Fucoidans as Potential Inhibitors of HIV-1

 ,
,
Abstract
:1. Introduction
2. Results and Discussion
2.1. Characterization of the Fucoidans from Brown Algae

{kind=link}
{kind=link}
{kind=link}
| Seaweed source | Fraction | Mw, kDa | The linkage between Fuc residues | SO3Na− (%) * | Monosaccharide composition, mol% |
|---|---|---|---|---|---|
| Fuc/Gal/Man/Rha/Xyl/Glc *** | |||||
| α-l-Fucans (F) | |||||
| S. cichorioides | ScF | 1160 | α-1,3 | 26.5 | 88.6/6.4/3.1/0/1.9/0 |
| F. evanescens | FeF ** | 620 | α-1,3;1,4 | 23.2 | 77.9/9.5/4.2/0/8.4/0 |
| F. evanescens | FeFDA | 20 | α-1,3;1,4 | 21.3 | 82.7/8.7/3.0/0/5.6/0 |
| Galactofucans (GF) | |||||
| S. gurjanovae | SgGF ** | 810 | α-1,3 | 28.2 | 64.3/20.7/14.6/0.4/0/0 |
| Al. ochotensis | AoGF | 860 | α-1,3 | 24.0 | 53.8/38.5/7.7/0/0/0 |
| S. japonica | SjGF ** | 1800 | α-1,3;1,4 | 23.3 | 49.9/44.1/5.3/0/1.1/0 |
| C. costata | CcGF ** | 160 | α-1,3 | 23.2 | 62.6/30.1/2.6/1.7/0/0 |
2.2. Antiviral Activity
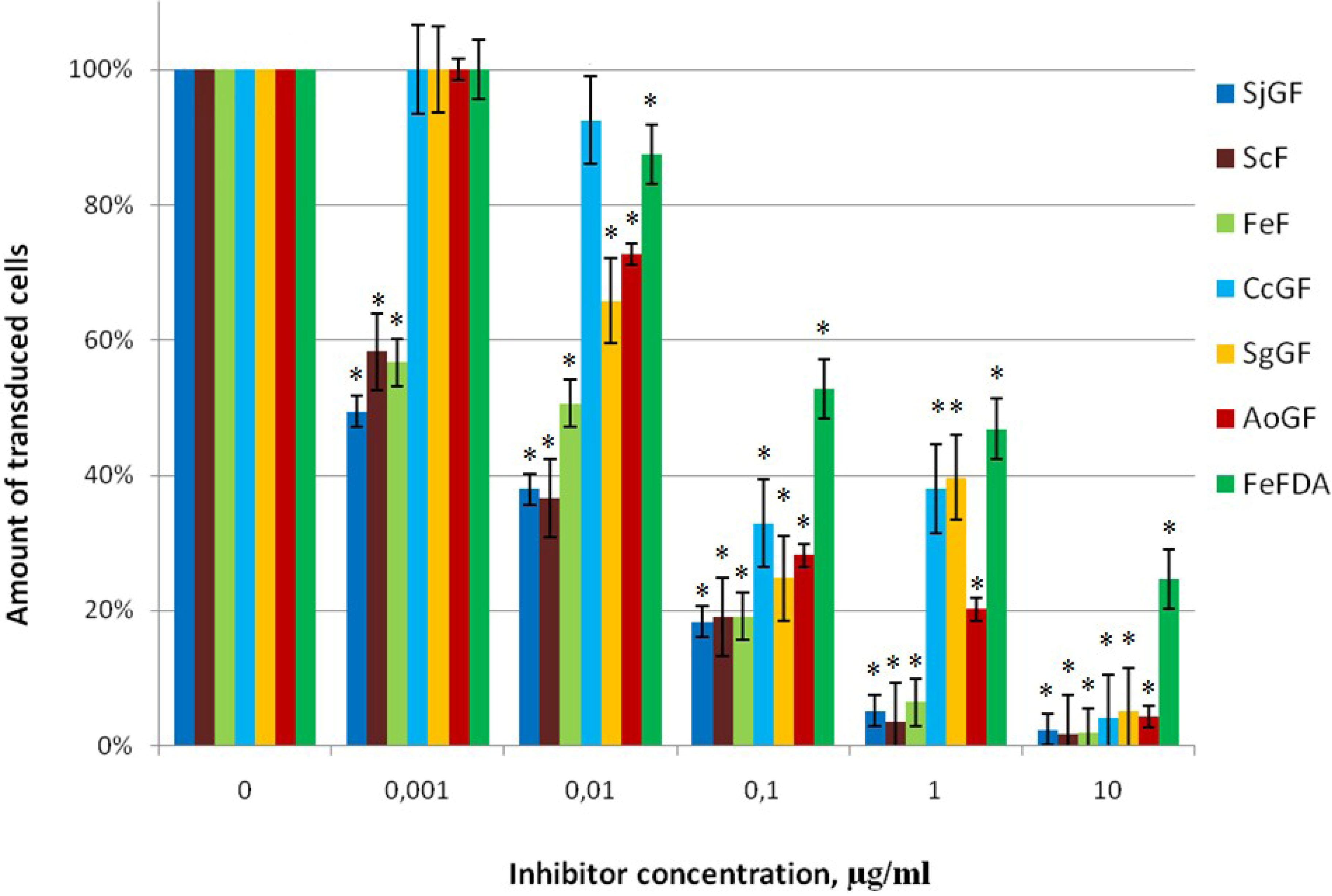
2.2.2. Inhibitor Activity of the Fucoidans Against Transduction of Jurkat Cells by Pseudo-HIV-1 Particles that Contain HIV-1 gp120+gp41 Envelope Protein

| Seaweed source | Fraction | Mw, kDa | IC50 (Pseudo-HIV-1 particles containing envelope protein gp120+gp41 HIV-1) µg/mL * | IC50 (Pseudo-HIV-1 particles containing envelope protein VSV-G) µg/mL * | IC50 (Mo-MuLV) µg/mL ** |
|---|---|---|---|---|---|
| α-l-Fucans (F) | |||||
| S. cichorioides | ScF | 1160 | 0.0050 ± 0.0003 | >100 | 0.0060 ± 0.0003 |
| F. evanescens | FeF | 620 | 0.0100 ± 0.0004 | >100 | 0.0060 ± 0.0003 |
| F. evanescens | FeFDA | 20 | 0.52 ± 0.02 | n.d. | 4.50 ± 0.14 |
| Galactofucans (GF) | |||||
| S. gurjanovae | SgGF | 810 | 0.0450 ± 0.003 | n.d. | 0.250 ± 0.009 |
| Al. ochotensis | AoGF | 860 | 0.0550 ± 0.0009 | n.d. | 0.0540 ± 0.0025 |
| S. japonica | SjGF | 1800 | 0.00100 ± 0.00003 | >100 | 0.0050 ± 0.0002 |
| C. costata | CcGF | 160 | 0.075 ± 0.005 | n.d. | 0.066 ± 0.003 |
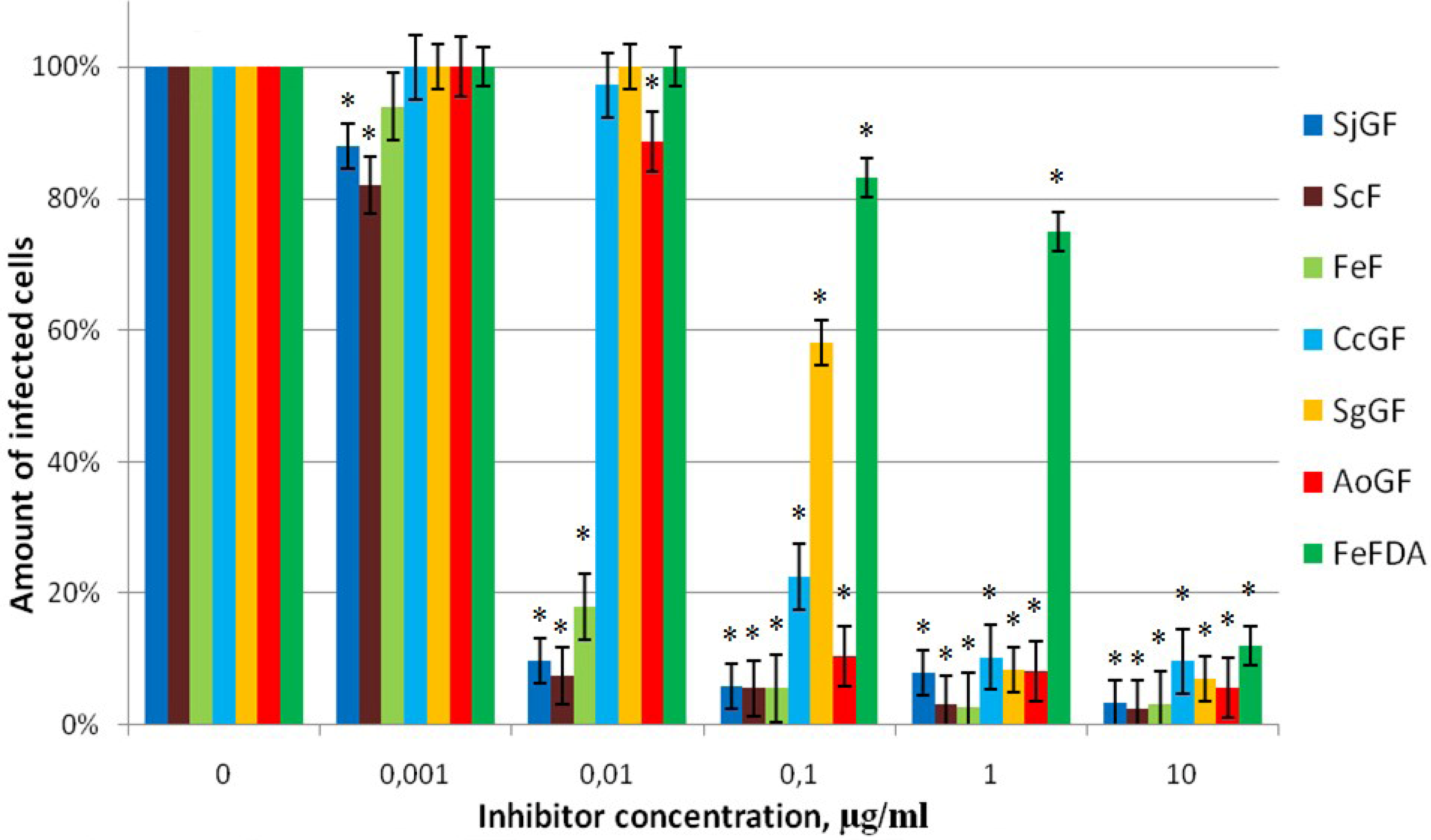
2.2.3. Inhibitor Activity of the Fucoidans Against Transduction of Jurkat Cells by Pseudo-HIV-1 Particles that Contain G Envelope Protein from VSV

2.2.4. Inhibitor Activity of the Fucoidans Against Mo-MuLV Infection of SC-1 Cells

3. Experimental Section
3.1. Biological Material
3.2. Isolation and Determination of the Fucoidan Structure
3.2.1. Extraction of Water-Soluble Polysaccharide from the Brown Algae
3.2.2. Analysis of Monosaccharide Composition
3.2.3. Deacetylating of the Fucoidan
3.2.4. Sulfate Group Content
3.2.5. Anion-Exchange Chromatography
3.2.6. Determination of Molecular Weight
3.2.7. NMR Spectroscopy
3.3. Biological Assays
3.3.1. Cell Culture
3.3.2. Generation of Jurkat Clone 4X4#3
Antibodies and FACS Analysis
3.3.3. Obtaining of Pseudo-HIV-1 Particles
3.3.4. Obtaining Replication Competent Virus Particles
3.3.5. Cytotoxicity Assay
4. Conclusions
Acknowledgments
Conflicts of Interest
References
- Rabenstein, D.L. Heparin and heparan sulfate: Structure and function. Nat. Prod. Rep. 2002, 19, 312–331. [Google Scholar] [CrossRef]
- O’Donnell, C.D.; Shukla, D. The importance of heparan sulfate in herpesvirus infection. Virol. Sin. 2008, 23, 383–393. [Google Scholar] [CrossRef]
- Vermeire, K.; Schols, D.; Bell, T.W. Inhibitors of HIV infection via the cellular CD4 receptor. Curr. Med. Chem. 2006, 13, 731–743. [Google Scholar] [CrossRef]
- Baba, M.; Snoeck, R.; Pauwels, R.; de Clercq, E. Sulfated polysaccharides are potent and selective inhibitors of various enveloped viruses, including herpes simplex virus, cytomegalovirus, vesicular stomatitis virus, and human immunodeficiency virus. Antimicrob. Agents Chemother. 1988, 32, 1742–1745. [Google Scholar] [CrossRef]
- Ghosh, T.; Chattopadhyay, K.; Marschall, M.; Karmakar, P.; Mandal, P.; Ray, B. Focus on antivirally active sulfated polysaccharides: From structure-activity analysis to clinical evaluation. Glycobiology 2009, 19, 2–15. [Google Scholar]
- Liu, H.; Geng, M.; Xin, X.; Li, F.; Zhang, Z.; Li, J.; Ding, J. Multiple and multivalent interactions of novel anti-AIDS drug candidates, sulfated polymannuronate (SPMG)-derived oligosaccharides, with gp120 and their anti-HIV activities. Glycobiology 2005, 15, 501–510. [Google Scholar]
- Damonte, E.B.; Matulewicz, M.C.; Cerezo, A.S. Sulfated seaweed polysaccharides as antiviral agents. Curr. Med. Chem. 2004, 11, 2399–2419. [Google Scholar] [CrossRef]
- Witvrouw, M.; de Clercq, E. Sulfated polysaccharides extracted from sea algae as potential antiviral drugs. Gen. Pharmacol. 1997, 29, 497–511. [Google Scholar] [CrossRef]
- Bilan, M.I.; Usov, A.I. Structural analysis of fucoidans. Nat. Prod. Commun. 2008, 3, 1639–1648. [Google Scholar]
- Kusaykin, M.I.; Yu Bakunina, I.; Sova, V.V.; Ermakova, S.P.; Kuznetsova, T.S.; Besednova, N.N.; Zaporozhets, T.S.; Zvyagintseva, T.N. Enzymatic transformation of brown seaweed polyanionic polysaccharides. Structure and biological action of the polysaccharides and products of their transformation. J. Biotechnol. 2008, 3, 904–915. [Google Scholar] [CrossRef]
- Bilan, M.I.; Grachev, A.A.; Shashkov, A.S.; Kelly, M.; Sanderson, C.J.; Nifantiev, N.E.; Usov, A.I. Further studies on the composition and structure of a fucoidan preparation from the brown alga Saccharina latissima. Carbohydr. Res. 2010, 345, 2038–2047. [Google Scholar] [CrossRef]
- Imbs, T.I.; Shevchenko, N.M.; Semenova, T.L.; Sukhoverkhov, S.V.; Zvyagintseva, T.N. Compositional heterogeneity of sulfated polysaccharides synthesized by the brown alga Costaria costata. Chem. Nat. Compd. 2011, 47, 96–97. [Google Scholar] [CrossRef]
- Queiroz, K.C.S.; Medeiros, V.P.; Queiroz, L.S.; Abreu, L.R.D.; Pocha, H.A.O.; Ferreira, C.V.; Juca, M.B.; Aoyama, H.; Leite, E.L. Inhibition of reverse transcriptase activity of HIV by polysaccharides of brown algae. Biomed. Pharmacother. 2008, 62, 303–307. [Google Scholar] [CrossRef]
- De Clercq, E. Anti-HIV drugs: 25 compounds approved within 25 years after the discovery of HIV. Int. J. Antimicrob. Agents 2009, 33, 307–320. [Google Scholar] [CrossRef]
- Stepanov, O.A.; Prokofjeva, M.M.; Stocking, K.; Varlamov, V.P.; Levov, A.N.; Vikhoreva, G.A.; Spirin, P.V.; Mikhailov, S.N.; Prassolov, V.S. Replication-competent gamma-retrovirus Mo-MuLV expressing green fluorescent protein as efficient tool for screening of inhibitors of retroviruses that use heparin sulfate as primary cell receptor. Mol. Biol. 2012, 46, 457–466. [Google Scholar] [CrossRef]
- Prokofjeva, M.M.; Riecken, K.; Spirin, P.V.; Yanvarév, D.V.; Düsedau, A.; Ellinger, B.; Fehse, B.; Stocking, C.; Prassolov, V.S. A new system for parallel drug screening against multiple-resistant HIV mutants based on lentiviral self-inactivating (SIN) vectors and multi-colour analyses. AIDS Res. Ther. 2013, 10, 1. [Google Scholar] [CrossRef]
- Khil’chenko, S.R.; Zaporozhets, T.S.; Shevchenko, N.M.; Zvyagintseva, T.N.; Vogel, U.; Seeberger, P.H.; Lepenies, B. Immunostimulatory activity of fucoidan from the brown alga Fucus evanescens: Role of sulfates and acetates. J. Carbohydr. Chem. 2011, 30, 291–305. [Google Scholar]
- Shevchenko, N.M.; Anastyuk, S.D.; Gerasimenko, N.I.; Dmitrenok, P.S.; Isakov, V.V.; Zvyagintseva, T.N. Polysaccharide and lipid composition of the brown seaweed Laminaria gurjanovae. Russ. J. Bioorg. Chem. 2007, 33, 88–98. [Google Scholar] [CrossRef]
- Makarenkova, I.D.; Kompanets, G.G.; Besednova, N.N.; Slonova, R.A.; Zvyagintseva, T.N.; Shevchenko, N.M. Inhibiting effects fucoidans on Hantaan virus adsorption on model of peritoneal macrophages in vitro. (in Russian). Vopr. Virusol. 2008, 53, 12–15. [Google Scholar]
- Makarenkova, I.D.; Deryabin, P.G.; Lvov, D.K.; Zvyagintseva, T.N.; Besednova, N.N. Antiviral activity of sulfated polysaccharide from the brown algae Laminaria japonica against avian influenza A (H5N1) virus infection in the cultured cells. Probl. Virol. 2010, 55, 41–45. [Google Scholar]
- Makarenkova, I.D.; Leonova, G.N.; Maystrovkaya, O.S.; Zvyagintseva, T.N.; Imbs, T.I.; Ermakova, S.P.; Besednova, N.N. Antiviral effect of brown algae-derivate sulphated polysaccharides in case of experimental tick-borne encephalitis: Tying structure and function. (in Russian). Pac. Med. J. 2012, 1, 44–46. [Google Scholar]
- Prokofjeva, M.M.; Spirin, P.V.; Yanvarev, D.V.; Ivanov, A.V.; Novikov, M.S.; Stepanov, O.A.; Gottikh, M.B.; Kochetkov, S.N.; Fehse, B.; Stocking, C.; et al. Screening of potential HIV-1 inhibitors/replication blockers using secure lentiviral in vitro system. Acta Naturae 2011, 3, 55–65. [Google Scholar]
- Adhikari, U.; Mateu, C.G.; Chattopadhyay, K.; Pujol, C.A.; Damonte, E.B.; Ray, B. Structure and antiviral activity of sulfated fucfns from Stoechospermum marginatum. Phytochemistry 2006, 67, 2474–2482. [Google Scholar] [CrossRef]
- Chattopadhyay, K.; Mateu, C.G.; Mandal, P.; Pujol, C.A.; Damonte, E.B.; Ray, B. Galactan sulfate of Grateloupia indica: Isolation, structural features and antiviral activity. Phytochemistry 2007, 68, 2428–2435. [Google Scholar]
- Chattopadhyay, K.; Ghosh, T.; Pujol, C.A.; Carlucci, M.J.; Damonte, E.B.; Ray, B. Polysaccharides from Gracilaria corticata: Sulfation, chemical characterization and anti-HSV activities. Int. J. Biol. Macromol. 2008, 43, 346–351. [Google Scholar] [CrossRef]
- Lee, J.-B.; Hou, X.; Hayashi, T. Effect of partial desulfation and oversulfation of sodium spirulan on the potency of anti-herpetic activities. Carbohydr. Polym. 2007, 69, 651–658. [Google Scholar] [CrossRef]
- Mandal, P.; Pujol, C.A.; Carlucci, M.J.; Chattopadhyay, K.; Damonte, E.B.; Ray, B. Sulfated xylomannan of Scinaia hetai: Isolation, structural features and antiviral activity. Phytochemistry 2008, 69, 2193–2199. [Google Scholar] [CrossRef]
- Harrop, H.A.; Rider, C.C. Heparin and its derivatives bind to HIV-1 recombinant envelope glycoproteins, rather than to recombinant HIV-1 receptor, CD4. Glycobiology 1998, 8, 131–137. [Google Scholar] [CrossRef]
- Moulard, M.; Lortat-Jacob, H.; Mondor, I.; Roca, G.; Wyatt, R.; Sodroski, J.; Zhao, L.; Olson, W.; Kwong, P.D.; Sattentau, Q.J. Selective interactions of polyanions with basic surfaces on human immunodeficiency virus type 1 gp120. J. Virol. 2000, 74, 1948–1960. [Google Scholar] [CrossRef]
- Vives, R.R.; Imberty, A.; Sattentau, Q.J.; Lortat-Jacob, H.J. Heparan sulfate targets the HIV-1 envelope glycoprotein gp120 coreceptor binding site. J. Biol. Chem. 2005, 280, 21353–21357. [Google Scholar]
- Carlucci, M.J.; Scolaro, L.A.; Errea, M.I.; Matulewicz, M.C.; Damonte, E.B. Antiviral activity of natural sulfated galactans on herpes virus multiplication in cell culture. Planta Med. 1997, 63, 429–432. [Google Scholar] [CrossRef]
- Nyberg, K.; Ekblad, M.; Bergstrom, T.; Freeman, C.; Parish, C.R.; Ferro, V.; Trybala, E. The low molecular weight heparan sulfate-mimetic, PI-88, inhibits cell-to-cell spread of herpes simplex virus. Antivir. Res. 2004, 63, 15–24. [Google Scholar] [CrossRef]
- Coil, D.A.; Miller, A.D. Phosphatisylserine is not the cell surface receptor for vesicular stomatitis virus. J. Virol. 2004, 78, 10920–10926. [Google Scholar] [CrossRef]
- Kusaykin, M.I.; Chizhov, A.O.; Grachev, A.A.; Alekseeva, S.A.; Yu Bakunina, I.; Nedashkovskaya, O.I.; Sova, V.V.; Zvyagintseva, T.N. A comparative study of specificity of fucoidanases from marine microorganisms and invertebrates. J. Appl. Phycol. 2006, 18, 369–373. [Google Scholar] [CrossRef]
- Vishchuk, O.S.; Tarbeeva, D.V.; Ermakova, S.P.; Zvyagintseva, T.N. Structural characteristics and biological activity of fucoidans from the brown algae Alaria sp. and Saccharina japonica of different reproductive status. Chem. Biodivers. 2012, 9, 817–828. [Google Scholar] [CrossRef]
- Imbs, T.I.; Shevchenko, N.M.; Sukhoverkhov, S.V.; Semenova, T.L.; Skriptsova, A.V.; Zvyagintseva, T.N. Seasonal variations of the composition and structural characteristics of polysaccharides from the brown alga Costaria costata. Chem. Nat. Compd. 2009, 45, 786–791. [Google Scholar] [CrossRef]
- Anastyuk, S.D.; Shevchenko, N.M.; Nazarenko, E.L.; Imbs, T.I.; Dmitrenok, P.S.; Zvyagintseva, T.N. Structural analysis of a highly sulfated fucan from the brown alga Laminaria cichorioides by tandem MALDI and ESI mass spectrometry. Carbohydr. Res. 2010, 345, 2206–2212. [Google Scholar] [CrossRef]
- Bilan, M.I.; Grachev, A.A.; Ustuzhanina, N.E.; Shashkov, A.S.; Nifantiev, N.E.; Usov, A.I. Structure of a fucoidan from the brown seaweed Fucus evanescens C. Ag. Carbohydr. Res. 2002, 337, 719–730. [Google Scholar] [CrossRef]
- Wang, J.; Zhang, Q.; Zhang, Z.; Zhang, H.; Nju, X. Structural studies on a novel fucogalactan sulfate extracted from the brown seaweed Laminaria japonica. Int. J. Biol. Macromol. 2010, 47, 126–131. [Google Scholar] [CrossRef]
- Anastyuk, S.D.; Imbs, T.I.; Shevchenko, N.M.; Dmitrenok, P.S.; Zvyagintseva, T.N. ESIMS analysis of fucoidan preparations from Costaria costata, extracted from alga at different life-stages. Carbohydr. Polym. 2012, 90, 993–1002. [Google Scholar] [CrossRef]
- Dodgson, K.S.; Price, R.G. A note on the determination of the ester sulphate content of sulphated polysaccharides. Biochem. J. 1962, 84, 106–110. [Google Scholar]
© 2013 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Prokofjeva, M.M.; Imbs, T.I.; Shevchenko, N.M.; Spirin, P.V.; Horn, S.; Fehse, B.; Zvyagintseva, T.N.; Prassolov, V.S. Fucoidans as Potential Inhibitors of HIV-1. Mar. Drugs 2013, 11, 3000-3014. https://doi.org/10.3390/md11083000
Prokofjeva MM, Imbs TI, Shevchenko NM, Spirin PV, Horn S, Fehse B, Zvyagintseva TN, Prassolov VS. Fucoidans as Potential Inhibitors of HIV-1. Marine Drugs. 2013; 11(8):3000-3014. https://doi.org/10.3390/md11083000
Chicago/Turabian StyleProkofjeva, Maria M., Tatyana I. Imbs, Natalya M. Shevchenko, Pavel V. Spirin, Stefan Horn, Boris Fehse, Tatyana N. Zvyagintseva, and Vladimir S. Prassolov. 2013. "Fucoidans as Potential Inhibitors of HIV-1" Marine Drugs 11, no. 8: 3000-3014. https://doi.org/10.3390/md11083000