Methylation of Dietary Flavones Increases Their Metabolic Stability and Chemopreventive Effects

Abstract
:
1. Introduction
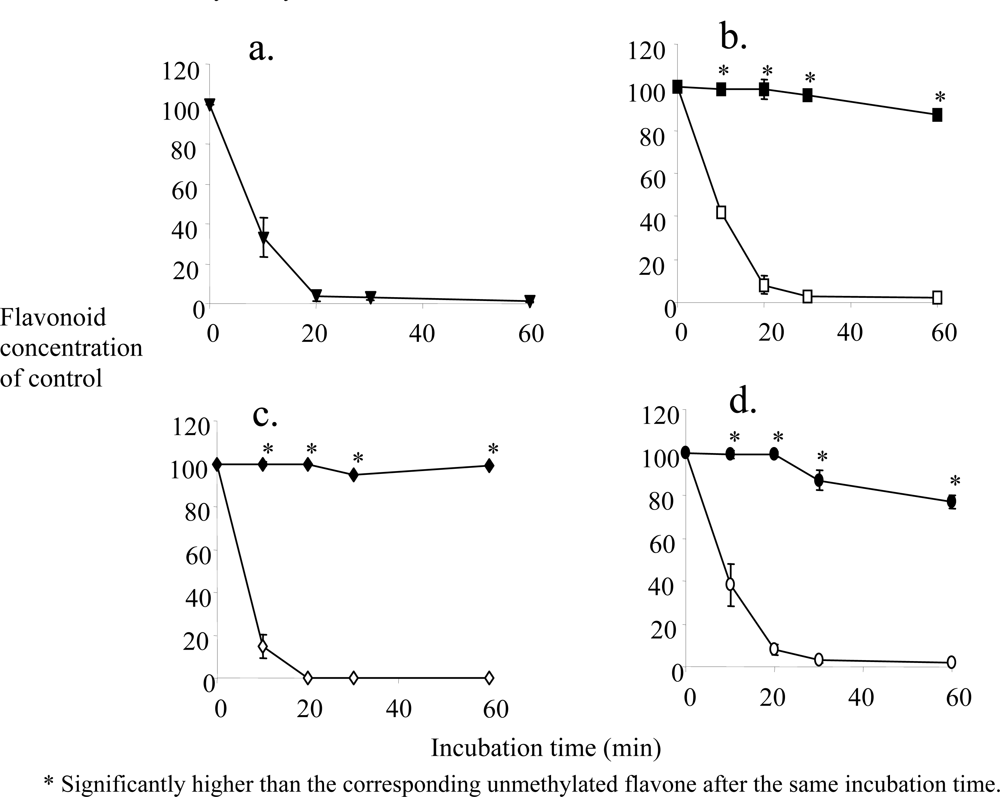
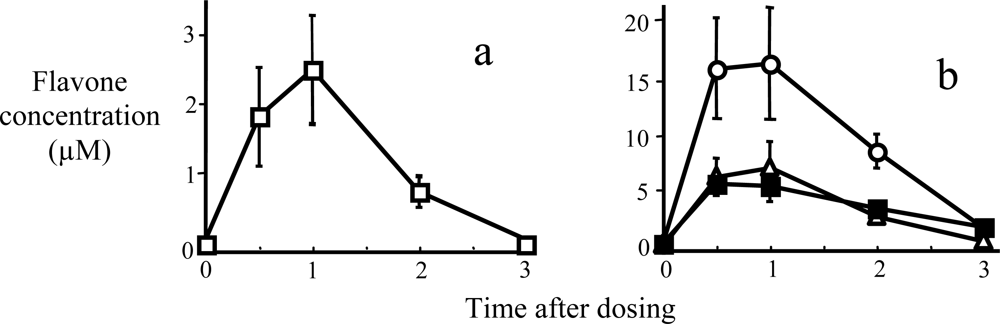
2. Metabolic Stability and Membrane Transport
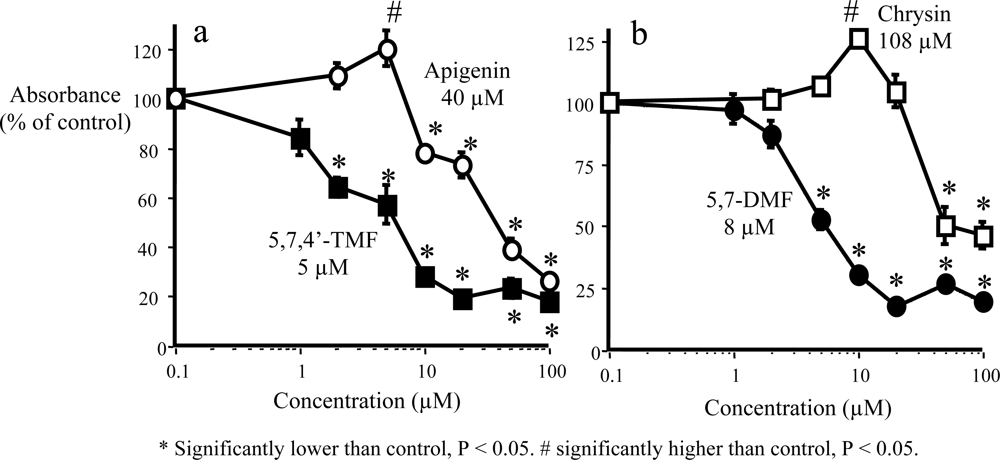
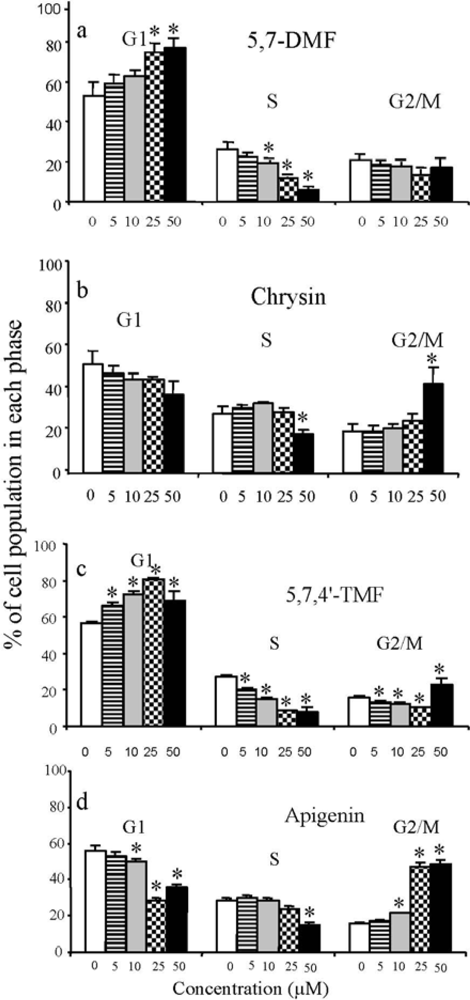
3. Anticancer and Hormonal Effects
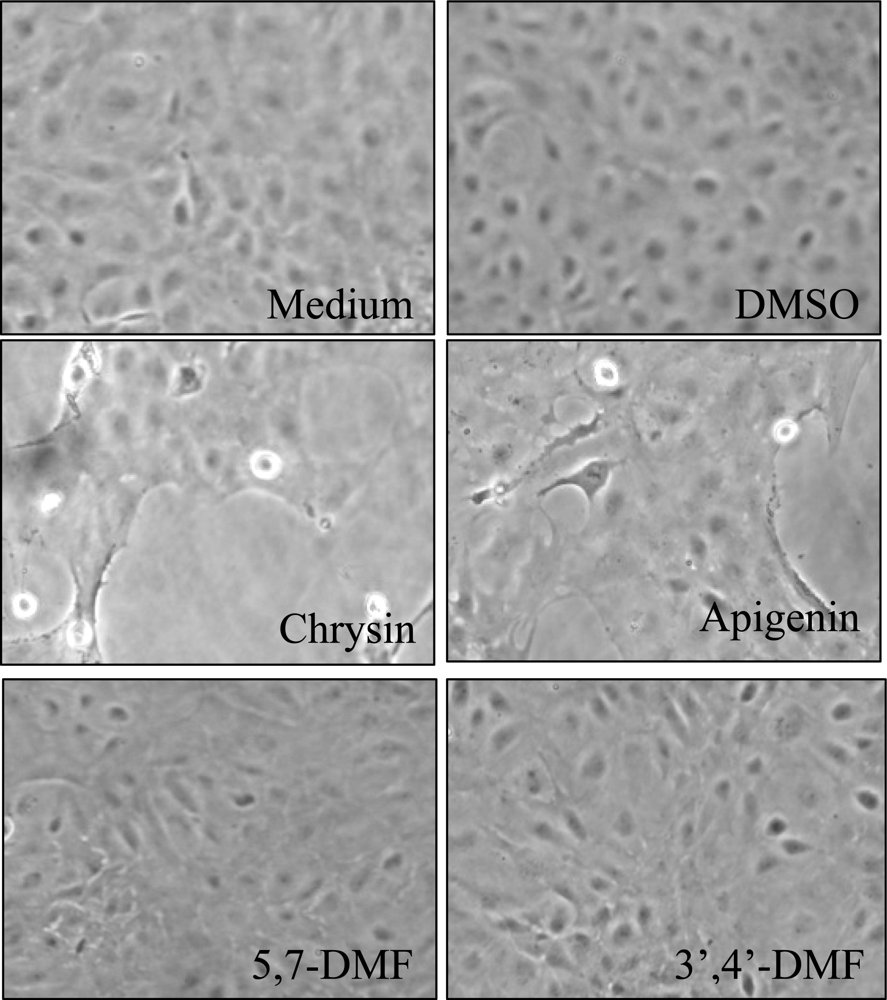
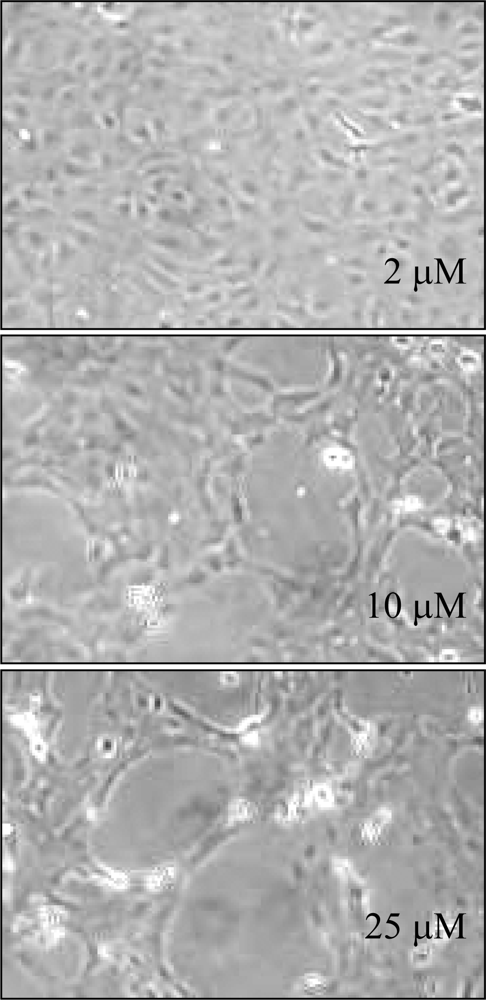
4. Toxic Side-Effects of Methylated Derivatives
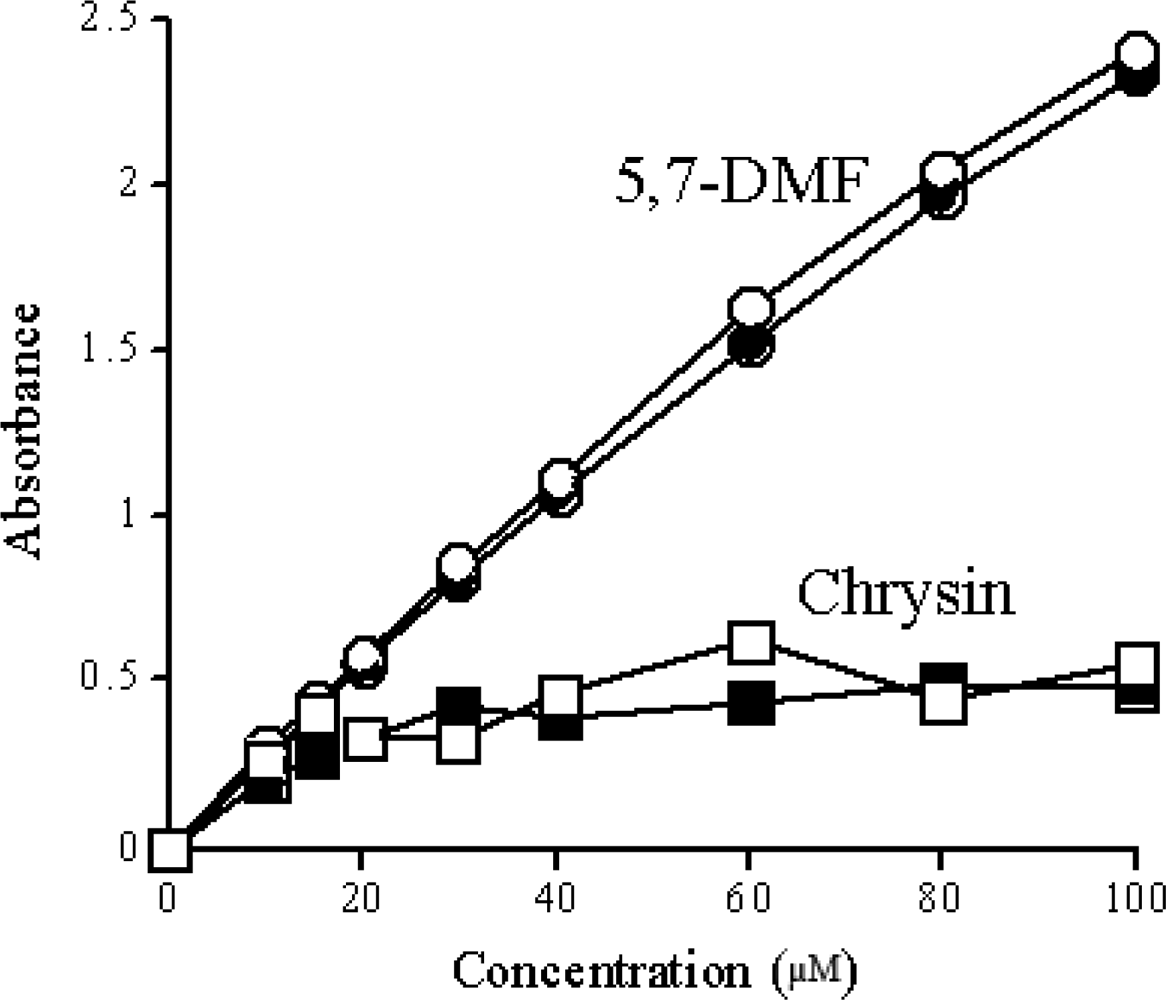
5. Solubility of Methylated Derivatives
6. Conclusions and Future Studies
Acknowledgments
References
- Middleton, EJ; Kandaswami, C; Theoharides, TC. The effects of plant flavonoids on mammalian cells: Implications for inflammation, heart disease, and cancer. Pharmacol. Rev 2000, 52, 673–751. [Google Scholar]
- Williams, RJ; Spencer, JPE; Rice-Evans, C. Flavonoids: Antioxidants and signalling molecules? Free Radic. Biol. Med 2004, 36, 838–849. [Google Scholar]
- Doostdar, H; Burke, MD; Mayer, RT. Bioflavonoids: Selective substrates and inhibitors for cytochrome P450 CYP1A and CYP1B1. Toxicology 2000, 144, 31–38. [Google Scholar]
- Guengerich, FP; Chun, Y-J; Kim, D; Gillam, EMJ; Shimada, T. Cytochrome P450 1B1: A target for inhibition in anticarcinogenesis strategies. Mut Res 2003, 523–524. [Google Scholar]
- Wen, X; Walle, T. Preferential induction of CYP1B1 by benzo[a]pyrene in human oral epithelial cells: Impact on DNA adduct formation and prevention by polyphenols. Carcinogenesis 2005, 26, 1774–1781. [Google Scholar]
- Wen, X; Walle, UK; Walle, T. 5,7-Dimethoxyflavone down-regulates CYP1A1 expression and benzo[a]pyrene-induced DNA binding in Hep G2 cells. Carcinogenesis 2005, 26, 803–809. [Google Scholar]
- Ciolino, HP; Daschner, PJ; Yeh, GC. Dietary flavonols quercetin and kaempferol are ligands of the aryl hydrocarbon receptor that affect CYP1A1 transcription differentially. Biochem. J 1999, 340, 715–722. [Google Scholar]
- Tsuji, PA; Walle, T. Inhibition of benzo[a]pyrene-activating enzymes and DNA-binding in human bronchial epithelial BEAS-2B cells by methoxylated flavonoids. Carcinogenesis 2006, 27, 1579–1585. [Google Scholar]
- Galijatovic, A; Otake, Y; Walle, UK; Walle, T. Induction of UDP-glucuronosyltransferase UGT1A1 by the flavonoid chrysin in Caco-2 cells—Potential role in carcinogen bioinactivation. Pharm. Res 2001, 18, 374–379. [Google Scholar]
- Gross-Steinmeyer, K; Stapleton, PL; Liu, F; Tracy, JH; Bammler, TK; Quigley, SD; Farin, FM; Buhler, DR; Safe, SH; Strom, SC; Eaton, DL. Phytochemical-induced changes in gene expression of carcinogen-metabolizing enzymes in cultured human primary hepatocytes. Xenobiotica 2004, 34, 619–632. [Google Scholar]
- Chou, FP; Chu, YC; Hsu, JD; Chiang, HC; Wang, CJ. Specific induction of glutathione S-transferase GSTM2 subunit expression by epigallocatechin gallate in rat liver. Biochem. Pharmacol 2000, 60, 643–650. [Google Scholar]
- Fang, J; Xia, C; Cao, Z; Zheng, JZ; Reed, E; Jiang, B-H. Apigenin inhibits VEGF and HIF-1 expression via PI3K/AKT/p70S6K1 and HDM2/p53 pathways. FASEB J 2005, 19, 342–353. [Google Scholar]
- Choi, H; Chun, Y-S; Kim, S-W; Kim, M-S; Park, J-W. Curcumin inhibits hypoxia-inducible factor-1 by degrading aryl hydrocarbon receptor nuclear translocator: A mechanism of tumor growth inhibition. Mol. Pharmacol 2006, 70, 1664–1671. [Google Scholar]
- Hou, Z; Sang, S; You, H; Lee, M-J; Hong, J; Chin, K-V; Yang, CS. Mechanism of action of (−)-epigallocatechin-3-gallate: Auto-oxidation-dependent inactivation of epidermal growth factor receptor and direct effects on growth inhibition in human esophageal cancer KYSE 150 cells. Cancer Res 2005, 65, 8049–8056. [Google Scholar]
- Lu, J; Papp, LV; Fang, J; Rodriguez-Nieto, S; Zhivotovsky, B; Holmgren, A. Inhibition of mammalian thioredoxin reductase by some flavonoids: Implications for myricetin and quercetin anticancer activity. Cancer Res 2006, 66, 4410–4418. [Google Scholar]
- Haghiac, M; Walle, T. Quercetin induces necrosis and apoptosis in the SCC-9 oral cancer cells. Nutr. Cancer 2005, 53, 220–231. [Google Scholar]
- Ermakova, SP; Kang, BS; Choi, BY; Choi, HS; Schuster, TF; Ma, W-Y; Bode, AM; Dong, Z. (−)-Epigallocatechin gallate overcomes resistance to etoposide-induced cell death by targeting the molecular chaperone glucose-regulated protein 78. Cancer Res 2006, 66, 9260–9268. [Google Scholar]
- Day, AJ; DuPont, MS; Ridley, S; Rhodes, M; Rhodes, MJC; Morgan, MRA; Williamson, G. Deglycosylation of flavonoid and isoflavonoid glycosides by human small intestine and liver β-glucosidase activity. FEBS Lett 1998, 436, 71–75. [Google Scholar]
- Day, AJ; Cañada, FJ; Diaz, JC; Kroon, PA; Mclauchlan, R; Faulds, CB; Plumb, GW; Morgan, MRA; Williamson, G. Dietary flavonoid and isoflavone glycosides are hydrolysed by the lactase site of lactase phlorizin hydrolase. FEBS Lett 2000, 468, 166–170. [Google Scholar]
- Graefe, EU; Wittig, J; Mueller, S; Riethling, A-K; Uehleke, B; Drewelow, B; Pforte, H; Jacobasch, G; Derendorf, H; Veit, M. Pharmacokinetics and bioavailability of quercetin glycosides in humans. J. Clin. Pharmacol 2001, 41, 492–499. [Google Scholar]
- Walle, T. Absorption and metabolism of flavonoids. Free Rad. Biol. Med 2004, 36, 829–837. [Google Scholar]
- Walle, T; Browning, AM; Steed, LS; Reed, SG; Walle, UK. Flavonoid glucosides are hydrolyzed and thus activated in the oral cavity in humans. J Nutr 2005, 135, 48–52. [Google Scholar]
- Manach, C; Donovan, JL. Pharmacokinetics and metabolism of dietary flavonoids in humans. Free Rad. Res 2004, 38, 771–785. [Google Scholar]
- Williamson, G; Manach, C. Bioavailability and bioefficacy of polyphenols in humans. II. Review of 93 intervention studies. Am. J. Clin. Nutr 2005, 81, 243S–255S. [Google Scholar]
- Walle, T; Otake, Y; Brubaker, JA; Walle, UK; Halushka, PV. Disposition and metabolism of the flavonoid chrysin in normal volunteers. Br. J. Clin. Pharmacol 2001, 51, 143–146. [Google Scholar]
- Goldberg, DM; Yan, J; Soleas, GJ. Absorption of three wine-related polyphenols in three different matrices by healthy subjects. Clin. Biochem 2003, 36, 79–87. [Google Scholar]
- Walle, T; Hsieh, F; DeLegge, MH; Oatis, JEJ; Walle, UK. High absorption but very low bioavailability of oral resveratrol in humans. Drug Metab. Dispos 2004, 32, 1377–1382. [Google Scholar]
- Walle, T; Walle, UK; Halushka, PV. Carbon dioxide is the major metabolite of quercetin in humans. J. Nutr 2001, 131, 2648–2652. [Google Scholar]
- Chow, HHS; Cai, Y; Alberts, DS; Hakim, I; Dorr, R; Shahi, F; Crowell, JA; Yang, CS; Hara, Y. Phase I pharmacokinetic study of tea polyphenols following single-dose administration of epigallocatechin gallate and polyphenol E. Cancer Epidemiol. Biomarkers Prev 2001, 10, 53–58. [Google Scholar]
- Warden, BA; Smith, LS; Beecher, GR; Balentine, DA; Clevidence, BA. Catechins are bioavailable in men and women drinking black tea throughout the day. J. Nutr 2001, 131, 1731–1737. [Google Scholar]
- Hong, J; Lambert, JD; Lee, S-H; Sinko, PJ; Yang, CS. Involvement of multidrug resistance-associated proteins in regulating cellular levels of (−)-epigallocatechin-3-gallate and its methyl metabolites. Biochem. Biophys. Res. Commun 2003, 310, 222–227. [Google Scholar]
- Vaidyanathan, JB; Walle, T. Cellular uptake of the anticancer tea flavonoid (−)-epicatechin-3-gallate (ECG) by an apical membrane transporter in Caco-2 cells. AAPS Annual Meeting, Toronto, Canada; 2002. [Google Scholar]
- Walton, MC; McGhie, TK; Reynolds, GW; Hendriks, WH. The flavonol quercetin-3-glucoside inhibits cyanidin-3-glucoside absorption in vitro. J. Agric. Food Chem 2006, 54, 4913–4920. [Google Scholar]
- Duarte Silva, I; Rodrigues, AS; Gaspar, J; Laires, A; Rueff, J. Metabolism of galangin by rat cytochromes P450: Relevance to the genotoxicity of galangin. Mutation Res 1997, 393, 247–257. [Google Scholar]
- Nielsen, SE; Breinholt, V; Justesen, U; Cornett, C; Dragsted, LO. In vitro biotransformation of flavonoids by rat liver microsomes. Xenobiotica 1998, 28, 389–401. [Google Scholar]
- Otake, Y; Walle, T. Oxidation of the flavonoids galangin and kaempferide by human liver microsomes and CYP1A1, CYP1A2, and CYP2C9. Drug Metab. Dispos 2002, 30, 103–105. [Google Scholar]
- Otake, Y; Hsieh, F; Walle, T. Glucuronidation versus oxidation of the flavonoid galangin by human liver microsomes and hepatocytes. Drug Metab. Dispos 2002, 30, 576–581. [Google Scholar]
- Breinholt, VM; Rasmussen, SE; Brøsen, K; Friedberg, TH. In vitro metabolism of genistein and tangeretin by human and murine cytochrome P450s. Pharmacol. Toxicol 2003, 93, 14–22. [Google Scholar]
- Hu, M; Krausz, K; Chen, J; Ge, X; Li, J; Gelboin, HL; Gonzalez, FJ. Identification of CYP1A2 as the main isoform for the phase I hydroxylated metabolism of genistein and a prodrug converting enzyme of methylated isoflavones. Drug Metab. Dispos 2003, 31, 924–931. [Google Scholar]
- Lu, Y; Zhang, C; Bucheli, P; Wei, D. Citrus flavonoids in fruit and traditional Chinese medicinal food ingredients in China. Plant Foods Hum. Nutr 2006, 61, 57–65. [Google Scholar]
- Nogata, Y; Sakamoto, K; Shiratsuchi, H; Ishii, T; Yano, M; Ohta, H. Flavonoid composition of fruit tissues of citrus species. Biosci. Biotechnol. Biochem 2006, 70, 178–192. [Google Scholar]
- Mizuno, M; Iinuma, M; Ohara, M; Tanaka, T; Iwamasa, M. Chemotaxonomy of the genus Citrus based on polymethoxyflavones. Chem. Pharm. Bull 1991, 39, 945–949. [Google Scholar]
- Jaipetch, T; Reutrakul, V; Tuntiwachwuttikul, P; Santisuk, T. Flavonoids in the black rhizomes of Boesenbergia pandurata. Phytochemistry 1983, 22, 625–626. [Google Scholar]
- Yenjai, C; Prasanphen, K; Daodee, S; Wongpanich, V; Kittakoop, P. Bioactive flavonoids from Kaempferia parviflora. Fitoterapia 2004, 75, 89–92. [Google Scholar]
- Cavalcante, SH; Fernandes, D; Paulino Fo, HF; Yoshida, M; Gottlieb, OR. Lignoids from the fruit of three Virola species. Phytochemistry 1985, 24, 1865–1866. [Google Scholar]
- Santos, LS; Corréa, MJC; Campos, LMO; Andrade, MA. Constituents from the leaves of Virola michelli. Fitoterapia 1996, 67, 555–556. [Google Scholar]
- Ahmad, F; Bakar, SA; Ibrahim, AZ; Read, RW. Constituents of the leaves of Piper caninum. Planta Med 1997, 63, 193–194. [Google Scholar]
- Walle, T. Methylation of dietary flavones greatly improves their hepatic metabolic stability and intestinal absorption. Mol. Pharmaceut 2007, 4, 826–832. [Google Scholar]
- Walle, T. Methoxylated flavones, a superior cancer chemopreventive flavonoid subclass? Sem. Cancer Biol 2007, 17, 354–362. [Google Scholar]
- Walle, T; Wen, X; Walle, UK. Improving metabolic stability of cancer chemopreventive polyphenols. Expert Opin. Drug Metab. Toxicol 2007, 3, 379–388. [Google Scholar]
- Wen, X; Walle, T. Methylated flavonoids have greatly improved intestinal absorption and metabolic stability. Drug Metab. Dispos 2006, 34, 1786–1792. [Google Scholar]
- Artursson, P; Karlsson, J. Correlation between oral drug absorption in humans and apparent drug permeability coefficients in human intestinal epithelial (Caco-2) cells. Biochem. Biophys. Res. Commun 1991, 175, 880–885. [Google Scholar]
- Walle, T; Walgren, RA; Walle, UK; Galijatovic, A; Vaidyanathan, JB. Understanding the bioavailability of flavonoids through studies in Caco-2 cells. In Flavonoids in Health and Disease; Rice-Evans, CA, Packer, L, Eds.; Marcel Dekker, Inc: New York, NY, USA, 2003; pp. 349–361. [Google Scholar]
- Walle, T; Ta, N; Kawamori, T; Wen, X; Tsuji, PA; Walle, UK. Cancer chemopreventive properties of orally bioavailable flavonoids—Methylated versus unmethylated flavones. Biochem. Pharmacol 2007, 73, 1288–1296. [Google Scholar]
- Walle, UK; Walle, T. Bioavailable flavonoids: Cytochrome P450-mediated metabolism of methoxyflavones. Drug Metab Dispos, in press.
- Manthey, JA; Guthrie, N; Grohmann, K. Biological properties of citrus flavonoids pertaining to cancer and inflammation. Curr. Med. Chem 2001, 8, 135–153. [Google Scholar]
- Manthey, JA; Guthrie, N. Antiproliferative activities of citrus flavonoids against six human cancer cell lines. J. Agric. Food Chem 2002, 50, 5837–5843. [Google Scholar]
- Chiang, L-C; Ng, LT; Lin, I-C; Kuo, P-L; Lin, C-C. Anti-proliferative effect of apigenin and its apoptotic induction in Hep G2 cells. Cancer Lett 2006, 237, 207–214. [Google Scholar]
- Trochon, V; Blot, E; Cymbalista, F; Engelmann, C; Tang, RP; Thomaidis, A; Vasse, M; Soria, J; Lu, H; Soria, C. Apigenin inhibits endothelial-cell proliferation in G(2)/M phase whereas it stimulates smooth-muscle cells by inhibiting P21 and P27 expression. Int. J. Cancer 2000, 85, 691–696. [Google Scholar]
- Ujiki, MB; Ding, X-Z; Salabat, MR; Bentrem, DJ; Golkar, L; Milam, B; Talamonti, MS; Bell, RHJ; Iwamura, T; Adrian, TE. Apigenin inhibits pancreatic cancer cell proliferation through G2/M cell cycle arrest. Mol. Cancer 2006, 5, 1–8. [Google Scholar]
- Pouget, C; Lauthier, F; Simon, A; Fagnere, C; Basly, J-P; Delage, C; Chulia, A-J. Flavonoids: Structural requirements for antiproliferative activity on breast cancer cells. Bioorg. Med. Chem. Lett 2001, 11, 3095–3097. [Google Scholar]
- Cai, H; Sale, S; Schmid, R; Britton, RG; Brown, K; Steward, WP; Gescher, AJ. Flavones as colorectal cancer chemopreventive agents—Phenyl-O-methylation enhances efficacy. Cancer Prevent. Res 2009, 2, 743–750. [Google Scholar]
- Stoner, G; Kaighn, M; Reddel, R; Resau, J; Bowman, D; Naito, Z; Matsukura, N; You, M; Galati, A; Harris, C. Establishment and characterization of SV40 T-antigen immortalized human esophageal epithelial cells. Cancer Res 1991, 51, 365–371. [Google Scholar]
- Reddel, RR; Ke, Y; Gerwin, BI; McMenamin, MG; Lechner, JF; Su, RT; Brash, DE; Park, J-B; Rhim, JS; Harris, CC. Transformation of human bronchial epithelial cells by infection with SV40 or adenovirus-12 SV40 hybrid virus, or transfection via strontium phosphate coprecipitation with a plasmid containing SV40 early region genes. Cancer Res 1988, 48, 1904–1909. [Google Scholar]
- Bock, KW; Köhle, C. Ah receptor- and TCDD-mediated liver tumor promotion: Clonal selection and expansion of cells evading growth arrest and apoptosis. Biochem. Pharmacol 2005, 69, 1403–1408. [Google Scholar]
- Levine-Fridman, A; Chen, L; Elferink, CJ. Cytochrome P4501A1 promotes G1 phase cell cycle progression by controlling aryl hydrocarbon receptor activity. Mol. Pharmacol 2004, 65, 461–469. [Google Scholar]
- Wen, X; Walle, T. Cytochrome P450 1B1, a biomarker and chemopreventive target for benzo[a] pyrene-initiated human esophageal cancer. Cancer Lett 2006, 246, 109–114. [Google Scholar]
- Ta, N; Walle, T. Aromatase inhibition by bioavailable methylated flavones. J. Steroid Biochem. Mol. Biol 2007, 107, 127–129. [Google Scholar]
- Kao, Y-C; Zhou, C; Sherman, M; Laughton, CA; Chen, S. Molecular basis of the inhibition of human aromatase (estrogen synthetase) by flavone and isoflavone phytoestrogens: A site-directed mutagenesis study. Environ. Health Perspect 1998, 106, 85–92. [Google Scholar]
- Wen, X; Walle, T. Methylation protects dietary flavonoids from rapid hepatic metabolism. Xenobiotica 2006, 36, 387–397. [Google Scholar]
- Galati, G; O'Brien, PJ. Potential toxicity of flavonoids and other dietary phenolics: Significance for their chemopreventive and anticancer properties. Free Rad. Biol. Med 2004, 37, 287–303. [Google Scholar]
- Chan, TS; Galati, G; Pannala, AS; Rice-Evans, CA; O'Brien, PJ. Simultaneous detection of the antioxidant and pro-oxidant activity of dietary polyphenolics in a peroxidase system. Free Rad. Biol. Med 2003, 37, 787–794. [Google Scholar]
- Awad, HM; Boersma, MG; Boeren, S; van Bladeren, PJ; Vervoort, J; Rietjens, IMCM. Quenching of quercetin quinone/quinone methides by different thiolate scavengers: Stability and reversibility of conjugate formation. Chem. Res. Toxicol 2003, 16, 822–831. [Google Scholar]
- Walle, T; Vincent, TS; Walle, UK. Evidence of covalent binding of the dietary flavonoid quercetin to DNA and protein in human intestinal and hepatic cells. Biochem. Pharmacol 2003, 65, 1603–1610. [Google Scholar]
- van der Woude, H; Alink, GM; van Rossum, BEJ; Walle, UK; van Steeg, H; Walle, T; Rietjens, IMCM. Formation of transient covalent protein and DNA adducts by quercetin in cells with and without oxidative enzyme activity. Chem. Res. Toxicol 2005, 18, 1907–1916. [Google Scholar]
- Tsuji, PA; Walle, T. Cytotoxic effects of the dietary flavones chrysin and apigenin in a normal trout liver cell line. Chem.—Biol. Interactions 2008, 171, 37–44. [Google Scholar]
- Goss, LB; Sabourin, TD. Utilization of alternative species for toxicity testing. J. Appl. Toxicol 1985, 5, 193–219. [Google Scholar]
- Lee, J-E; Safe, S. 3′,4′-Dimethoxyflavone as an aryl hydrocarbon receptor antagonist in human breast cancer cells. Toxicol. Sci 2000, 58, 235–242. [Google Scholar]
- Ueno, I; Nakano, N; Hirono, I. Metabolic fate of [14C]quercetin in the ACI rat. Jap. J. Exp. Med 1983, 53, 41–50. [Google Scholar]
- Gugler, R; Leschik, M; Dengler, HJ. Disposition of quercetin in man after single oral and intravenous doses. Eur. J. Clin. Pharmacol 1975, 9, 229–234. [Google Scholar]
- Walle, T; Walle, UK. Medical University of South Carolina, Charleston, SC, USA. Unpublished work, 2008.



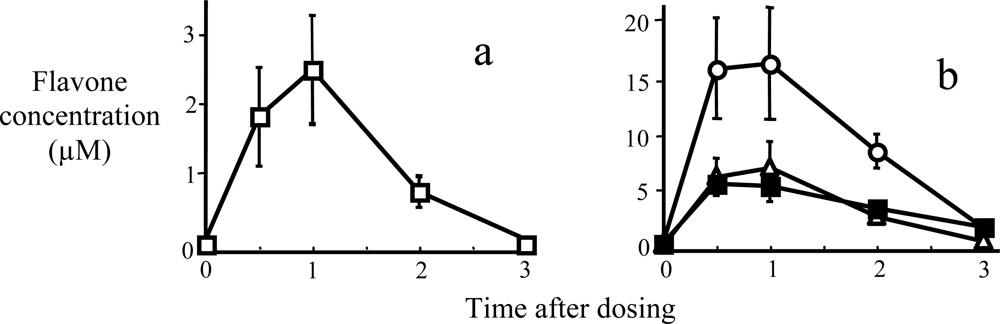






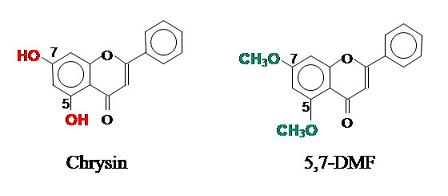{kind=link}
{kind=link}
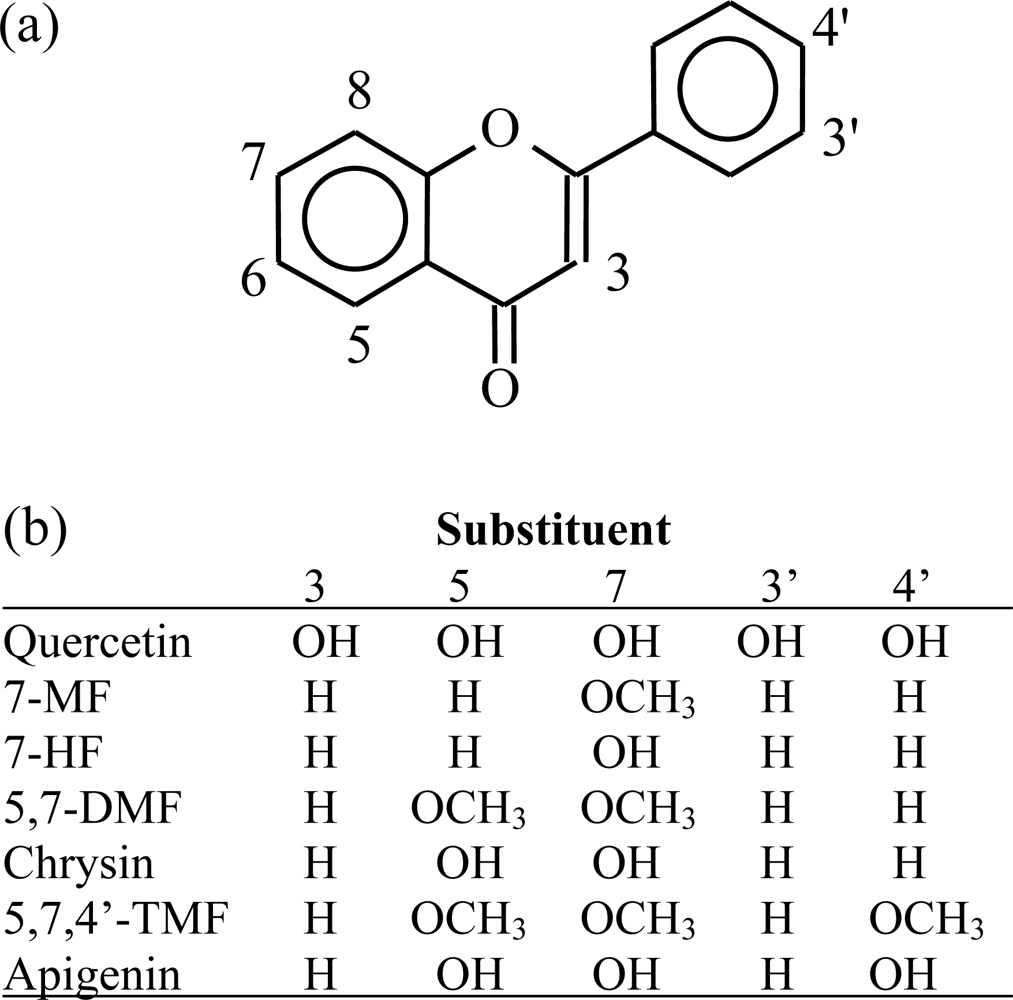{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Flavone | Half-life (min) | Clint (ml min−1 kg−1) |
|---|---|---|
| 5,7-DMF | 97.1 | 12.8 |
| 5-MF | 68.2 | 18.3 |
| Sinensetin | 44.8 | 27.8 |
| 5,7,4’-TMF | 33.3 | 37.4 |
| 3’,4’-DMF | 30.9 | 40.3 |
| 7,4’-DMF | 28.6 | 43.6 |
| 5,3’-DMF | 20.8 | 60.0 |
| Tangeretin | 17.4 | 71.7 |
| 7-MF | 15.7 | 79.4 |
| Kaempferide | 15.2 | 82.0 |
| 7,3’-DMF | 13.6 | 91.7 |
| 5,4’-DMF | 10.5 | 119.0 |
| 3’-MF | 8.9 | 140.0 |
| 4’-MF | 7.8 | 161.0 |
| Tectochrysin | 4.4 | 283.0 |
© 2009 by the authors; licensee Molecular Diversity Preservation International, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Walle, T. Methylation of Dietary Flavones Increases Their Metabolic Stability and Chemopreventive Effects. Int. J. Mol. Sci. 2009, 10, 5002-5019. https://doi.org/10.3390/ijms10115002
Walle T. Methylation of Dietary Flavones Increases Their Metabolic Stability and Chemopreventive Effects. International Journal of Molecular Sciences. 2009; 10(11):5002-5019. https://doi.org/10.3390/ijms10115002
Chicago/Turabian StyleWalle, Thomas. 2009. "Methylation of Dietary Flavones Increases Their Metabolic Stability and Chemopreventive Effects" International Journal of Molecular Sciences 10, no. 11: 5002-5019. https://doi.org/10.3390/ijms10115002