 | 1.55 | 48.7 | −3.05 | |
| 100 | 46.1 | −1.46 | 46.8 | −2.89 | 51.3 | −3.76 | |
| AA/TT | 10 | 40.2 | 1.96 | 37.9 | 2.35 | 44.0 | −0.78 |
| 20 | 41.5 | 1.15 | 41.1 | 0.86 | 45.4 | −1.18 | |
| 50 | 43.3 | 0.19 | 43.5 | −0.85 | 47.9 | −1.75 | |
| 100 | 44.7 | −0.74 | 45.1 | −2.14 | 49.9 | −2.35 | |
4. Summary and Perspectives
Acknowledgments
Conflicts of Interest
References and Notes
- Meersman, F.; Dobson, C.M.; Heremans, K. Protein unfolding, amyloid fibril formation and configurational energy landscapes under high pressure conditions. Chem. Soc. Rev. 2006, 35, 908–917. [Google Scholar]
- Silva, J.L.; Foguel, D.; Royer, C.A. Pressure provides new insights into protein folding, dynamics and structure. Trends Biochem. Sci. 2001, 26, 612–618. [Google Scholar] [CrossRef]
- Heremans, K.; Smeller, L. Protein structure and dynamics at high pressure. Biochim. Biophys. Acta 1998, 1386, 353–370. [Google Scholar]
- Akasaka, K. Probing conformational fluctuation of proteins by pressure perturbation. Chem. Rev. 2006, 106, 1814–1835. [Google Scholar]
- Hawley, S. Reversible pressure-temperature denaturation of chymotrypsinogen. Biochemistry 1971, 10, 2436–2442. [Google Scholar] [CrossRef]
- Frye, K.J.; Royer, C.A. Probing the contribution of internal cavities to the volume change of protein unfolding under pressure. Protein Sci. 1998, 7, 2217–2222. [Google Scholar]
- Paliwal, A.; Asthagiri, D.; Bossev, D.P.; Paulaitis, M.E. Pressure denaturation of staphylococcal nuclease studied by neutron small-angle scattering and molecular simulation. Biophys. J. 2004, 87, 3479–3492. [Google Scholar]
- Day, R.; García, A.E. Water penetration in the low and high pressure native states of ubiquitin. Proteins Struct. Funct. Bioinform. 2008, 70, 1175–1184. [Google Scholar]
- Imai, T.; Sugita, Y. Dynamic correlation between pressure-induced protein structural transition and water penetration. J. Phys. Chem. B 2010, 114, 2281–2286. [Google Scholar] [CrossRef]
- Fabian, H.; Naumann, D. Protein Folding and Misfolding: Shining Light by Infrared Spectroscopy; Springer: Heidelberg, Germany, 2012. [Google Scholar]
- Bridgman, P.W. The coagulation of albumin by pressure. J. Biol. Chem. 1914, 19, 511–512. [Google Scholar]
- Heden, C.; Lindahl, T.; Toplin, I. The stability of deoxyribonucleic acid solutions under high pressure. Acta Chem. Scand. 1964, 18, 1150–1158. [Google Scholar]
- Palumbo, S.L.; Ebbinghaus, S.W.; Hurley, L.H. Formation of a unique end-to-end stacked pair of G-quadruplexes in the hTERT core promoter with implications for inhibition of telomerase by G-quadruplex-interactive ligands. J. Am. Chem. Soc. 2009, 131, 10878–10891. [Google Scholar]
- Endoh, T.; Kawasaki, Y.; Sugimoto, N. Suppression of gene expression by G-quadruplexes in open reading frames depends on G-quadruplex stability. Angew. Chem. Int. Ed. 2013, 125, 5632–5636. [Google Scholar]
- Throughout this review, the volumetric change ∆V (= ∆Vtr) means the value for formation of the folded conformation and the unit of pressure is MPa. Some of these values in this review were recalculated from the original values in the references.
- Weida, B.; Gill, S. Pressure effect on deoxyribonucleic acid transition. Biochim. Biophys. Acta 1966, 112, 179–181. [Google Scholar]
- Gunter, T.; Gunter, K. Pressure dependence of the helix-coil transition temperature for polynucleic acid helices. Biopolymers 1972, 11, 667–678. [Google Scholar] [CrossRef]
- Nordmeier, E. Effects of pressure on the helix-coil transition of calf thymus DNA. J. Phys. Chem. 1992, 96, 1494–1501. [Google Scholar] [CrossRef]
- Hawley, S.; Macleod, R. Pressure-Temperature stability of DNA in neutral salt solutions. Biopolymers 1974, 13, 1417–1426. [Google Scholar]
- Panick, G.; Vidugiris, G.J.; Malessa, R.; Rapp, G.; Winter, R.; Royer, C.A. Exploring the temperature-pressure phase diagram of staphylococcal nuclease. Biochemistry 1999, 38, 4157–4164. [Google Scholar]
- Panick, G.; Malessa, R.; Winter, R.; Rapp, G.; Frye, K.J.; Royer, C.A. Structural characterization of the pressure-denatured state and unfolding/refolding kinetics of staphylococcal nuclease by synchrotron small-angle X-ray scattering and Fourier-transform infrared spectroscopy. J. Mol. Biol. 1998, 275, 389–402. [Google Scholar] [CrossRef]
- Panick, G.; Winter, R. Pressure-Induced unfolding/refolding of ribonuclease A: Static and kinetic Fourier transform infrared spectroscopy study. Biochemistry 2000, 39, 1862–1869. [Google Scholar] [CrossRef]
- Wu, J.Q.; Macgregor, R.B., Jr. Pressure dependence of the melting temperature of dA-dT polymers. Biochemistry 1993, 32, 12531–12537. [Google Scholar]
- Wu, J.Q.; Macgregor, R.B. Pressure dependence of the helix-coil transition temperature of poly [d (G-C)]. Biopolymers 1995, 35, 369–376. [Google Scholar]
- Hughes, F.; Steiner, R. Effects of pressure on the helix–coil transitions of the poly A–poly U system. Biopolymers 1966, 4, 1081–1090. [Google Scholar] [CrossRef]
- Markley, J.L.; Northrop, D.B. High Pressure Effects in Biophysics and Enzymology; Oxford University Press: New York, USA, 1996; pp. 274–207. [Google Scholar]
- Chalikian, T.V.; Völker, J.; Srinivasan, A.R.; Olson, W.K.; Breslauer, K.J. The hydration of nucleic acid duplexes as assessed by a combination of volumetric and structural techniques. Biopolymers 1999, 50, 459–471. [Google Scholar]
- Krzyźaniak, A.; Salański, P.; Jurczak, J.; Barciszewski, J. B-Z DNA reversible conformation changes effected by high pressure. FEBS Lett. 1991, 279, 1–4. [Google Scholar] [CrossRef]
- Benson, S.W.; Siebert, E.D. A simple two-structure model for liquid water. J. Am. Chem. Soc. 1992, 114, 4269–4276. [Google Scholar] [CrossRef]
- Kitchen, D.B.; Reed, L.H.; Levy, R.M. Molecular dynamics simulation of solvated protein at high pressure. Biochemistry 1992, 31, 10083–10093. [Google Scholar] [CrossRef]
- Leberman, R.; Soper, A. Effect of high salt concentrations on water structure. Nature 1995, 378, 364–366. [Google Scholar] [CrossRef]
- Krzyżaniak, A.; Barciszewski, J.; Fürste, J.P.; Bald, R.; Erdmann, V.A.; Salański, P.; Jurczak, J. AZ-RNA conformational changes effected by high pressure. Int. J. Biol. Macromol. 1994, 16, 159–162. [Google Scholar]
- Girard, E.; Prange, T.; Dhaussy, A.-C.; Migianu-Griffoni, E.; Lecouvey, M.; Chervin, J.-C.; Mezouar, M.; Kahn, R.; Fourme, R. Adaptation of the base-paired double-helix molecular architecture to extreme pressure. Nucleic Acids Res. 2007, 35, 4800–4808. [Google Scholar]
- Fourme, R.; Ascone, I.; Kahn, R.; Girard, E.; Mezouar, M.; Lin, T.; Johnson, J.; Winter, R. New trends in macromolecular crystallography at high hydrostatic pressure. In Adv. High Press. Sci. Technol. II; Springer: Heidelberg, Germany, 2003; pp. 161–170. [Google Scholar]
- Girard, E.; Kahn, R.; Mezouar, M.; Dhaussy, A.-C.; Lin, T.; Johnson, J.E.; Fourme, R. The first crystal structure of a macromolecular assembly under high pressure: CpMV at 330 MPa. Biophys. J. 2005, 88, 3562–3571. [Google Scholar] [CrossRef]
- Li, H.; Yamada, H.; Akasaka, K. Effect of pressure on individual hydrogen bonds in proteins. Basic pancreatic trypsin inhibitor. Biochemistry 1998, 37, 1167–1173. [Google Scholar]
- Williamson, M.P.; Akasaka, K.; Refaee, M. The solution structure of bovine pancreatic trypsin inhibitor at high pressure. Protein Sci. 2003, 12, 1971–1979. [Google Scholar] [CrossRef]
- Refaee, M.; Tezuka, T.; Akasaka, K.; Williamson, M.P. Pressure-Dependent changes in the solution structure of hen egg-white lysozyme. J. Mol. Biol. 2003, 327, 857–865. [Google Scholar]
- Wilton, D.J.; Tunnicliffe, R.B.; Kamatari, Y.O.; Akasaka, K.; Williamson, M.P. Pressure-induced changes in the solution structure of the GB1 domain of protein G. Proteins Struct. Funct. Bioinform. 2008, 71, 1432–1440. [Google Scholar]
- Wilton, D.J.; Ghosh, M.; Chary, K.; Akasaka, K.; Williamson, M.P. Structural change in a B-DNA helix with hydrostatic pressure. Nucleic Acids Res. 2008, 36, 4032–4037. [Google Scholar]
- Chalikian, T.V.; Sarvazyan, A.P.; Plum, G.E.; Breslauer, K.J. Influence of base composition, base sequence, and duplex structure on DNA hydration: apparent molar volumes and apparent molar adiabatic compressibilities of synthetic and natural DNA Duplexes at 25. degree. C. Biochemistry 1994, 33, 2394–2401. [Google Scholar]
- Lin, M.-C.; Eid, P.; Wong, P.T.; Macgregor, R.B. High pressure fourier transform infrared spectroscopy of poly (dA) poly (dT), poly (dA) and poly (dT). Biophys. Chem. 1999, 76, 87–94. [Google Scholar] [CrossRef]
- Chalikian, T.V.; Völker, J.; Plum, G.E.; Breslauer, K.J. A more unified picture for the thermodynamics of nucleic acid duplex melting: A characterization by calorimetric and volumetric techniques. Proc. Natl. Acad. Sci. USA 1999, 96, 7853–7858. [Google Scholar]
- Rayan, G.; Macgregor, R.B. Comparison of the heat-and pressure-induced helix-coil transition of two DNA copolymers. J. Phys. Chem. B 2005, 109, 15558–15565. [Google Scholar]
- Dubins, D.N.; Lee, A.; Macgregor, R.B.; Chalikian, T.V. On the stability of double stranded nucleic acids. J. Am. Chem. Soc. 2001, 123, 9254–9259. [Google Scholar]
- Macgregor, R.B. Chain length and oligonucleotide stability at high pressure. Biopolymers 1996, 38, 321–328. [Google Scholar]
- Rougee, M.; Faucon, B.; Mergny, J.; Barcelo, F.; Giovannangeli, C.; Garestier, T.; Helene, C. Kinetics and thermodynamics of triple-helix formation: Effects of ionic strength and mismatched. Biochemistry 1992, 31, 9269–9278. [Google Scholar]
- Lin, M.-C.; Macgregor, R.B. Activation volume of DNA duplex formation. Biochemistry 1997, 36, 6539–6544. [Google Scholar]
- Lin, M.-C.; Macgregor, R.B. The activation volume of a DNA helix-coil transition. Biochemistry 1996, 35, 11846–11851. [Google Scholar]
- Lin, M.C.; Macgregor, R.B. Pressure-jump relaxation kinetics of a DNA triplex helix-coil equilibrium. Biopolymers 1997, 42, 129–132. [Google Scholar]
- Dubins, D.N.; Macgregor, R.B. Volumetric properties of the formation of double stranded DNA: A nearest-neighbor analysis. Biopolymers 2004, 73, 242–257. [Google Scholar] [CrossRef]
- Sugimoto, N.; Nakano, S.-I.; Katoh, M.; Matsumura, A.; Nakamuta, H.; Ohmichi, T.; Yoneyama, M.; Sasaki, M. Thermodynamic parameters to predict stability of RNA/DNA hybrid duplexes. Biochemistry 1995, 34, 11211–11216. [Google Scholar]
- Freier, S.M.; Kierzek, R.; Jaeger, J.A.; Sugimoto, N.; Caruthers, M.H.; Neilson, T.; Turner, D.H. Improved free-energy parameters for predictions of RNA duplex stability. Proc. Natil. Acad. Sci. USA 1986, 83, 9373–9377. [Google Scholar]
- SantaLucia, J.; Allawi, H.T.; Seneviratne, P.A. Improved nearest-neighbor parameters for predicting DNA duplex stability. Biochemistry 1996, 35, 3555–3562. [Google Scholar] [CrossRef]
- Till, M.S.; Ullmann, G.M. McVol-A program for calculating protein volumes and identifying cavities by a Monte Carlo algorithm. J. Mol. Model. 2010, 16, 419–429. [Google Scholar] [CrossRef]
- Connolly, M.L. Solvent-Accessible surfaces of proteins and nucleic acids. Science 1983, 221, 709–713. [Google Scholar]
- Richards, F.M. Areas, volumes, packing and protein structure. Annu. Rev. Biophys. Bioeng. 1977, 6, 151–76. [Google Scholar] [CrossRef]
- Richards, F.M. Calculation of molecular volumes and areas for structures of known geometry. Methods Enzymol. 1985, 115, 440–464. [Google Scholar] [CrossRef]
- Robinson, C.R.; Sligar, S.G. Hydrostatic pressure reverses osmotic pressure effects on the specificity of EcoRI-DNA interactions. Biochemistry 1994, 33, 3787–3793. [Google Scholar] [CrossRef]
- Lynch, T.W.; Kosztin, D.; McLean, M.A.; Schulten, K.; Sligar, S.G. Dissecting the molecular origins of specific protein-nucleic acid recognition: Hydrostatic pressure and molecular dynamics. Biophys. J. 2002, 82, 93–98. [Google Scholar]
- Robinson, C.R.; Sligar, S.G. Heterogeneity in molecular recognition by restriction endonucleases: Osmotic and hydrostatic pressure effects on BamHI, Pvu II, and EcoRV specificity. Proc. Natl. Acad. Sci. USA 1995, 92, 3444–3448. [Google Scholar] [CrossRef]
- Royer, C.A.; Chakerian, A.E.; Matthews, K.S. Macromolecular binding equilibria in the lac repressor system: Studies using high-pressure fluorescence spectroscopy. Biochemistry 1990, 29, 4959–4966. [Google Scholar]
- Mohana-Borges, R.; Pacheco, A.B.; Sousa, F.J.; Foguel, D.; Almeida, D.F.; Silva, J.L. LexA repressor forms stable dimers in solution the role of specific DNA in tightening protein-protein interactions. J. Biol. Chem. 2000, 275, 4708–4712. [Google Scholar]
- Merrin, J.; Kumar, P.; Libchaber, A. Effects of pressure and temperature on the binding of RecA protein to single-stranded DNA. Proc. Natl. Acad. Sci. USA 2011, 108, 19913–19918. [Google Scholar] [CrossRef]
- Miyoshi, D.; Sugimoto, N. G-Quartet, G-Quadruplex, and G-Wire Regulated by Chemical Stimuli. Methods. Mol. Biol. 2011, 749, 93–104. [Google Scholar]
- Gellert, M.; Lipsett, M.N.; Davies, D.R. Helix formation by guanylic acid. Proc. Natl. Acad. Sci. USA 1962, 48, 2013–2018. [Google Scholar]
- Williamson, J.R.; Raghuraman, M.; Cech, T.R. Monovalent cation-induced structure of telomeric DNA: The G-quartet model. Cell 1989, 59, 871–880. [Google Scholar]
- Smith, F.W. Quadruplex structure of Oxytricha telomeric DNA oligonucleotides. Nature 1992, 356, 164–168. [Google Scholar]
- Davis, J.T. G-Quartets 40 Years Later: From 5'-GMP to molecular biology and supramolecular chemistry. Angew. Chem. Int. Ed. 2004, 43, 668–698. [Google Scholar] [CrossRef]
- Patel, D.J.; Phan, A.T.; Kuryavyi, V. Human telomere, oncogenic promoter and 5'-UTR G-quadruplexes: Diverse higher order DNA and RNA targets for cancer therapeutics. Nucleic Acids Res. 2007, 35, 7429–7455. [Google Scholar] [CrossRef]
- Heddi, B.; Phan, A.T. Structure of human telomeric DNA in crowded solution. J. Am. Chem. Soc. 2011, 133, 9824–9833. [Google Scholar]
- Zhang, D.-H.; Fujimoto, T.; Saxena, S.; Yu, H.-Q.; Miyoshi, D.; Sugimoto, N. Monomorphic RNA G-quadruplex and polymorphic DNA G-quadruplex structures responding to cellular environmental factors. Biochemistry 2010, 49, 4554–4563. [Google Scholar]
- Williamson, J.R. G-Quartet structures in telomeric DNA. Annu. Rev. Biophys. Biomol. Struct. 1994, 23, 703–730. [Google Scholar] [CrossRef]
- Petraccone, L.; Spink, C.; Trent, J.O.; Garbett, N.C.; Mekmaysy, C.S.; Giancola, C.; Chaires, J.B. Structure and stability of higher-order human telomeric quadruplexes. J. Am. Chem. Soc. 2011, 133, 20951–20961. [Google Scholar]
- Todd, A.K.; Johnston, M.; Neidle, S. Highly prevalent putative quadruplex sequence motifs in human DNA. Nucleic Acids Res. 2005, 33, 2901–2907. [Google Scholar]
- Huppert, J.L.; Balasubramanian, S. G-Quadruplexes in promoters throughout the human genome. Nucleic Acids Res. 2007, 35, 406–413. [Google Scholar] [CrossRef]
- Endoh, T.; Kawasaki, Y.; Sugimoto, N. Translational halt during elongation caused by G-quadruplex formed by mRNA. Methods 2013, in press. [Google Scholar]
- Endoh, T.; Kawasaki, Y.; Sugimoto, N. Stability of RNA quadruplex in open reading frame determines proteolysis of human estrogen receptor α. Nucleic Acids Res. 2013. in press.
- Moyzis, R.K.; Buckingham, J.M.; Cram, L.S.; Dani, M.; Deaven, L.L.; Jones, M.D.; Meyne, J.; Ratliff, R.L.; Wu, J.-R. A highly conserved repetitive DNA sequence,(TTAGGG) n, present at the telomeres of human chromosomes. Proc. Natl. Acad. Sci. USA 1988, 85, 6622–6626. [Google Scholar]
- Zahler, A.M.; Williamson, J.R.; Cech, T.R.; Prescott, D.M. Inhibition of telomerase by G-quartet DMA structures. Nature 1991, 350, 718–720. [Google Scholar]
- Yaku, H.; Fujimoto, T.; Murashima, T.; Miyoshi, D.; Sugimoto, N. Phthalocyanines: A new class of G-Quadruplex-Ligands with many potential applications. Chem. Commun. 2012, 48, 6203–6216. [Google Scholar]
- Cahoon, L.A.; Seifert, H.S. An alternative DNA structure is necessary for pilin antigenic variation in Neisseria gonorrhoeae. Science 2009, 325, 764–767. [Google Scholar] [CrossRef]
- Cheung, I.; Schertzer, M.; Rose, A.; Lansdorp, P.M. Disruption of dog-1 in Caenorhabditis elegans triggers deletions upstream of guanine-rich DNA. Nat. Genet. 2002, 31, 405–409. [Google Scholar]
- Rodriguez, R.; Miller, K.M.; Forment, J.V.; Bradshaw, C.R.; Nikan, M.; Britton, S.; Oelschlaegel, T.; Xhemalce, B.; Balasubramanian, S.; Jackson, S.P. Small-molecule–induced DNA damage identifies alternative DNA structures in human genes. Nat. Chem. Biol. 2012, 8, 301–310. [Google Scholar]
- Siddiqui-Jain, A.; Grand, C.L.; Bearss, D.J.; Hurley, L.H. Direct evidence for a G-quadruplex in a promoter region and its targeting with a small molecule to repress c-MYC transcription. Proc. Natl. Acad. Sci. USA 2002, 99, 11593–11598. [Google Scholar] [CrossRef]
- Hazel, P.; Huppert, J.; Balasubramanian, S.; Neidle, S. Loop-Length-Dependent folding of G-quadruplexes. J. Am. Chem. Soc. 2004, 126, 16405–16415. [Google Scholar]
- Rachwal, P.A.; Brown, T.; Fox, K.R. Effect of G-tract length on the topology and stability of intramolecular DNA quadruplexes. Biochemistry 2007, 46, 3036–3044. [Google Scholar] [CrossRef]
- Kankia, B.I.; Marky, L.A. Folding of the thrombin aptamer into a G-quadruplex with Sr2+: Stability, heat, and hydration. J. Am. Chem. Soc. 2001, 123, 10799–10804. [Google Scholar] [CrossRef]
- Olsen, C.M.; Marky, L.A. Energetic and hydration contributions of the removal of methyl groups from thymine to form uracil in G-quadruplexes. J. Phys. Chem. B 2008, 113, 9–11. [Google Scholar] [CrossRef]
- Miyoshi, D.; Karimata, H.; Sugimoto, N. Hydration regulates thermodynamics of G-quadruplex formation under molecular crowding conditions. J. Am. Chem. Soc. 2006, 128, 7957–7963. [Google Scholar] [CrossRef]
- Yu, H.; Gu, X.; Nakano, S.-I.; Miyoshi, D.; Sugimoto, N. Beads-on-a-string structure of long telomeric DNAs under molecular crowding conditions. J. Am. Chem. Soc. 2012, 134, 20060–20069. [Google Scholar]
- Fan, H.Y.; Shek, Y.L.; Amiri, A.; Dubins, D.N.; Heerklotz, H.; Macgregor, R.B., Jr.; Chalikian, T.V. Volumetric characterization of sodium-induced G-quadruplex formation. J. Am. Chem. Soc. 2011, 133, 4518–4526. [Google Scholar]
- Wang, Y.; Patel, D.J. Solution structure of the human telomeric repeat d[AG3(T2AG3)3] G-tetraplex. Structure 1993, 1, 263–282. [Google Scholar] [CrossRef]
- Edward, J.T.; Farrell, P.G. Relation between van der Waals and partial molal volumes of organic molecules in water. Can. J. Chem. 1975, 53, 2965–2970. [Google Scholar]
- Kharakoz, D. Partial molar volumes of molecules of arbitrary shape and the effect of hydrogen bonding with water. J. Solut. Chem. 1992, 21, 569–595. [Google Scholar]
- Likhodi, O.; Chalikian, T.V. Partial molar volumes and adiabatic compressibilities of a series of aliphatic amino acids and oligoglycines in D2O. J. Am. Chem. Soc. 1999, 121, 1156–1163. [Google Scholar] [CrossRef]
- Conway, B. The evaluation and use of properties of individual ions in slution. J. Solut. Chem. 1978, 7, 721–770. [Google Scholar] [CrossRef]
- Chalikian, T.V. Structural thermodynamics of hydration. J. Phys. Chem. B 2001, 105, 12566–12578. [Google Scholar] [CrossRef]
- Takahashi, S.; Sugimoto, N. Effect of pressure on the stability of G-quadruplex DNA: Thermodynamics under crowding conditions. Angew Chem. Int. Ed. in press.
- Zimmerman, S.B.; Minton, A.P. Macromolecular crowding: Biochemical, biophysical, and physiological consequences. Annu. Rev. Biophys. Biomol. Struct. 1993, 22, 27–65. [Google Scholar] [CrossRef]
- Minton, A.P. The influence of macromolecular crowding and macromolecular confinement on biochemical reactions in physiological media. J. Biol. Chem. 2001, 276, 10577–10580. [Google Scholar] [CrossRef]
- Ell is, R.J.; Minton, A.P. Join the crowd. Nature 2003, 425, 27–28. [Google Scholar]
- Nakano, S.; Karimata, H.; Ohmichi, T.; Kawakami, J.; Sugimoto, N. The effect of molecular crowding with nucleotide length and cosolute structure on DNA duplex stability. J. Am. Chem. Soc. 2004, 126, 14330–14331. [Google Scholar]
- Miyoshi, D.; Nakamura, K.; Tateishi-Karimata, H.; Ohmichi, T.; Sugimoto, N. Hydration of watson-crick base pairs and dehydration of hoogsteen base pairs inducing structural polymorphism under molecular crowding conditions. J. Am. Chem. Soc. 2009, 131, 3522–3531. [Google Scholar]
- Macaya, R.F.; Schultze, P.; Smith, F.W.; Roe, J.A.; Feigon, J. Thrombin-Binding DNA aptamer formsa unimolecular quadruplex structure in solution. Proc. Natl. Acad. Sci. USA 1993, 90, 3745–3749. [Google Scholar]
- Sasahara, K.; Sakurai, M.; Nitta, K. Volume and compressibility changes for short poly (ethylene glycol)-water system at various temperatures. Coll. Polym. Sci. 1998, 276, 643–647. [Google Scholar] [CrossRef]
- Sakurai, M. Partial molar volumes of ethylene glycol and water in their mixtures. J. Chem. Eng. Data 1991, 36, 424–427. [Google Scholar]
- Senior, M.M.; Jones, R.A.; Breslauer, K.J. Influence of loop residues on the relative stabilities of DNA hairpin structures. Proc. Natl. Acad. Sci. USA 1988, 85, 6242–6246. [Google Scholar]
- Varani, G.; Wimberly, B.; Tinoco, I., Jr. Conformation and dynamics of an RNA internal loop. Biochemistry 1989, 28, 7760–7772. [Google Scholar]
- Vallone, P.M.; Paner, T.M.; Hilario, J.; Lane, M.J.; Faldasz, B.D.; Benight, A.S. Melting studies of short DNA hairpins: Influence of loop sequence and adjoining base pair identity on hairpin thermodynamic stability. Biopolymers 1999, 50, 425–442. [Google Scholar] [CrossRef]
- Nakano, S.-i.; Hirayama, H.; Miyoshi, D.; Sugimoto, N. Dimerization of nucleic acid hairpins in the conditions caused by neutral cosolutes. J. Phys. Chem. B 2012, 116, 7406–7415. [Google Scholar]
- Chalikian, T.V.; Totrov, M.; Abagyan, R.; Breslauer, K.J. The hydration of globular proteins as derived from volume and compressibility measurements: Cross correlating thermodynamic and structural data. J. Mol. Biol. 1996, 260, 588–603. [Google Scholar] [CrossRef]
- Chalikian, T.V. Volumetric properties of proteins. Annu. Rev. Biophys. Biomol. Struct. 2003, 32, 207–235. [Google Scholar] [CrossRef]
- Fujimoto, T.; Nakano, S.-I.; Sugimoto, N.; Miyoshi, D. Thermodynamics-Hydration relationships within loops that affect G-quadruplexes under molecular crowding conditions. J. Phys. Chem. B 2012, 117, 963–972. [Google Scholar]
- Garcia, A.E.; Paschek, D. Simulation of the pressure and temperature folding/unfolding equilibrium of a small RNA hairpin. J. Am. Chem. Soc. 2008, 130, 815–817. [Google Scholar] [CrossRef]
- Muhuri, S.; Mimura, K.; Miyoshi, D.; Sugimoto, N. Stabilization of three-way junctions of DNA under molecular crowding conditions. J. Am. Chem. Soc. 2009, 131, 9268–9280. [Google Scholar]
- Rajendran, A.; Nakano, S.-I.; Sugimoto, N. Molecular crowding of the cosolutes induces an intramolecular i-motif structure of triplet repeat DNA oligomers at neutral pH. Chem. Commun. 2010, 46, 1299–1301. [Google Scholar] [CrossRef]
- Stewart, M.P.; Helenius, J.; Toyoda, Y.; Ramanathan, S.P.; Muller, D.J.; Hyman, A.A. Hydrostatic pressure and the actomyosin cortex drive mitotic cell rounding. Nature 2011, 469, 226–230. [Google Scholar]
- Kappor, S.; Werkmüller, A.; Goody, R.S.; Waldmann, H.; Winter, R. Pressure modulation of ras-membrane interactions and intervesicle transfer. J. Am. Chem. Soc. 2013, 135, 6149–6156. [Google Scholar]
© 2013 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Takahashi, S.; Sugimoto, N. Effect of Pressure on Thermal Stability of G-Quadruplex DNA and Double-Stranded DNA Structures. Molecules 2013, 18, 13297-13319. https://doi.org/10.3390/molecules181113297
Takahashi S, Sugimoto N. Effect of Pressure on Thermal Stability of G-Quadruplex DNA and Double-Stranded DNA Structures. Molecules. 2013; 18(11):13297-13319. https://doi.org/10.3390/molecules181113297
Chicago/Turabian StyleTakahashi, Shuntaro, and Naoki Sugimoto. 2013. "Effect of Pressure on Thermal Stability of G-Quadruplex DNA and Double-Stranded DNA Structures" Molecules 18, no. 11: 13297-13319. https://doi.org/10.3390/molecules181113297