Exploitation of SPR to Investigate the Importance of Glycan Chains in the Interaction between Lactoferrin and Bacteria


1
Biosciences Department, Teagasc Food Research Centre, Moorepark, Fermoy, P61C996 Co. Cork, Ireland
2
Glycoscience Group, National Centre for Biomedical Engineering Science, National University of Ireland Galway, H91TK33, Galway, Ireland
*
Author to whom correspondence should be addressed.
Sensors 2017, 17(7), 1515; https://doi.org/10.3390/s17071515
Submission received: 15 May 2017
/
Revised: 21 June 2017
/
Accepted: 22 June 2017
/
Published: 27 June 2017
(This article belongs to the Special Issue Sensors for Glycoproteins and Glycated Proteins)
Abstract
:Bovine lactoferrin (LF) has been shown to prevent adhesion to and invasion of mammalian cell lines by pathogenic bacteria, with evidence for direct bacterial binding by the milk glycoprotein. However, the glycosylation pattern of LF changes over the lactation cycle. In this study, we aim to investigate the effect that this variation has on the milk glycoprotein’s ability to interact with pathogens. Surface plasmon resonance technology was employed to compare the binding of LF from colostrum (early lactation) and mature milk (late lactation) to a panel of pathogenic bacteria (Staphylococcus aureus, Escherichia coli, Cronobacter sakazakii, Streptococcus pneumoniae, Pseudomonas aeruginosa, Listeria monocytogenes and Salmonella typhimurium). Novel interactions with LF were identified for C. sakazakii, S. pneumoniae and P. aeruginosa with the highest binding ability observed for mature milk LF in all cases, with the exception of S. typhimurium. The difference in bacterial binding observed may be as a result of the varying glycosylation profiles. This work demonstrates the potential of LF as a functional food ingredient to prevent bacterial infection.
1. Introduction
Bovine milk lactoferrin (LF) is a single chain, iron-binding, glycosylated protein [1] present in the whey protein fraction of milk [2] (http://www.uniprot.org/uniprot/B9VPZ5). There are two lactoferrin variants present in bovine milk: LF-a and LF-b. Five glycosylation potential sites (Asn233, Asn281, Asn368, Asn476 and Asn545) were identified through a combination of digestion and HPLC on LF, of which four are always utilised but Asn281 is glycosylated only on LF-a [3,4,5]. Lactoferrin may have high-mannose type, complex type or hybrid type N-linked glycans attached. The associated glycan chains are composed of N-acetyl-glucosamine (GlcNAc), galactose (Gal), N-acetyl-galactosamine (GalNAc), fucose (Fuc), mannose (Man), N-acetyl-neuraminic acid (Neu5Ac) and N-glycolyl-neuraminic acid (Neu5Gc) (as previously reviewed [6]) and variation in the glycosylation pattern of LF has been described over the lactation cycle [7]. A greater degree of heterogeneity in glycan structure has been reported for LF from early lactation, with an abundance of high-mannose type glycans present on th glycoprotein in mature milk [7]. The glycan chains in mature milk LF are 65% oligomannose type, consisting of multiple isomers of high-mannose type glycans, containing five to nine mannose residues, 38% of the total mannose structures accounted for by Man8 [8]. Neu5Gc was shown to be present on bovine LF only in the initial days postpartum and the total sialic acid content decreased as lactation progressed [7].
LF has a wide variety of associated biological activities including antimicrobial [9], immunomodulatory [10], prebiotic [11,12], stimulation of bone formation [13] and anticancer [14,15]. These bioactivities have previously been reviewed by several authors [16,17] and the glycan component of LF has an important role to play in many of these properties [6]. The presence of the glycan chains influences the tertiary structure of the protein, and enhances its ion binding ability [18,19,20] and its resistance to enzyme digestion [21]. The attached glycans are also believed to have a role in LF’s antibacterial [22,23,24] and antiviral [25,26] activities. However, to date, information on how glycosylation variation over lactation influences bioactivity is limited.
Biomolecular interaction analysis (Biacore) involves the use of surface plasmon resonance (SPR) to measure the binding of candidate compounds or cells to specific ligands. SPR measures the changes in the refractive index near a planar chip surface induced by binding of soluble molecules to immobilized counterpart molecules on the sensor chip [27]. It has been widely used for the quantification and kinetic analysis of receptor-ligand interactions [28,29,30,31,32,33]. Recently, this methodology has been identified as a reliable, high throughput method for profiling the interaction of whole bacterial cells with an immobilised glycans [28]. The authors validated the Biacore assay by investigating the interaction between Campylobacter jejuni and the milk oligosaccharide, 2’-fucosyllactose, to which it is known to bind [34,35]. The assay monitors interactions dynamically over time in a continuous flow system and may mimic natural conditions in a more realistic manner when compared with static adhesion assays [28]. A number of similar studies to monitor whole bacterial cell interactions with immobilised compounds of interest have also been documented [36,37,38,39]. Of particular interest is a study using SPR to distinguish between five closely related Escherichia coli strains, including the enterohemorragic E. coli O157:H7 based on direct differential carbohydrate recognitions [40].
The current study aims to exploit SPR to investigate the importance of the glycan chains in the interaction between LF and bacteria. Commercially available LF from colostrum and mature bovine milk was biotinylated and immobilised on the streptavidin coated surface of a Streptavidin SA chip. A number of pathogenic bacterial strains were selected and screened for interactions with both glycovariants.
2. Materials and Methods
2.1. Materials
LF from colostrum and mature milk was purchased from Sigma-Aldrich Co. (Dublin, Ireland). Mueller-Hinton broth and Brain Heart Infusion broth were purchased from Oxoid (Basingstoke, Hampshire, UK). The Biacore X instrument, SA chip, biotin CAPture kit, HBS-EP buffer (10 mmol·L−1 Hepes, 150 mmol·L−1 NaCl, 3.8 mmol·L−1 ethylenediaminetetraacetic acid (EDTA), 0.05% (v/v) Tween 20), and amine coupling kit were purchased from GE Healthcare (Buckinghamshire, UK). The EZ-Link biotin-PEG4-hydrazide kit, sodium acetate buffer (pH 5.5), and Zeba desalt spin column were purchased from Pierce (Rockford, IL, USA). Sodium meta-periodate was purchased from Sigma-Aldrich Co. (Dublin, Ireland).
2.2. Bacterial Strains and Culture Conditions
Bacterial strains used in this study and their respective growth media are listed in Table 1.
Bacterial culture stocks were maintained in their respective growth media containing 50% glycerol at −80 °C in the culture collection at Teagasc Food Research Centre, Moorepark, Fermoy, P61C996, Co. Cork, Ireland and propagated twice prior to use. All bacterial strains were grown aerobically for 12–24 h at 37 °C. Bacterial cells were washed three times in HBS–EP buffer and re-suspended to a concentration of 1 × 108 colony-forming units (CFU) mL−1 for screening studies unless otherwise stated.
2.3. Biotinylation of Lactoferrin
LF samples were biotinylated using EZLink biotin-PEG4-hydrazide as per the manufacturer’s instructions. Briefly, 2 mg of LF was dissolved in 1 mL of 0.1 mol·L−1 sodium acetate buffer (pH 5.5). An amount of 1 mL of cold sodium meta-periodate solution (20 mmol·L−1 periodate) was added and the solution was mixed well. The mixture was protected from light and incubated for 30 min at room temperature. Excess periodate was removed using a Zeba desalt spin column equilibrated with 0.1 mol·L−1 sodium acetate buffer (pH 5.5). One part of 50 mmol L−1 biotin-hydrazide solution was added to nine parts of the treated sample and incubated for 3–4 h at room temperature. Unbound biotin remaining in the sample was removed by passing the solution sequentially through two Zeba desalt spin columns. The sample was then stored at 4 °C until use.
2.4. Biacore Assay
The entire analysis was carried out on a Biacore X instrument at a constant temperature (25 °C) and flow rate (10 µL·min−1), unless otherwise stated, using HBS–EP as the run buffer and a SA chip. The streptavidin-coated SA chip surface was primed with a short injection of 1 mol·L−1 NaCl and 50 mmol·L−1 NaOH (filtered and degassed). Whole bacterial cells were harvested and resuspended in HBS–EP running buffer as described in Section 2.2. In order to confirm the absence of non-specific binding of the selected bacterial strains to the SA chip surface, cell suspensions (1 × 108 CFU mL−1) were injected over the chip surface at 10·µL·min−1, and the binding signal was measured. Signal change was reported in response units (RU). The chip surface was washed with HBS–EP running buffer between bacterial injections to ensure full removal of microbial cells.
Thereafter, biotin-labelled LF (from either colostrum or mature milk) was diluted in HBS–EP buffer (50 µg·mL−1), and 100 µL of this solution was injected over the surface at a flow rate of 10 µL·min−1. The chip surface was then washed with HBS–EP buffer to ensure the removal of any non-immobilized molecules. The RU increase following each LF injection was monitored to ensure comparable levels of analyte were immobilised. Subsequently, bacterial injections were repeated to evaluate bacterial binding to immobilised colostrum and mature milk LF.
2.5. Statistical Analysis
All experiments were performed in triplicate and results are presented as mean values ± standard deviations of three replicate experiments. Nonspecific binding of the analyte to the test surface was eliminated from all experiments through the use of a reference surface.
3. Results
In the current study, SPR was used to investigate the effect of changes in LF glycosylation over lactation on its ability to bind to pathogenic bacteria. A panel of pathogenic bacteria was initially exposed to the surface of the SA chip, minus the immobilised analyte, to ensure the absence of non-specific binding to the chip surface, which could result in false results during the test runs. Bacterial solutions were washed and resuspended in buffer prior to exposure to the chip surface to ensure no salts were present in the samples. The chip was also washed between exposures to different bacterial strains and also on a daily basis. The system was also optimized in terms of the flow rate to ensure that cell deposition did not occur. For all bacteria screened, non-specific binding was not observed (data not shown). Concentrations of bacteria at approximately 1 × 108 CFU·mL−1 were used in the assays as previous studies have shown that concentrations in this range are generally required to generate a measurable signal [28].
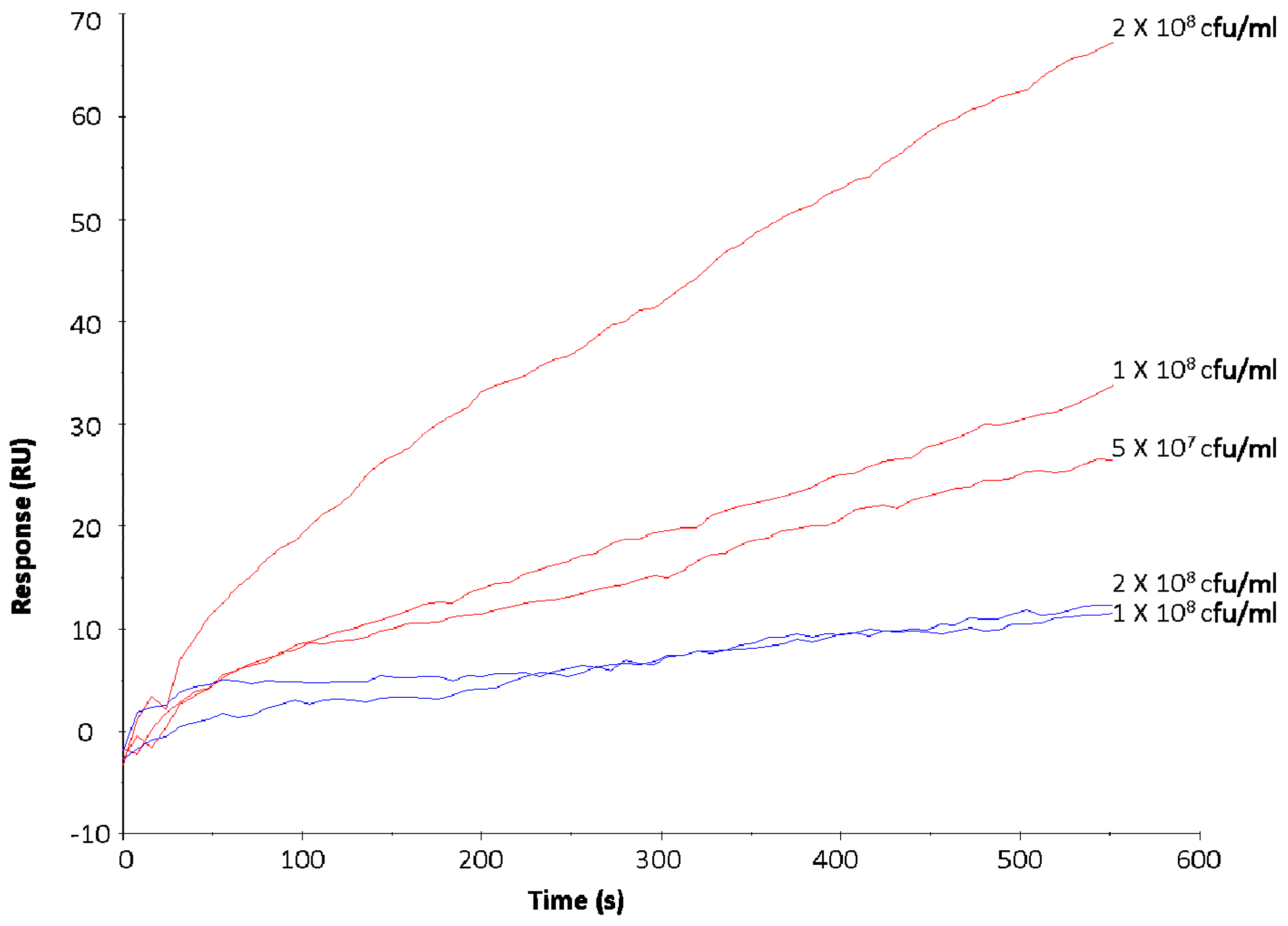
Based on previous studies [23,41,42] and the results generated in the initial phase of this work, S. aureus DPC 5971 and E. coli P1432 were identified as positive and negative controls for binding to LF respectively. Exposure of bacterial suspensions with increasing cell number resulted in an increase in the RU value with S. aureus DPC 5971, confirming that the injected bacterial cells were interacting with the immobilised LF (Figure 1). It should be noted that at the end of the injection period of 600 s, the RU values were maintained before a gradual decline in RU was observed, indicating the specific nature of the interactions (data not shown).
No increase in RU value was observed for E. coli P1432 following exposure of increasing concentrations of bacterial cells to immobilised LF (Figure 1), confirming the lack of binding of this strain to the analyte. These strains were exposed to the surface of the SA chip with immobilised LF at the beginning of each experimental set to ensure consistent performance of the chip. The response of E. coli P1432 (colostrum: 16.8 ± 4.3 RU; mature milk: 17 ± 7.1) was considered a baseline minimum RU value and used as a reference for screening the other pathogens for positive interactions.
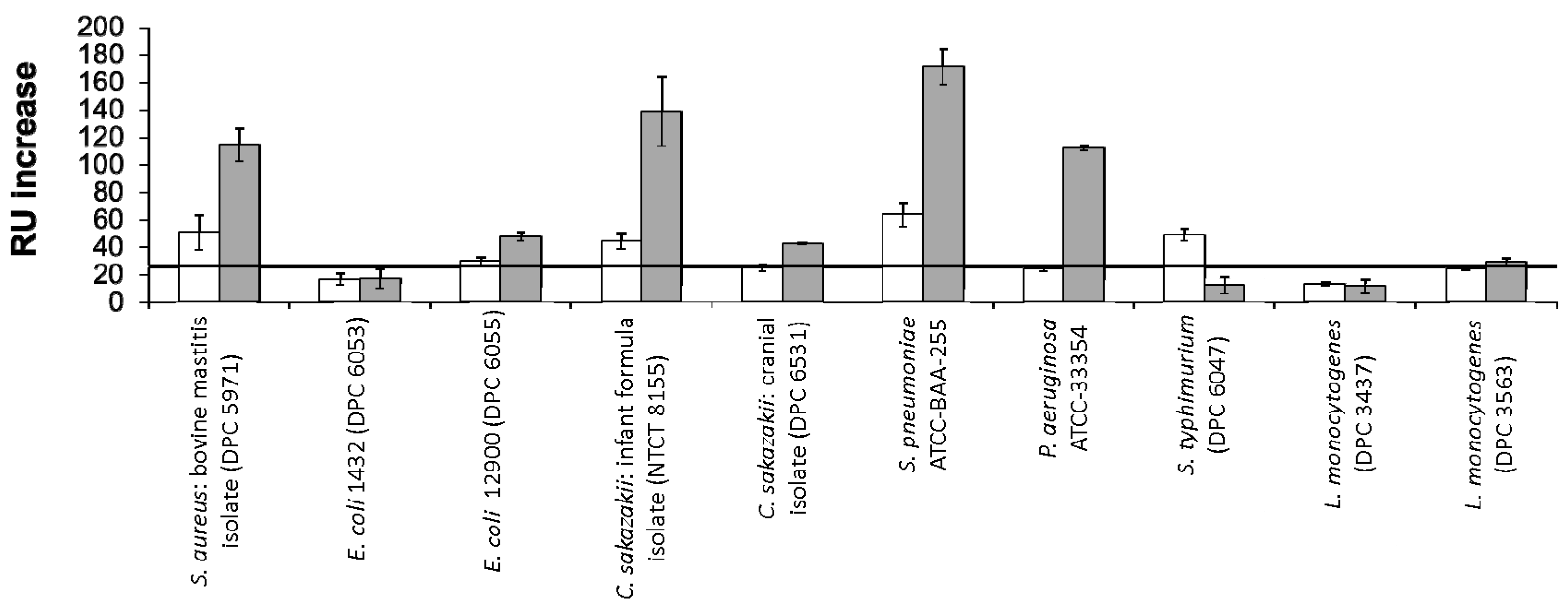
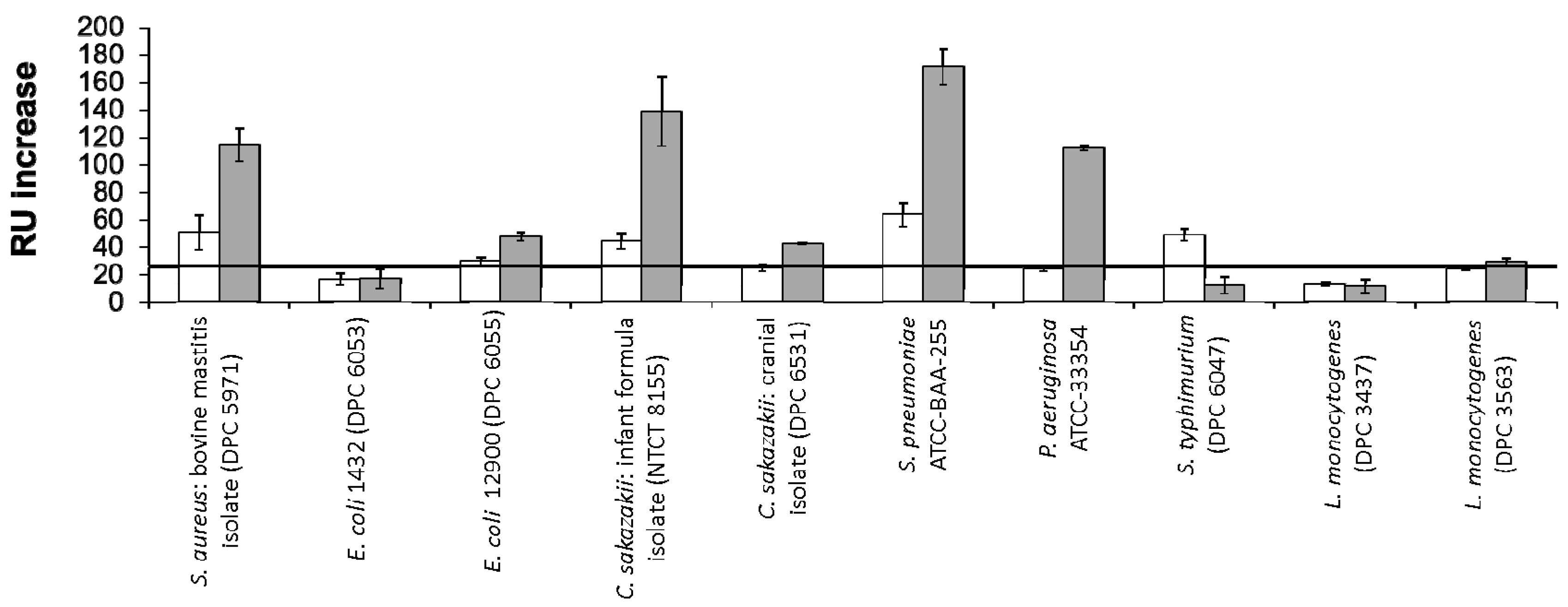
RU changes following exposure of the bacterial strains to the immobilised colostrum and mature milk LF are shown in Figure 2. Of the positive interactions profiled, the strongest signals were observed for C. sakazakii NCTC 8155, S. pneumoniae ATCC BAA-255 and P. aeruginosa ATCC 33354. Neither L. monocytogenes strain tested displayed binding to either LF glycovariant. For the majority of strains which displayed positive binding, a stronger interaction was observed for mature milk LF versus colostrum LF. S. typhimurium ATCC BAA-185 was the sole exception to this, interacting only with the glycovariant of the protein isolated from colostrum.
4. Discussion
The greater interaction with mature LF may be linked to the abundance of high-mannose type glycans on this glycovariant. Teraguchi, Shin, Fukuwatari and Shimamura [23] described the relationship between LF and members of the Escherichia family and the dependence of the observed interaction on the presence of high-mannose type glycans on the milk protein. Members of this family which express type 1 fimbriae can recognise and bind to oligomannose glycan chains on eukaryotic cell surfaces [43], which can facilitate bacterial adhesion to and invasion of the cells. Teraguchi et al. [23] demonstrated that the high mannose-type glycans on bovine LF acted as receptors for the mannose specific type 1 fimbriae, therefore preventing bacterial interaction with the eukaryotic cell by acting as a decoy receptor. When human LF was tested, this activity was not observed, potentially as a result of the presence of only complex type glycans on the human protein variant. The same group went on to demonstrate that bovine LF caused the agglutination of type 1 fimbriated E. coli cells as a result of the specific interaction between the mannose residues on the glycoprotein and the type 1 fimbriae of the bacteria. The results presented here suggest that bovine LF may also have similar interactions with other pathogenic bacteria as a result of its direct bacterial interaction via its oligomannose type glycans. The glycoprotein may have a role as a more non-specific defence mechanism, inhibiting bacterial adhesion to mammalian cells.
The lower binding observed for LF isolated from early lactation may also be linked to the presence of more diverse antimicrobial elements in colostrum such as immunoglobulins and oligosaccharides. Therefore, there may be less of a biological requirement for LF to contribute to the inhibition of pathogenic infection. Also, the presence of sialic acid on colostrum LF glycans suggests that this glycovariant may have an alternative method of anti-bacterial activity in early lactation. Sialylation has previously been linked to LF’s antimicrobial activity as a result of its calcium chelation activity, competing for loosely bound calcium ions involved in the stabilization of lipopolysaccharides in the outer membrane of bacterial cells [18,44].
S. aureus DPC 5971 is a bovine mastitis isolate [45] and was selected as the positive control for this study as receptors for LF have previously been identified in S. aureus strains associated with mastitis infection [41]. The LF concentration in milk has been shown to increase in cows suffering from subclinical mastitis [46], possibly as part of an immune response to infection. The results of this study demonstrate that although S. aureus DPC 5971 binds to both colostrum and mature milk LF, the glycoprofile present on mature milk LF is more favourable for bacterial binding, suggesting that the S. aureus receptors which interact with LF are regulated by the glycosylation present on the protein.
Two E. coli O157:H7 strains were included in this study: E. coli P1432 and E. coli NCTC 12900; the former was included as the negative control. O157 strains have previously been shown to be unable to produce type 1 fimbriae (because of a deletion in the fim regulatory region) [42] described by Teraguchi, Shin, Fukuwatari and Shimamura [23] as the mode of binding for E. coli cells to LF mannose residues. Of interest, a recent study proposes a method for pathotyping E. coli based on type 1 fimbrial interaction with a mannosylated surface using shear force [47]. Minimal binding was observed for E. coli NCTC 12900 with mature milk LF. LF has been shown to bind to different E. coli strains with varying levels of efficiency, for both the human and bovine variants of the protein [48,49].
Two C. sakazakii strains were exposed to immobilised LF in this study: C. sakazakii NCTC 8155, a strain isolated from dried milk powder; and C. sakazakii DPC 6531, a brain tumour isolate. The most significant binding was observed for C. sakazakii NCTC 8155. LF has been shown to display anti-bacterial [50,51] and anti-infective [52] activity against C. sakazakii. This study confirms the direct binding of LF to C. sakazakii, which may give further insight into its mode of antibacterial action, which to date is believed to be dependent on the protein’s iron chelating ability. Galactoligosaccharides [53] and LF-containing bovine whey powders [54] have previously been shown to inhibit the adherence of Cronobacter strains to gastrointestinal cells and in some cases prevent invasion. For both strains, a higher RU value was noted when the immobilised analyte was the mature milk glycovariant, again highlighting the importance of the mature milk glycoprofile in bacterial binding. In relation to the strain specificity observed for C. sakazakii, LF binds more intensely to the strain isolated from milk powder. Milk powder is a common source of C. sakazakii contamination and the World Health Organisation (WHO) has issued guidelines relating to the recommended minimum temperature for reconstitution to reduce infection risk. Fortification of milk powders and infant milk formulas with LF as a secondary hurdle to infection may further reduce the annual incidences of C. sakazakii infection, which are most common in neonates, infants and immunocompromised adults.
Novel interactions for S. pneumoniae ATCC BAA-255 and P. aeruginosa ATCC 33354 with LF were observed in this study, both interacting to a greater extent with the glycoprotein from mature milk. Two proteins from S. pneumoniae which bind to human LF have previously been identified [55]. These proteins have been suggested to be elements of a novel virulence mechanism, harnessing LF’s iron binding activity to overcome iron limitation at mucosal surfaces. Antibacterial activity against P. aeruginosa has been observed upon exposure to a peptide derived from LF, lactoferricin B [56]. However, this is the first study, to the best of our knowledge, that demonstrates binding of bovine milk derived LF to the whole bacterial cells. Both S. pneumoniae [57] and P. aeruginosa [58] have previously been shown to have affinity for neutral oligosaccharides with terminal D-Gal residues, suggesting that the complex type glycans in mature milk have a role to play in the binding of these strains. Complex glycans have a higher level of sialylation in colostrum [7], inhibiting recognition of terminal Gal residues by bacterial lectins. Therefore, the mature milk glycoprofile offers terminal D-Gal residues as targets for bacterial binding.
S. typhimurium was the sole bacterial strain to show greater binding to colostrum LF. S. typhimurium adhesion to intestinal cells has previously been shown to be linked to cellular glycosylation and can be inhibited by lectins PNA, AIA, ECA, RCA I and WGA which compete for the glycan receptors including Galβ(1–3)GalNAc on the epithelial cell surface [59]. Binding patterns of these lectins to LF over lactation was previously profiled and all lectins displayed the highest interaction with colostrum LF [7], suggesting the glycosylation profile of LF from early lactation is in some way homologous to the binding sites on Caco-2 cells for S. typhimurium. Although cows of all ages can be infected with Salmonella bacteria, serious infections and deaths are most often seen in young calves [60]. The glycosylation pattern of LF in early lactation may contribute to the immune protection delivered to the calf via its mother’s milk and may have evolved to offer protection against infections most likely to occur in early life.
Colostrum LF has a higher N-acetylneuraminic acid (Neu5Ac) content compared to mature milk LF [7] and this monosaccharide is closely linked to LF’s antibacterial activity, as previously reviewed [6]. However, minimal bacterial binding to colostrum LF was observed in this study. This suggests that bovine LF has two modes of action for its anti-microbial activity; transitioning from a Neu5Ac dependent functionality in early lactation, to a more non-specific role in mature milk as a decoy receptor, which can bind a number of pathogenic bacteria, preventing adhesion and infection. This would suggest LF’s glycosylation changes over the lactation period to deliver the most optimum protection relevant to the life-stage of the offspring.
Acknowledgments
Noelle O’Riordan is in receipt of a Teagasc Walsh Fellowship. The authors would also like to thank Dr. Jonathan Lane for helpful advice.
Author Contributions
Rita Hickey and Lokesh Joshi conceived the study design; Noelle O’Riordan performed the experiments and analyzed the data; Michelle Kilcoyne contributed to data analysis. All authors contributed to writing the paper.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Kumar, J.; Weber, W.; Munchau, S.; Yadav, S.; Singh, S.B.; Saravanan, K.; Paramasivam, M.; Sharma, S.; Kaur, P.; Bhushan, A.; et al. Crystal structure of human seminal diferric lactoferrin at 3.4 angstrom resolution. Indian J. Biochem. Biophys. 2003, 40, 14–21. [Google Scholar] [PubMed]
- Severin, S.; Wenshui, X. Milk biologically active components as nutraceuticals: Review. Crit Rev. Food Sci. Nutr. 2005, 45, 645–656. [Google Scholar] [CrossRef] [PubMed]
- Wei, Z.; Nishimura, T.; Yoshida, S. Presence of a glycan at a potential N-glycosylation site, Asn-281, of bovine lactoferrin. J. Dairy Sci. 2000, 83, 683–689. [Google Scholar] [CrossRef]
- Baker, E.N.; Baker, H.M. Molecular structure, binding properties and dynamics of lactoferrin. Cell. Mol. Life Sci. 2005, 62, 2531–2539. [Google Scholar] [CrossRef] [PubMed]
- Pierce, A.; Colavizza, D.; Benaissa, M.; Maes, P.; Tartar, A.; Montreuil, J.; Spik, G. Molecular cloning and sequence analysis of bovine lactotransferrin. Eur. J. Biochem. 1991, 196, 177–184. [Google Scholar] [CrossRef] [PubMed]
- O'Riordan, N.; Kane, M.; Joshi, L.; Hickey, R.M. Structural and functional characteristics of bovine milk protein glycosylation. Glycobiology 2014, 24, 220–236. [Google Scholar] [CrossRef] [PubMed]
- O'Riordan, N.; Gerlach, J.Q.; Kilcoyne, M.; O’Callaghan, J.; Kane, M.; Hickey, R.M.; Joshi, L. Profiling temporal changes in bovine milk lactoferrin glycosylation using lectin microarrays. Food Chem. 2014, 165, 388–396. [Google Scholar] [CrossRef] [PubMed]
- Van Leeuwen, S.S.; Schoemaker, R.J.W.; Timmer, C.J.; Kamerling, J.P.; Dijkhuizen, L. Use of Wisteria floribunda agglutinin affinity chromatography in the structural analysis of the bovine lactoferrin N-linked glycosylation. Biochim. Biophys. Acta 2012, 1820, 1444–1455. [Google Scholar] [CrossRef] [PubMed]
- Gonzalez-Chavez, S.A.; Arevalo-Gallegos, S.; Rascon-Cruz, Q. Lactoferrin: Structure, function and applications. Int. J. Antimicrob. Agents 2009, 33, 301.e1–301.e8. [Google Scholar] [CrossRef] [PubMed]
- Debbabi, H.; Dubarry, M.; Rautureau, M.; Tome, D. Bovine lactoferrin induces both mucosal and systemic immune response in mice. J. Dairy Res. 1998, 65, 283–293. [Google Scholar] [CrossRef] [PubMed]
- Kim, W.S.; Ohashi, M.; Tanaka, T.; Kumura, H.; Kim, G.Y.; Kwon, I.K.; Goh, J.S.; Shimazaki, K. Growth-promoting effects of lactoferrin on L. acidophilus and Bifidobacterium spp. Biometals 2004, 17, 279–283. [Google Scholar] [CrossRef] [PubMed]
- Rahman, M.M.; Kim, W.S.; Ito, T.; Kumura, H.; Shimazaki, K. Growth promotion and cell binding ability of bovine lactoferrin to Bifidobacterium longum. Anaerobe 2009, 15, 133–137. [Google Scholar] [CrossRef] [PubMed]
- Cornish, J.; Callon, K.E.; Naot, D.; Palmano, K.P.; Banovic, T.; Bava, U.; Watson, M.; Lin, J.M.; Tong, P.C.; Chen, Q.; et al. Lactoferrin is a potent regulator of bone cell activity and increases bone formation in vivo. Endocrinology 2004, 145, 4366–4374. [Google Scholar] [CrossRef] [PubMed]
- Tsuda, H.; Kozu, T.; Iinuma, G.; Ohashi, Y.; Saito, Y.; Saito, D.; Akasu, T.; Alexander, D.B.; Futakuchi, M.; Fukamachi, K.; et al. Cancer prevention by bovine lactoferrin: From animal studies to human trial. Biometals 2010, 23, 399–409. [Google Scholar] [CrossRef] [PubMed]
- Tsuda, H.; Sekine, K.; Fujita, K.; Ligo, M. Cancer prevention by bovine lactoferrin and underlying mechanisms a review of experimental and clinical studies. Biochem. Cell Biol. 2002, 80, 131–136. [Google Scholar] [CrossRef] [PubMed]
- Adlerova, L.; Bartoskova, A.; Faldyna, M. Lactoferrin: A review. Vet. Med. 2008, 53, 457–468. [Google Scholar]
- Garcia-Montoya, I.A.; Cendon, T.S.; Arevalo-Gallegos, S.; Rascon-Cruz, Q. Lactoferrin a multiple bioactive protein: An overview. Biochim. Biophys. Acta 2012, 1820, 226–236. [Google Scholar] [CrossRef] [PubMed]
- Rossi, P.; Giansanti, F.; Boffi, A.; Ajello, M.; Valenti, P.; Chiancone, E.; Antonini, G. Ca2+ binding to bovine lactoferrin enhances protein stability and influences the release of bacterial lipopolysaccharide. Biochem. Cell Biol. 2002, 80, 41–48. [Google Scholar] [CrossRef] [PubMed]
- Legrand, D.; Mazurier, J.; Colavizza, D.; Montreuil, J.; Spik, G. Properties of the iron-binding site of the N-terminal lobe of human and bovine lactotransferrins—Importance of the glycan moiety and of the noncovalent interactions between the N-terminal and C-terminal lobes in the stability of the iron-binding site. Biochem. J. 1990, 266, 575–581. [Google Scholar] [PubMed]
- Moore, S.A.; Anderson, B.F.; Groom, C.R.; Haridas, M.; Baker, E.N. Three-dimensional structure of diferric bovine lactoferrin at 2.8 Å resolution. J. Mol. Biol. 1997, 274, 222–236. [Google Scholar] [CrossRef] [PubMed]
- Van Veen, H.A.; Geerts, M.E.; van Berkel, P.H.; Nuijens, J.H. The role of N-linked glycosylation in the protection of human and bovine lactoferrin against tryptic proteolysis. Eur. J. Biochem. 2004, 271, 678–684. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Hirmo, S.; Willen, R.; Wadstrom, T. Inhibition of Helicobacter pylori infection by bovine milk glycoconjugates in a BALB/cA mouse model. J. Med. Microbiol. 2001, 50, 430–435. [Google Scholar] [CrossRef] [PubMed]
- Teraguchi, S.; Shin, K.; Fukuwatari, Y.; Shimamura, S. Glycans of bovine lactoferrin function as receptors for the type 1 fimbrial lectin of Escherichia coli. Infect. Immun. 1996, 64, 1075–1077. [Google Scholar] [PubMed]
- Yoshida, S.; Wei, Z.; Shinmura, Y.; Fukunaga, N. Separation of lactoferrin-a and -b from bovine colostrum. J. Dairy Sci. 2000, 83, 2211–2215. [Google Scholar] [CrossRef]
- Kawasaki, Y.; Isoda, H.; Shinmoto, H.; Tanimoto, M.; Dosako, S.; Idota, T.; Nakajima, I. Inhibition by k-casein glycomacropeptide and lactoferrin of influenza virus hemmaglutination. Biosci. Biotechnol. Biochem. 1993, 57, 1214–1215. [Google Scholar] [CrossRef] [PubMed]
- Kawasaki, Y.; Isoda, H.; Tanimoto, M.; Dosako, S.; Idota, T.; Ahiko, K. Inhibition by lactoferrin and kappa-casein glycomacropeptide of binding of cholera-toxin to its receptor. Biosci. Biotechnol. Biochem. 1992, 56, 195–198. [Google Scholar] [CrossRef] [PubMed]
- Huber, W.; Mueller, F. Biomolecular interaction analysis in drug discovery using surface plasmon resonance technology. Curr. Pharm. Des. 2006, 12, 3999–4021. [Google Scholar] [CrossRef] [PubMed]
- Lane, J.A.; Mehra, R.K.; Carrington, S.D.; Hickey, R.M. Development of biosensor-based assays to identify anti-infective oligosaccharides. Anal. Biochem. 2011, 410, 200–205. [Google Scholar] [CrossRef] [PubMed]
- Lin, P.-H.; Chen, R.-H.; Lee, C.-H.; Chang, Y.; Chen, C.-S.; Chen, W.-Y. Studies of the binding mechanism between aptamers and thrombin by circular dichroism, surface plasmon resonance and isothermal titration calorimetry. Colloids Surf. B 2011, 88, 552–558. [Google Scholar] [CrossRef] [PubMed]
- Šípová, H.; Zhang, S.; Dudley, A.M.; Galas, D.; Wang, K.; Homola, J. Surface plasmon resonance biosensor for rapid label-free detection of microribonucleic acid at subfemtomole level. Anal. Chem. 2010, 82, 10110–10115. [Google Scholar] [CrossRef] [PubMed]
- Milkani, E.; Lambert, C.R.; McGimpsey, W.G. Direct detection of acetylcholinesterase inhibitor binding with an enzyme-based surface plasmon resonance sensor. Anal. Biochem. 2011, 408, 212–219. [Google Scholar] [CrossRef] [PubMed]
- Schlick, K.H.; Cloninger, M.J. Inhibition binding studies of glycodendrimer/lectin interactions using surface plasmon resonance. Tetrahedron 2010, 66, 5305–5310. [Google Scholar] [CrossRef] [PubMed]
- Nilsson, C.E.; Abbas, S.; Bennemo, M.; Larsson, A.; Hämäläinen, M.D.; Frostell-Karlsson, Å. A novel assay for influenza virus quantification using surface plasmon resonance. Vaccine 2010, 28, 759–766. [Google Scholar] [CrossRef] [PubMed]
- Morrow, A.L.; Ruiz-Palacios, G.M.; Altaye, M.; Jiang, X.; Guerrero, M.L.; Meinzen-Derr, J.K.; Farkas, T.; Chaturvedi, P.; Pickering, L.K.; Newburg, D.S. Human milk oligosaccharide blood group epitopes and innate immune protection against Campylobacter and calicivirus diarrhea in breastfed infants. In Protecting Infants through Human Milk; Pickering, L., Morrow, A., Ruiz-Palacios, G., Schanler, R., Eds.; Kluwer academic/Plenum publishers: New York, NY, USA, 2004; Volume 554, pp. 443–446. [Google Scholar]
- Newburg, D.S.; Ruiz-Palacios, G.M.; Morrow, A.L. Human milk glycans protect infants against enteric pathogens. Annu. Rev. Nutr. 2005, 25, 37–58. [Google Scholar] [CrossRef] [PubMed]
- Huang, I.N.; Okawara, T.; Watanabe, M.; Kawai, Y.; Kitazawa, H.; Ohnuma, S.; Shibata, C.; Horii, A.; Kimura, K.; Taketomo, N.; et al. New screening methods for probiotics with adhesion properties to sialic acid and sulphate residues in human colonic mucin using the biacore assay. J. Appl. Microbiol. 2013, 114, 854–860. [Google Scholar] [CrossRef] [PubMed]
- Kinoshita, H.; Uchida, H.; Kawai, Y.; Kitazawa, H.; Miura, K.; Shiiba, K.; Horii, A.; Saito, T. Quantitative evaluation of adhesion of lactobacilli isolated from human intestinal tissues to human colonic mucin using surface plasmon resonance (biacore assay). J. Appl. Microbiol. 2007, 102, 116–123. [Google Scholar] [CrossRef] [PubMed]
- Holmes, S.D.; May, K.; Johansson, V.; Markey, F.; Critchley, I.A. Studies on the interaction of Staphylococcus aureus and Staphylococcus epidermidis with fibronectin using surface plasmon resonance (biacore). J. Microbiol. Methods 1997, 28, 77–84. [Google Scholar] [CrossRef]
- Oli, M.W.; McArthur, W.P.; Brady, L.J. A whole cell BIAcore assay to evaluate P1-mediated adherence of Streptococcus mutans to human salivary agglutinin and inhibition by specific antibodies. J. Microbiol. Methods 2006, 65, 503–511. [Google Scholar] [CrossRef] [PubMed]
- Bulard, E.; Bouchet-Spinelli, A.; Chaud, P.; Roget, A.; Calemczuk, R.; Fort, S.; Livache, T. Carbohydrates as new probes for the identification of closely related Escherichia coli strains using surface plasmon resonance imaging. Anal. Chem. 2015, 87, 1804–1811. [Google Scholar] [CrossRef] [PubMed]
- Naidu, A.S.; Andersson, M.; Miedzobrodzki, J.; Forsgren, A.; Watts, J.L. Bovine lactoferrin receptors in Staphylococcus aureus isolated from bovine mastitis. J. Dairy Sci. 1991, 74, 1218–1226. [Google Scholar] [CrossRef]
- Roe, A.J.; Currie, C.; Smith, D.G.; Gally, D.L. Analysis of type 1 fimbriae expression in verotoxigenic Escherichia coli: A comparison between serotypes O157 and O26. Microbiology 2001, 147, 145–152. [Google Scholar] [CrossRef] [PubMed]
- Abraham, S.N.; Sun, D.X.; Dale, J.B.; Beachey, E.H. Conservation of the D-mannose-adhesion protein among type-1 fimbriated members of the family enterobacteriaceae. Nature 1988, 336, 682–684. [Google Scholar] [CrossRef] [PubMed]
- Jaques, L.W.; Brown, E.B.; Barrett, J.M.; Brey, W.S., Jr.; Weltner, W., Jr. Sialic acid. A calcium-binding carbohydrate. J. Biol. Chem. 1977, 252, 4533–4538. [Google Scholar] [PubMed]
- O’Flaherty, S.; Flynn, J.; Coffey, A.; Fitzgerald, G.; Meaney, B.; Ross, P. Molecular Characterisation of Bacteriophage K towards Applications for the Biocontrol of Pathogenic Staphylococci. Ph.D. Thesis, University College, Cork, Ireland, 2005. [Google Scholar]
- Hagiwara, S.; Kawai, K.; Anri, A.; Nagahata, H. Lactoferrin concentrations in milk from normal and subclinical mastitic cows. J. Vet. Med. Sci. 2003, 65, 319–323. [Google Scholar] [CrossRef] [PubMed]
- Szunerits, S.; Zagorodko, O.; Cogez, V.; Dumych, T.; Chalopin, T.; Alvarez Dorta, D.; Sivignon, A.; Barnich, N.; Harduin-Lepers, A.; Larroulet, I.; et al. Differentiation of crohn’s disease-associated isolates from other pathogenic Escherichia coli by fimbrial adhesion under shear force. Biology 2016, 5, 14. [Google Scholar] [CrossRef] [PubMed]
- Erdei, J.; Forsgren, A.; Naidu, A.S. Lactoferrin binds to porins OmpF and OmpC in Escherichia coli. Infect. Immun. 1994, 62, 1236–1240. [Google Scholar] [PubMed]
- Naidu, S.S.; Erdei, J.; Czirok, E.; Kalfas, S.; Gado, I.; Thoren, A.; Forsgren, A.; Naidu, A.S. Specific binding of lactoferrin to Escherichia coli isolated from human intestinal infections. APMIS 1991, 99, 1142–1150. [Google Scholar] [CrossRef] [PubMed]
- Wakabayashi, H.; Yamauchi, K.; Takase, M. Inhibitory effects of bovine lactoferrin and lactoferricin b on Enterobacter sakazakii. Biocontrol Sci. 2008, 13, 29–32. [Google Scholar] [CrossRef] [PubMed]
- Harouna, S.; Carramiñana, J.J.; Navarro, F.; Pérez, M.D.; Calvo, M.; Sánchez, L. Antibacterial activity of bovine milk lactoferrin on the emerging foodborne pathogen Cronobacter sakazakii: Effect of media and heat treatment. Food Control 2015, 47, 520–525. [Google Scholar] [CrossRef]
- Quintero-Villegas, M.; Wittke, A.; Hutkins, R. Adherence inhibition of Cronobacter sakazakii to intestinal epithelial cells by lactoferrin. Curr. Microbiol. 2014, 69, 574–579. [Google Scholar] [CrossRef] [PubMed]
- Quintero, M.; Maldonado, M.; Perez-Munoz, M.; Jimenez, R.; Fangman, T.; Rupnow, J.; Wittke, A.; Russell, M.; Hutkins, R. Adherence inhibition of Cronobacter sakazakii to intestinal epithelial cells by prebiotic oligosaccharides. Curr. Microbiol. 2011, 62, 1448–1454. [Google Scholar] [CrossRef] [PubMed]
- Halpin, R.; Brady, D.; O’Riordan, E.; O’Sullivan, M. Untreated and enzyme-modified bovine whey products reduce association of Salmonella typhimurium, Escherichia coli O157: H7 and Cronobacter malonaticus (formerly Enterobacter sakazakii) to Caco-2 cells. J. Appl. Microbiol. 2010, 108, 406–415. [Google Scholar] [CrossRef] [PubMed]
- Hammerschmidt, S.; Bethe, G.; Remane, P.H.; Chhatwal, G.S. Identification of pneumococcal surface protein A as a lactoferrin-binding protein of Streptococcus pneumoniae. Infect. Immun. 1999, 67, 1683–1687. [Google Scholar] [PubMed]
- Bellamy, W.; Takase, M.; Wakabayashi, H.; Kawase, K.; Tomita, M. Antibacterial spectrum of lactoferricin b, a potent bactericidal peptide derived from the N-terminal region of bovine lactoferrin. J. Appl. Bacteriol. 1992, 73, 472–479. [Google Scholar] [CrossRef] [PubMed]
- Andersson, B.; Dahmen, J.; Frejd, T.; Leffler, H.; Magnusson, G.; Noori, G.; Eden, C.S. Identification of an active disaccharide unit of a glycoconjugate receptor for pneumococci attaching to human pharyngeal epithelial cells. J. Exp. Med. 1983, 158, 559–570. [Google Scholar] [CrossRef] [PubMed]
- Lesman-Movshovich, E.; Lerrer, B.; Gilboa-Garber, N. Blocking of Pseudomonas aeruginosa lectins by human milk glycans. Can. J. Microbiol. 2003, 49, 230–235. [Google Scholar] [CrossRef] [PubMed]
- Giannasca, K.T.; Giannasca, P.J.; Neutra, M.R. Adherence of Salmonella typhimurium to Caco-2 cells: Identification of a glycoconjugate receptor. Infect. Immun. 1996, 64, 135–145. [Google Scholar] [PubMed]
- Robertsson, J.A.; Lindberg, A.A.; Hoiseth, S.; Stocker, B.A. Salmonella typhimurium infection in calves: Protection and survival of virulent challenge bacteria after immunization with live or inactivated vaccines. Infect. Immun. 1983, 41, 742–750. [Google Scholar] [PubMed]
Figure 1.
Increasing concentrations of Staphylococcus aureus DPC 5971 (red) and Escherichia coli O157:H7 P1432 (blue) injected over the surface of a streptavidin (SA) chip with mature LF immobilised to evaluate effect of increasing bacterial numbers on the response units (RU) response.
Figure 1.
Increasing concentrations of Staphylococcus aureus DPC 5971 (red) and Escherichia coli O157:H7 P1432 (blue) injected over the surface of a streptavidin (SA) chip with mature LF immobilised to evaluate effect of increasing bacterial numbers on the response units (RU) response.

Figure 2.
RU changes following exposure of a panel of pathogenic bacteria to LF from colostrum (white) or mature milk (grey) immobilised on a Biacore SA chip. E. coli P1432 was identified as a negative control and its response was marked as the minimum requirement for consideration as a positive interaction (black horizontal line). All bacteria were applied at a concentration of 1 × 108 CFU·mL−1.
Figure 2.
RU changes following exposure of a panel of pathogenic bacteria to LF from colostrum (white) or mature milk (grey) immobilised on a Biacore SA chip. E. coli P1432 was identified as a negative control and its response was marked as the minimum requirement for consideration as a positive interaction (black horizontal line). All bacteria were applied at a concentration of 1 × 108 CFU·mL−1.

{kind=link}
{kind=link}
Table 1.
List of bacterial strains tested.
| Strain | Media |
|---|---|
| Staphylococcus aureus DPC 5971 | Muller-Hinton |
| Escherichia coli O157:H7 P1432 | Muller-Hinton |
| Escherichia coli O157:H7 NCTC 12900 | Muller-Hinton |
| Cronobacter sakazakii NCTC 8155 | Brain Heart Infusion |
| Cronobacter sakazakii DPC 6531 | Brain Heart Infusion |
| Streptococcus pneumoniae ATCC BAA-255 | Todd Hewitt + 0.5% yeast extract |
| Pseudomonas aeruginosa ATCC 33354 | Tryptic soy broth |
| Salmonella enterica subsp. enterica serovar Typhimurium ATCC BAA-185 | Brain Heart Infusion |
| Listeria monocytogenes DPC 3437 | Brain Heart Infusion |
| Listeria monocytogenes NCTC 11994 | Brain Heart Infusion |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
O’Riordan, N.; Kilcoyne, M.; Joshi, L.; Hickey, R.M. Exploitation of SPR to Investigate the Importance of Glycan Chains in the Interaction between Lactoferrin and Bacteria. Sensors 2017, 17, 1515. https://doi.org/10.3390/s17071515
AMA Style
O’Riordan N, Kilcoyne M, Joshi L, Hickey RM. Exploitation of SPR to Investigate the Importance of Glycan Chains in the Interaction between Lactoferrin and Bacteria. Sensors. 2017; 17(7):1515. https://doi.org/10.3390/s17071515
Chicago/Turabian StyleO’Riordan, Noelle, Michelle Kilcoyne, Lokesh Joshi, and Rita M. Hickey. 2017. "Exploitation of SPR to Investigate the Importance of Glycan Chains in the Interaction between Lactoferrin and Bacteria" Sensors 17, no. 7: 1515. https://doi.org/10.3390/s17071515
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.