Polyunsaturated Aldehydes from Large Phytoplankton of the Atlantic Ocean Surface (42°N to 33°S)



Abstract
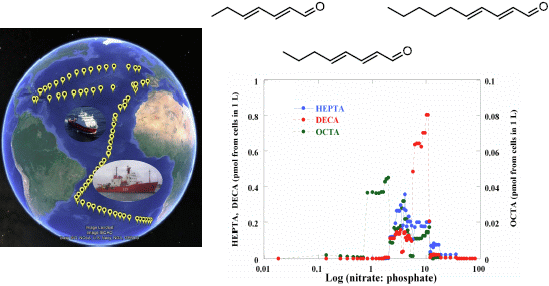
:
1. Introduction
2. Results and Discussion
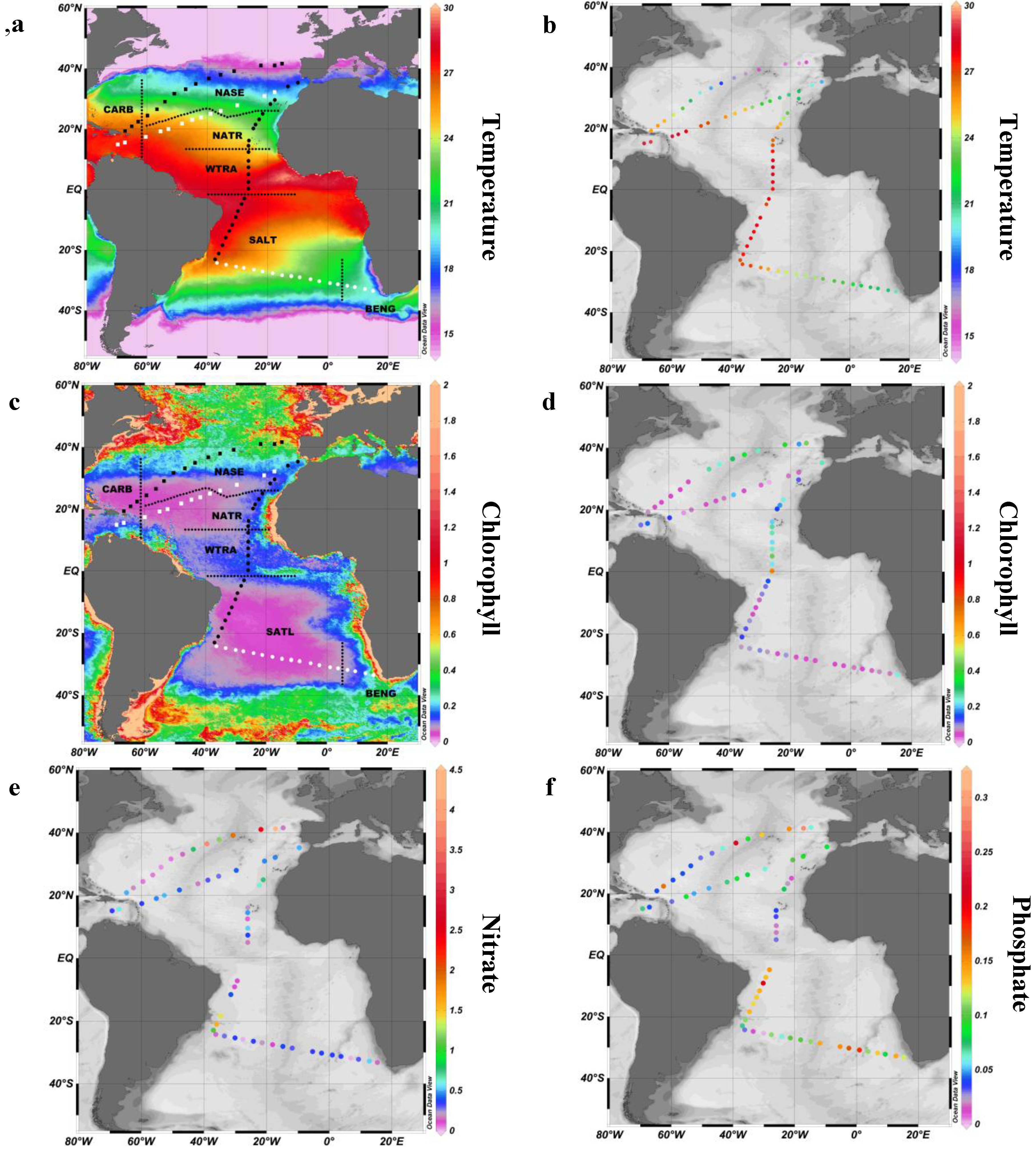
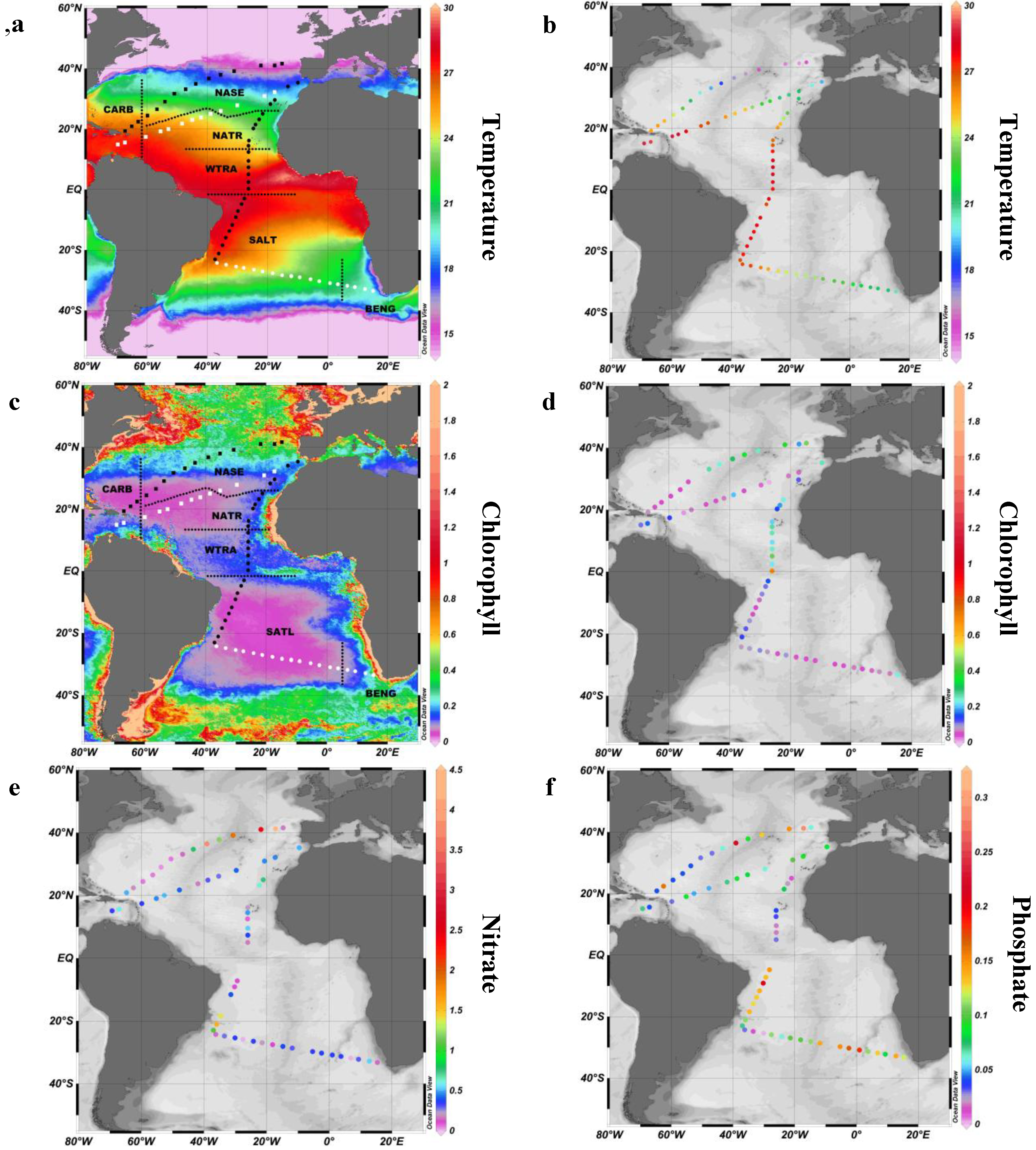
2.1. Study Site
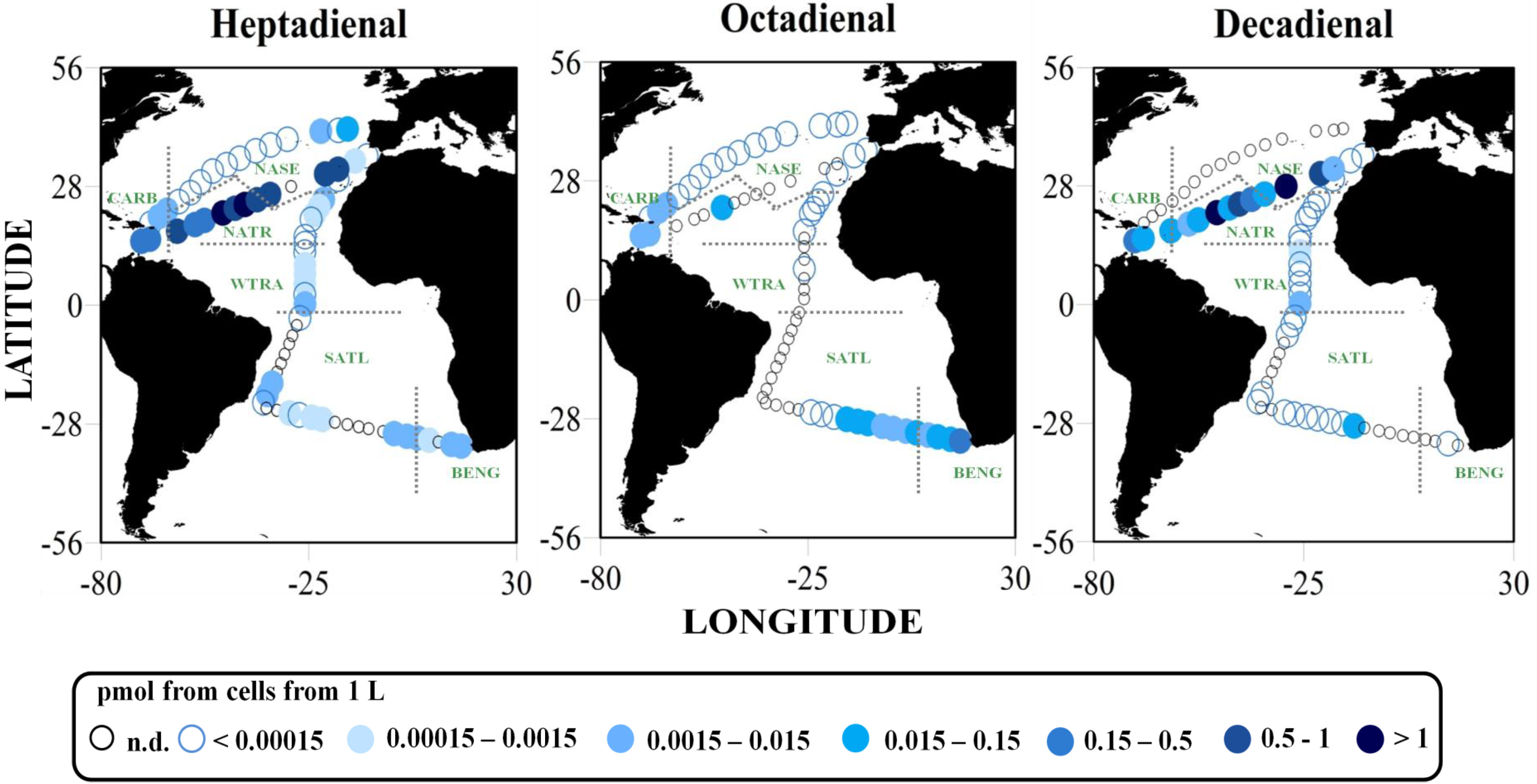
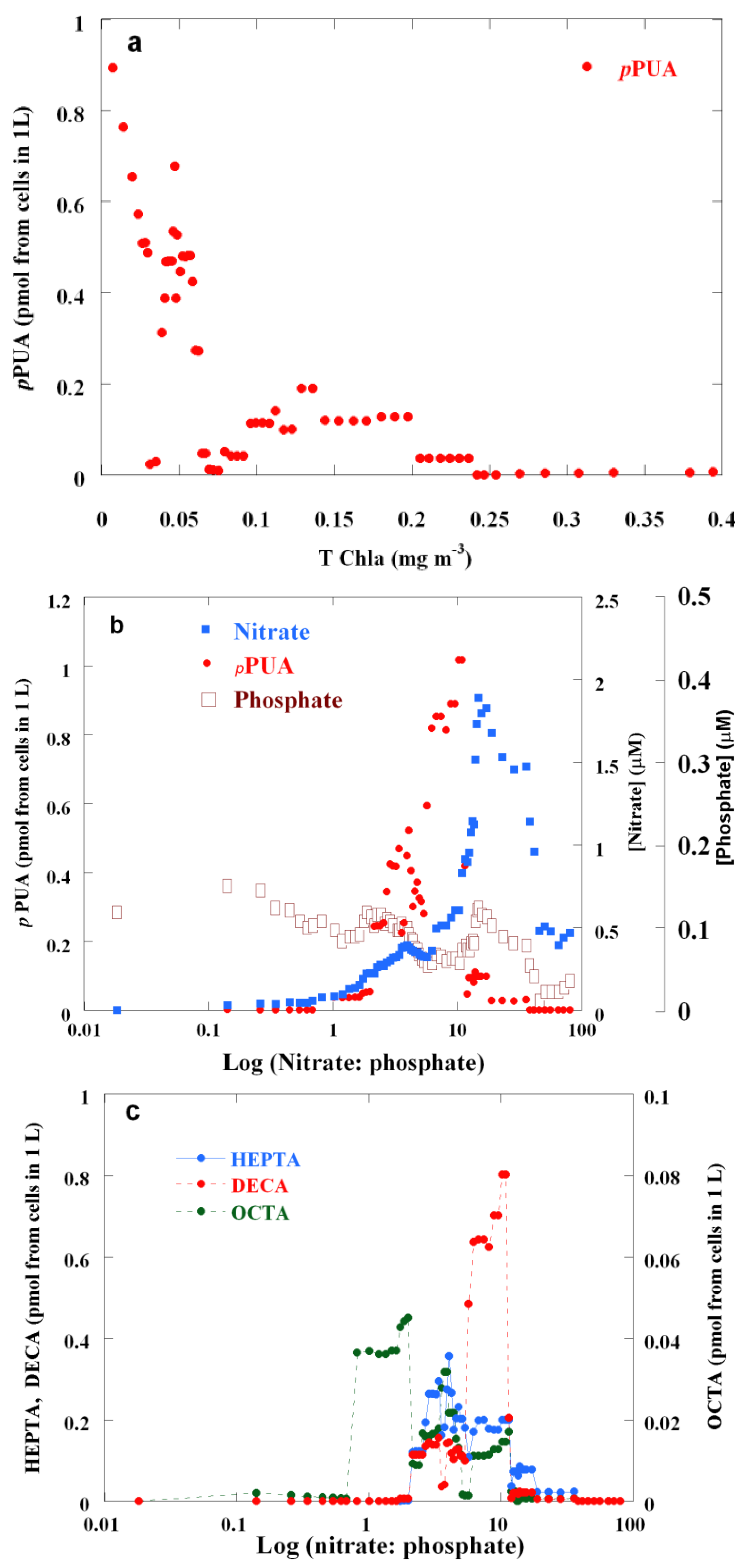
2.2. Surface pPUA Distribution and Biogeographical Patterns

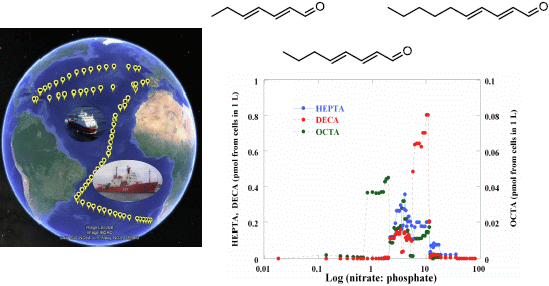{kind=link}
{kind=link}
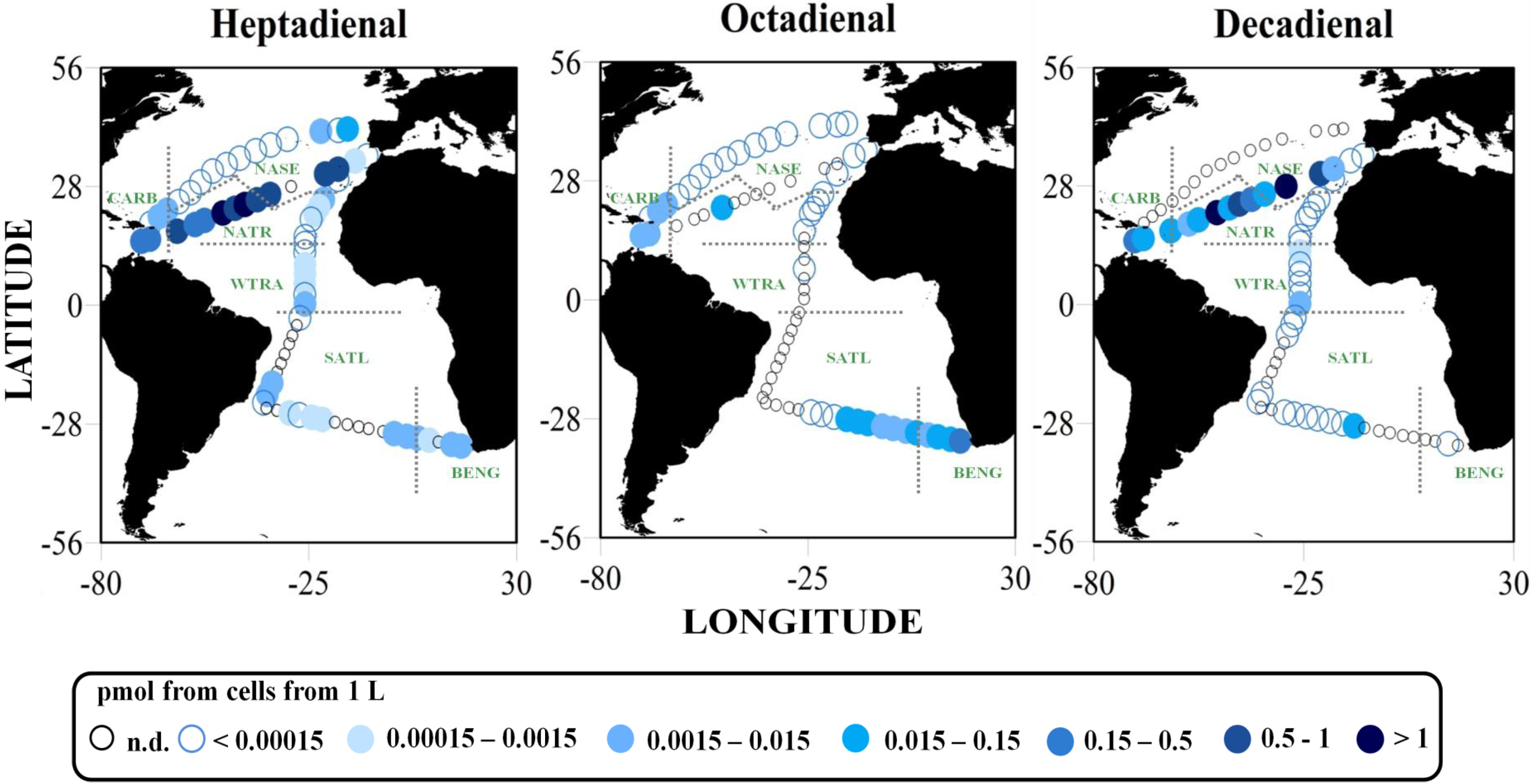{kind=link}
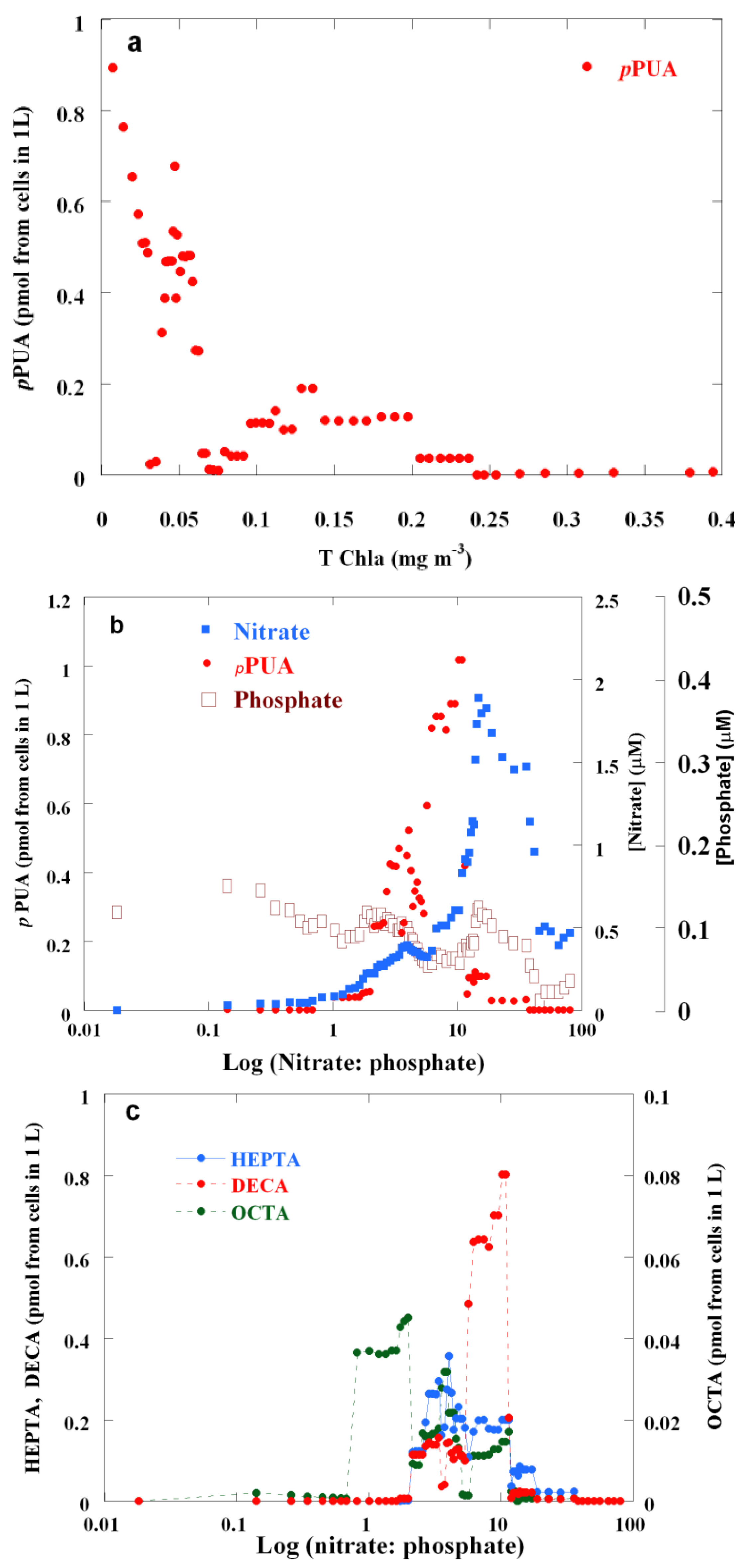{kind=link}
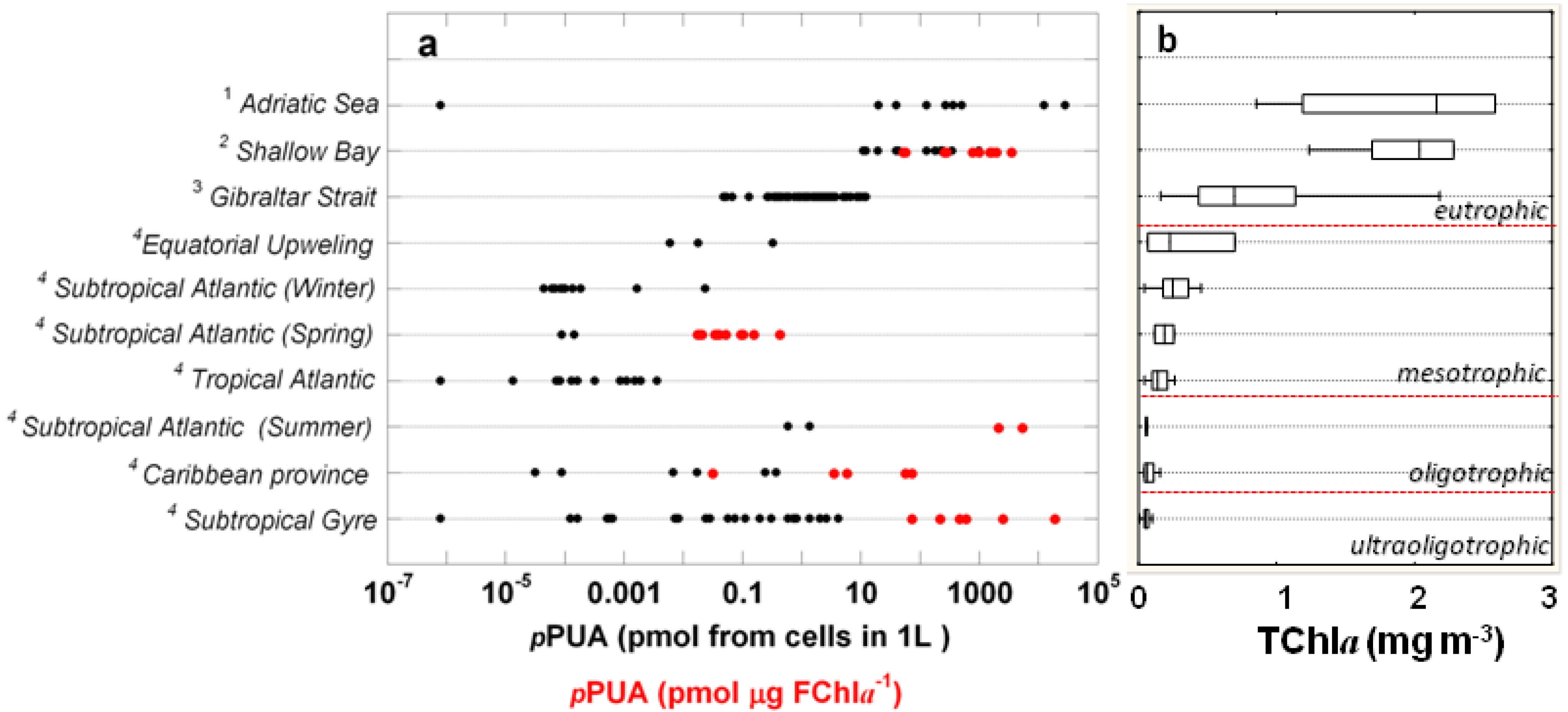{kind=link}
| Province (Acronym) | Date | N (Sites/Viable PUA Samples) | Transect | Season | Temperature (°C) | Salinity | Total Chla (mg m−3) | Nitrate (μM) | Phosphate (μM) |
|---|---|---|---|---|---|---|---|---|---|
| Caribbean (CARB) | 03/23/2011–03/26/2011 | 4/7 | T1 | Spring Boreal | 25.28 ± 0.62 | 36.43 ± 0.34 | 0.05 ± 0.014 | 0.24 ± 0.22 | 0.08 ± 0.06 |
| 06/20/2011–06/21/2011 | 2/2 | T2 | Summer Boreal | 28.95 ± 0.21 | 35.01 ± 0.01 | 0.13 ± 0.033 | 0.45 ± 0.16 | 0.05 ± 0.02 | |
| North Atlantic Subtropical Gyre east (NASE) | 12/16/2010–12/24/2010 | 5/10 | T3 | Winter Boreal | 21.15 ± 1.66 | 36.82 ± 0.24 | 0.009 ± 0.060 | 0.61 ± 0.17 | 0.04 ± 0.04 |
| 03/26/2011–04/10/2011 | 12/22 | T1 | Spring Boreal | 17.5 ± 2.83 | 36.31 ± 0.39 | 0.250 ± 0.127 | 1.43 ± 1.50 | 0.11 ± 0.08 | |
| 06/22/2011–06/30/2011 | 8/8 | T2 | Summer Boreal | 22.47 ± 1.49 | 37.13 ± 0.35 | 0.039 ± 0.021 | 0.40 ± 0.07 | 0.08 ± 0.01 | |
| North Atlantic Tropical Gyral Province (NATR) | 12/25/2010–12/30/2011 | 6/12 | T3 | Winter Boreal | 25.95 ± 1.96 | 36.5 ± 0.43 | 0.210 ± 0.046 | 0.34 ± 0.24 | 0.04 ± 0.02 |
| 06/30/2011–07/11/2011 | 9/9 | T2 | Summer Boreal | 27.16 ± 1.40 | 36.74 ± 0.73 | 0.078 ± 0.062 | 0.36 ± 0.12 | 0.06 ± 0.02 | |
| Western Tropical Atlantic (WTRA) (Equatorial upwelling) | 12/31/2011–01/10/2012 | 7/14 | T3 | Summer Austral | 27.9 ± 0.50 | 35.94 ± 0.67 | 0.293 ± 0.205 | 0.35 ± 0.18 | 0.05 ± 0.06 |
| 27.49 | 36.10 | 0.693 | |||||||
| South Atlantic Gyral Province (SATL) | 01/19/2011–02/02/2011 | 23/45 | T4 | Summer Austral | 25.2 ± 2.24 | 36.43 ± 0.46 | 0.066 ± 0.028 | 0.25 ± 0.13 | 0.10 ± 0.05 |
| Benguela Current Coastal Province (BENG) | 02/02/2011–02/05/2011 | 3/6 | T4 | Summer Austral | 20.96 ± 0.47 | 35.54 ± 0.16 | 0.119 ± 0.090 | 0.32 ± 0.18 | 0.11 ± 0.03 |

3. Experimental Section
3.1. Study Area
3.2. Sampling
3.3. Polyunsaturated Aldehydes Extraction and Quantification
3.4. Statistics
4. Conclusions
Abbreviations
| NASA | National Aeronautics and Space Administration |
| MODIS | Moderate Resolution Imaging Spectroradiometer |
| GSFC | Goddard Space Flight Center |
| CTD | Conductivity/Temperature/Depth profiler |
| GC-MS | Gas Chromatography-Mass spectrometry |
| PTFE | Polytetrafluoroethylene |
| INDIAL | Acronym for the spanish project “Regulación Ambiental e Implicaciones tróficas de la liberación de aldehídos volátiles por diatomeas” |
| CEI·MAR | Campus of International Excellence of the Sea |
Supplementary Files
Acknowledgments
Conflicts of Interest
References
- Hay, M.E. Marine chemical ecology: What’s known and what’s next? J. Exp. Mar. Biol. Ecol. 1996, 200, 103–134. [Google Scholar] [CrossRef]
- Miralto, A.; Barone, G.; Romano, G.; Poulet, S.A.; Ianora, A.; Russo, G.L.; Buttino, I.; Mazzarella, G.; Laabir, G.; Cabrini, M.; et al. The insidious effect of diatoms on copepod reproduction. Nature 1999, 402, 173–176. [Google Scholar] [CrossRef]
- Caldwell, G.S. The influence of bioactive oxylipins from marine diatoms on invertebrate reproduction and development. Mar. Drugs 2009, 7, 367–400. [Google Scholar] [CrossRef]
- Pohnert, G. Wound-activated chemical defense in unicellular planktonic algae. Angew. Chem. Int. Ed. 2000, 39, 4352–4354. [Google Scholar] [CrossRef]
- D’Ippolito, G.; Tucci, S.; Cutignano, A.; Romano, G.; Cimino, G.; Miralto, A.; Fontana, A. The role of complex lipids in the synthesis of bioactive aldehydes of the marine diatom Skeletonema costatum. Biochim. Biophys. Acta 2004, 1686, 100–107. [Google Scholar] [CrossRef]
- Fontana, A.G.; Cutignano, A.; Miralto, A.; Ianora, A.; Romano, G.; Cimino, G. Chemistry of oxilipins pathways in marine diatoms. Pure Appl. Chem. 2007, 79, 475–484. [Google Scholar]
- Kirchman, D.L. Oceanography: phytoplankton death in the sea. Nature 1995, 398, 293–294. [Google Scholar] [CrossRef]
- Suttle, C.A. Viruses in the sea. Nature 2005, 437, 356–360. [Google Scholar] [CrossRef]
- Ianora, S.A.; Poulet, S.A.; Miralto, A. The effects of diatoms on copepod reproduction: A review. Phycologia 2003, 42, 351–363. [Google Scholar] [CrossRef]
- Ribalet, F.; Berges, J.A.; Ianora, A.; Casotti, R. Growth inhibition of cultured marine phytoplankton by toxic algal-derived polyunsaturated aldehydes. Aquat. Toxicol. 2007, 85, 219–227. [Google Scholar] [CrossRef]
- Ianora, A.; Miralto, A. Toxigenic effects of diatoms on grazers, phytoplankton and other microbes: A review. Ecotoxicology 2010, 19, 493–511. [Google Scholar] [CrossRef]
- Romano, G.; Miralto, A.; Ianora, A. Teratogenic effects of diatom metabolites on sea urchin Paracentrotus lividus embryos. Mar. Drugs 2010, 8, 950–967. [Google Scholar] [CrossRef]
- Dutz, J.; Koski, M.; Jónasdóttir, S.H. Copepod reproduction is unaffected by diatom aldehydes or lipid composition. Limnol. Oceanogr. 2008, 53, 225–235. [Google Scholar] [CrossRef]
- Flynn, K.; Irigoien, X. Aldehyde-induced insidious effects cannot be considered as a diatom defence mechanism against copepods. Mar. Ecol. Prog. Ser. 2009, 377, 79–89. [Google Scholar] [CrossRef]
- Balestra, C.; Alonso-Sáez, L.; Gasol, J.M.; Casotti, R. Group-specific effects on coastal bacterioplankton of polyunsaturated aldehydes produced by diatoms. Inter-Res. Aquat. Microb. Ecol. 2011, 63, 123–131. [Google Scholar] [CrossRef]
- Paul, C.; Reunamo, A.; Lindehoff, E.; Bergvisk, J.; Mausz, M.A.; Larsson, H.; Richter, H.; Wängberg, S.A.; Leskinen, P.; Bảmstedt, U.; et al. Diatom derived polyunsaturated aldehydes do not structure the planktonic microbial community in a mesocosm study. Mar. Drugs 2012, 10, 775–792. [Google Scholar] [CrossRef]
- Vardi, A.; Formiggini, F.; Casotti, R.; De Martino, A.; Ribalet, F.; Miralto, A.; Bowler, C. A stress surveillance system based on calcium and nitric oxide in marine diatoms. PLoS Biol. 2006, 4, 410–419. [Google Scholar] [CrossRef]
- Vidoudez, C.; Casotti, R.; Bastianini, M.; Pohnert, G. Quantification of dissolved and particulate polyunsaturated aldehydes in the Adriatic Sea. Mar. Drugs 2011, 9, 500–513. [Google Scholar] [CrossRef]
- Wichard, T.; Poulet, S.A.; Halsband-Lenk, C.; Albaina, A.; Harris, R.; Liu, D.; Pohnert, G. Survey of the chemical defense potential of diatoms: Screening of fifty one species for α,β,γ,δ-unsaturated aldehydes. J. Chem. Ecol. 2005, 31, 949–958. [Google Scholar] [CrossRef]
- Pohnert, G. Chemical noise in the silent ocean. J. Plankton Res. 2010, 32, 141–144. [Google Scholar] [CrossRef]
- Chisholm, S.W. What limits phytoplankton growth? Oceanus 1992, 35, 36–46. [Google Scholar]
- Li, W.K.W. Macroecological patterns of phytoplankton in the northwestern North Atlantic Ocean. Nature 2002, 419, 154–157. [Google Scholar] [CrossRef]
- Longhurst, A.R. The Atlantic Ocean. In Ecological Geography of the Sea; Academic Press: San Diego, CA, USA, 1998; pp. 131–268. [Google Scholar]
- MODIS Web. Available online: http://modis.gsfc.nasa.gov (accessed on 17 May 2013).
- Acevedo-Trejos, E.; Brandt, G.; Merico, A.; Smith, S.L. Biogeographical patterns of phytoplankton community size structure in the oceans. Global Ecol. Biogeogr. 2013, 22, 1060–1070. [Google Scholar] [CrossRef]
- Taylor, R.L.; Abrahamsoom, K.; Godhe, A.; Wängberg, S.A. Seasonal variability in polyunsaturated aldehyde production potential among strains of Skeletonema marinoi (Bacillariphyceae). J. Phycol. 2009, 45, 46–53. [Google Scholar] [CrossRef]
- Bartual, A.; Ortega, M.J. Temperature differentially affects the persistence of polyunsaturated aldehydes in seawater. Environ. Chem. 2013, 10, 403–408. [Google Scholar] [CrossRef]
- Marañón, E.; Holligan, P.M.; Barciela, R.; González, N.; Mouriño, B.; Pazó, M.J.; Varela, M. Patterns of phytoplankton size structure and productivity in contrasting open-ocean environments. Mar. Ecol. Prog. Ser 2001, 216, 43–56. [Google Scholar] [CrossRef]
- Férnandez, A.; Mouriño-Carballido, B.; Bode, A.; Varela, M.; Marañón, E. Latitudinal distribution of Trichodesmium spp. and N2 fixation in the Atlantic Ocean. Biogeosci. Discuss 2010, 7, 2195–2225. [Google Scholar] [CrossRef]
- Capone, D.G.; Burns, J.A.; Montoya, J.P.; Subramaniam, A.; Mahaffey, C.; Gunderson, T.; Michaels, A.F.; Carpenter, E.J. Nitrogen fixation by Trichodesmium spp.: An important source of new nitrogen to the tropical and subtropical North Atlantic Ocean. Global Biogeochem. Cycles 2005, 19. [Google Scholar] [CrossRef]
- Post, A.F.; Dedej, Z.; Gottlieb, R.; Li, H.; Thomas, D.N.; El-Absawi, M.; El-Naqqar, A.; El-Gharabawi, M.; Sommer, U. Spatial and temporal distribution of Trichodesmium spp. in the stratified Gulf of Aqaba, Red Sea. Mar. Ecol. Prog. Ser. 2002, 239, 241–250. [Google Scholar] [CrossRef]
- Hansen, E.; Ernstsen, A.; Eilertsen, H.C. Isolation and characterisation of a cytotoxic polyunsaturated aldehyde from the marine phytoplankter Phaeocystis pouchetii (Hariot) Lagerheim. Toxicology 2004, 199, 207–217. [Google Scholar] [CrossRef]
- Shushkina, E.A.; Vinogradov, M.E.; Lebedeva, L.P.; Anokhina, L.L. Productivity characteristics of epipelagic communities of the World’s Oceans. Oceanology 1997, 37, 346–353. [Google Scholar]
- Ribalet, F.; Vidoudez, C.; Cassin, D.; Pohnert, G.; Ianora, A.; Miralto, A.; Casotti, R. High plasticity in the production of diatom-derived polyunsaturated aldehydes under nutrient limitation: Physiological and ecological implications. Protist 2009, 160, 444–451. [Google Scholar] [CrossRef]
- Vidoudez, C.; Pohnert, G. Comparative metabolomics of the diatom Skeletonema marinoi in different growth phases. Metabolomics 2012, 8, 654–669. [Google Scholar] [CrossRef]
- Ribalet, F.; Wichard, T.; Pohnert, G.; Ianotar, A.; Miralto, A.; Casotti, R. Age and nutrient limitation enhance polyunsaturated aldehyde production in marine diatoms. Phytochemistry 2007, 68, 2059–2067. [Google Scholar] [CrossRef]
- Martin, P.; Van Mooy, B.A.S.; Heithoff, A.; Dyhrman, S.T. Phosphorus supply drives rapid turnover of membrane phospholipids in the diatom Thalassiosira pseudonana. ISME J. 2011, 5, 1057–1060. [Google Scholar] [CrossRef]
- Pohnert, G. Phospholipase A2 activity triggers the wound-activated chemical defense in the diatom Thalassiosira rotula. Plant Physiol. 2002, 129, 103–111. [Google Scholar] [CrossRef]
- Litchman, E.; Klausmeier, C.A.; Yoshiyama, K. Contrasting size evolution in marine and freshwater diatoms. Proc. Natl. Acad. Sci. USA 2013, 106, 2665–2670. [Google Scholar]
- Platt, T.; Subba Pao, D.V.; Irwin, B. Photosynthesis of picoplankton in the oligotrophic ocean. Nature 1983, 301, 702–704. [Google Scholar] [CrossRef]
- Granéli, E.; Weberg, M.; Salomon, P.S. Harmful algal blooms of allelopathic microalgal species: The role of eutrophication. Harmful Algae 2008, 8, 94–102. [Google Scholar] [CrossRef]
- Giovanni: The Bridge between Data and Science. Available online: http://giovanni.gsfc.nasa.gov/giovanni/ (accessed on 22 January 2013).
- Morillo-García, S.; University of Cádiz, Puerto Real, Cádiz, Spain. Papel de los aldehídos poli-insaturados (PUAs) en ecosistemas planctónicos marinos. 2013; unpublished work. [Google Scholar]
- Morillo-García, S.; Valcárcel-Pérez, N.; Cózar, A.; Ortega, M.J.; Macías, D.; Ramírez-Romero, E.; García, C.M.; Echevarría, F.; Bartual, A. Potential polyunsaturated aldehydes in the Strait of Gibraltar under two tidal regimes. Mar. Drugs 2014. submitted for publication. [Google Scholar]
- Yentsch, C.S.; Menzel, D.W. A method for the determination of phytoplankton chlorophyll and phaeophytin by fluorescence. Deep Sea Res. 1963, 10, 221–231. [Google Scholar]
- Grasshoff, K.; Ehrhardt, M.; Kremling, K. Methods of Seawater Analysis, 3rd ed.; Wiley VCH: Weinheim, Germany, 2007. [Google Scholar]
- Moreno-Ostos, E. Libro blanco de métodos y técnicas de trabajo oceanográfico. In Proceeding of Expedición de Circunnavegación MALASPINA 2010 (Cambio Global y Exploración de la Biodiversidad del Océano), CSIC, Madrid, Spain, 22 May 2012; p. 688.
- Wichard, T.; Poulet, S.; Pohnert, G. Determination and quantification of α,β,γ,δ-unsaturated aldehydes as pentafluorobenzyl-oxime derivates in diatom cultures and natural phytoplankton populations: Application in marine field studies. J. Chromatogr. B 2005, 814, 155–161. [Google Scholar] [CrossRef]
© 2014 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Bartual, A.; Arandia-Gorostidi, N.; Cózar, A.; Morillo-García, S.; Ortega, M.J.; Vidal, M.; Cabello, A.M.; González-Gordillo, J.I.; Echevarría, F. Polyunsaturated Aldehydes from Large Phytoplankton of the Atlantic Ocean Surface (42°N to 33°S). Mar. Drugs 2014, 12, 682-699. https://doi.org/10.3390/md12020682
Bartual A, Arandia-Gorostidi N, Cózar A, Morillo-García S, Ortega MJ, Vidal M, Cabello AM, González-Gordillo JI, Echevarría F. Polyunsaturated Aldehydes from Large Phytoplankton of the Atlantic Ocean Surface (42°N to 33°S). Marine Drugs. 2014; 12(2):682-699. https://doi.org/10.3390/md12020682
Chicago/Turabian StyleBartual, Ana, Néstor Arandia-Gorostidi, Andrés Cózar, Soledad Morillo-García, María Jesús Ortega, Montserrat Vidal, Ana María Cabello, Juan Ignacio González-Gordillo, and Fidel Echevarría. 2014. "Polyunsaturated Aldehydes from Large Phytoplankton of the Atlantic Ocean Surface (42°N to 33°S)" Marine Drugs 12, no. 2: 682-699. https://doi.org/10.3390/md12020682
APA StyleBartual, A., Arandia-Gorostidi, N., Cózar, A., Morillo-García, S., Ortega, M. J., Vidal, M., Cabello, A. M., González-Gordillo, J. I., & Echevarría, F. (2014). Polyunsaturated Aldehydes from Large Phytoplankton of the Atlantic Ocean Surface (42°N to 33°S). Marine Drugs, 12(2), 682-699. https://doi.org/10.3390/md12020682