
Squid Pen Chitin Chitooligomers as Food Colorants Absorbers



Abstract
:1. Introduction

2. Results and Discussion
2.1. Isolation and Identification of a Chitosanase-Producing Strain
2.2. TKU033 Chitosanase Production and Purification

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Step | Total Protein (mg) | Total Activity (U) | Specific Activity (U/mg) | Purification Fold | Yield (%) |
|---|---|---|---|---|---|
| Culture supernatant | 4032.0 | 19.4 | 0.005 | 1.0 | 100 |
| (NH4)2SO4 ppt | 838.4 | 5.9 | 0.007 | 1.4 | 30 |
| DEAE-Sepharose | 46.5 | 1.7 | 0.037 | 7.4 | 9 |
| Macro-prep DEAE | 5.8 | 0.3 | 0.052 | 10.4 | 2 |

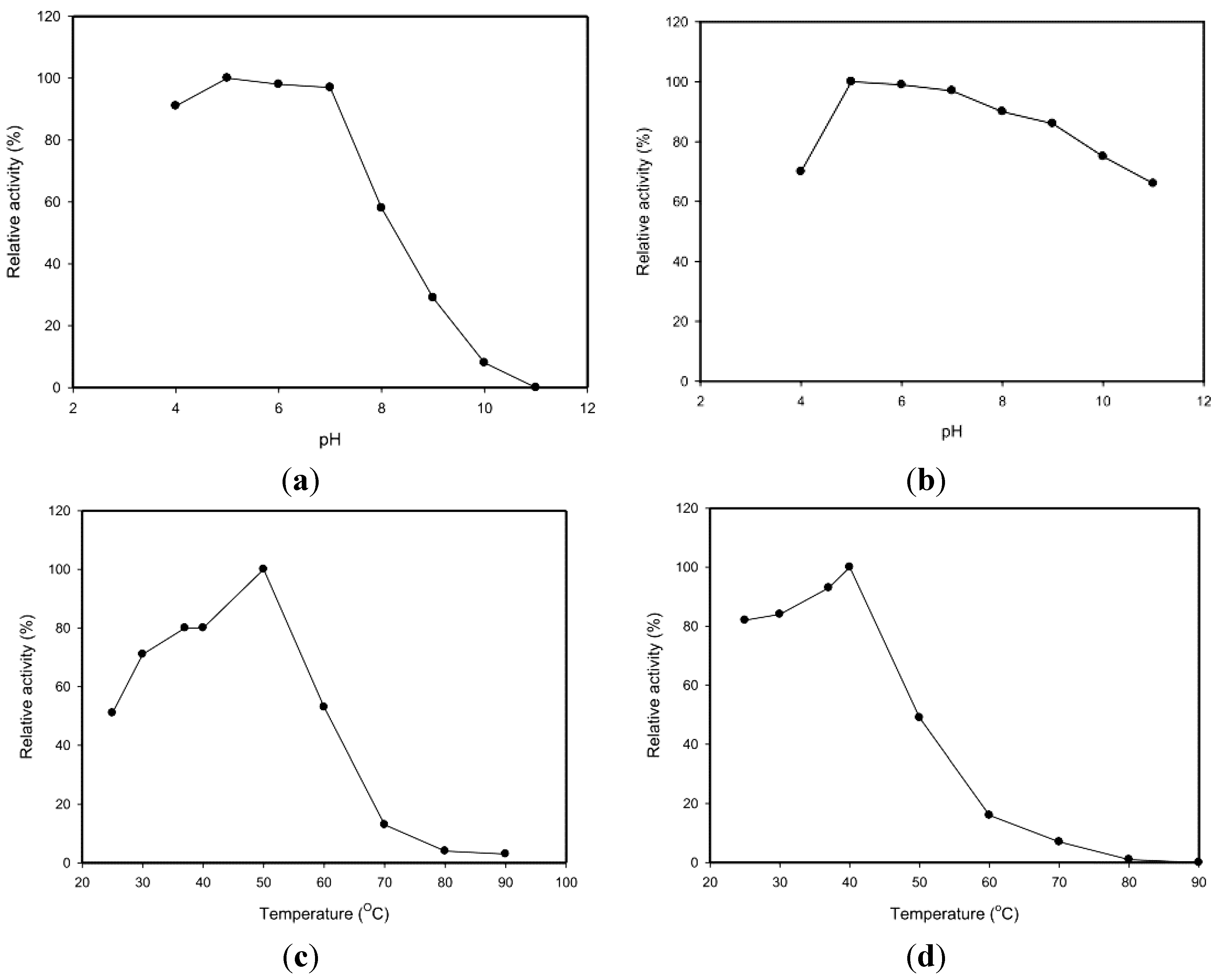
2.3. Effects of pH and Temperature

2.4. Substrate Specificity
2.5. Effects of Various Inhibitors and Metal Ions
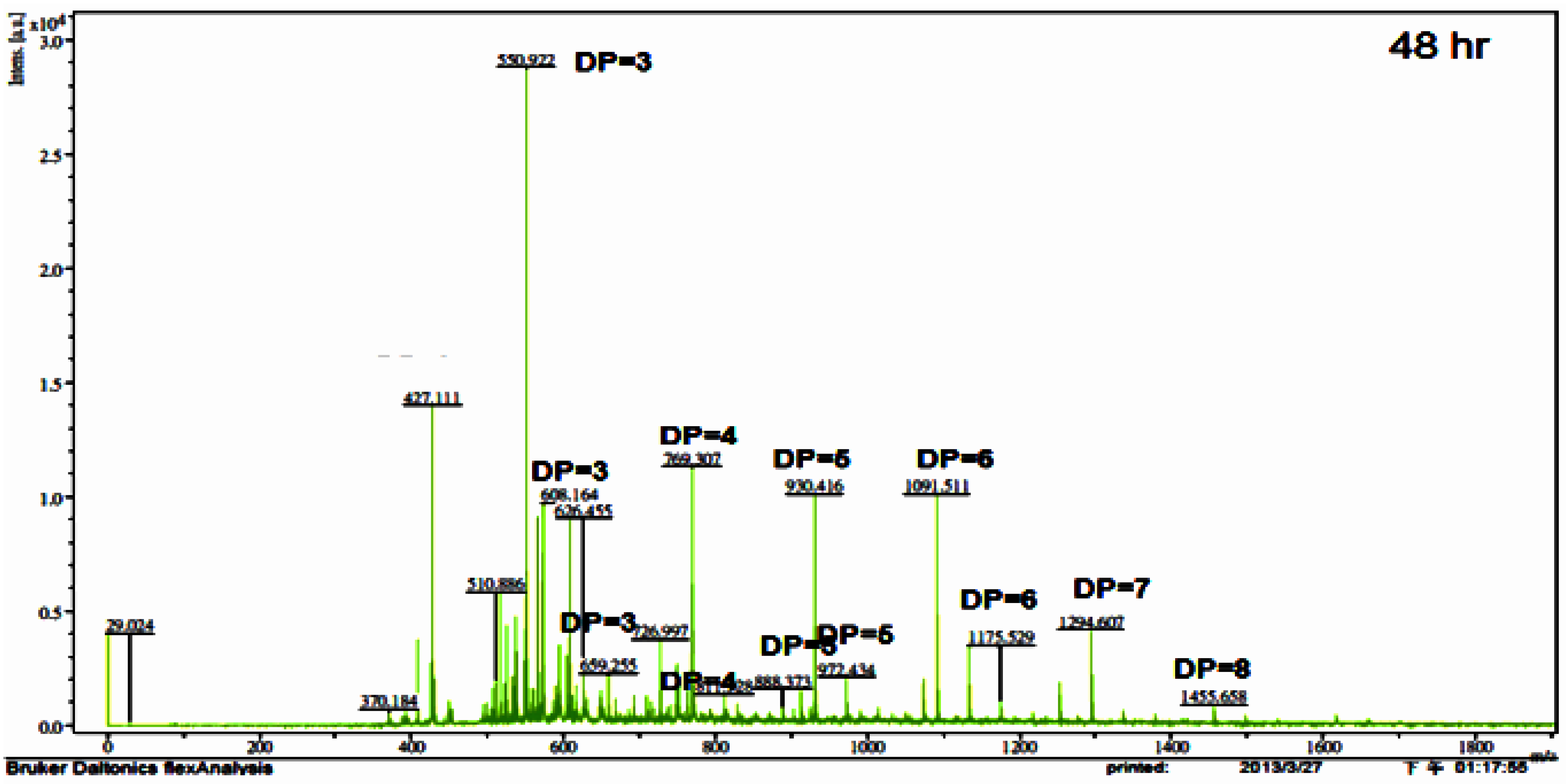
2.6. COS Preparation and Product Analysis

| m/z | Ion Composition | DP | Hydrolysis Time (h) | |||||
|---|---|---|---|---|---|---|---|---|
| 2 | 4 | 6 | 12 | 24 | 48 | |||
| 550 | (GlcN)2-GlcNAc | 3 | + | + | + | + | + | |
| 608 | GlcN-(GlcNAc)2 | 3 | + | + | + | + | ||
| 659 | (GlcNAc)3 | 3 | + | + | + | |||
| 692 | (GlcN)4 | 4 | + | |||||
| 727 | (GlcN)3-GlcNAc | 4 | + | + | ||||
| 769 | (GlcN)2-(GlcNAc)2 | 4 | + | + | + | + | + | + |
| 811 | GlcN-(GlcNAc)3 | 4 | + | + | + | + | ||
| 888 | (GlcN)4-GlcNAc | 5 | + | + | + | |||
| 930 | (GlcN)3-(GlcNAc)2 | 5 | + | + | + | + | + | + |
| 972 | (GlcN)2-(GlcNAc)3 | 5 | + | + | + | + | + | |
| 1091 | (GlcN)4-(GlcNAc)2 | 6 | + | + | + | |||
| 1133 | (GlcN)3-(GlcNAc)3 | 6 | + | + | + | |||
| 1175 | (GlcN)2-(GlcNAc)4 | 6 | + | |||||
| 1294 | (GlcN)4-(GlcNAc)3 | 7 | + | + | + | + | + | + |
| 1455 | (GlcN)5-(GlcNAc)3 | 8 | + | + | + | |||
| 1616 | (GlcN)6-(GlcNAc)3 | 9 | + | + | ||||
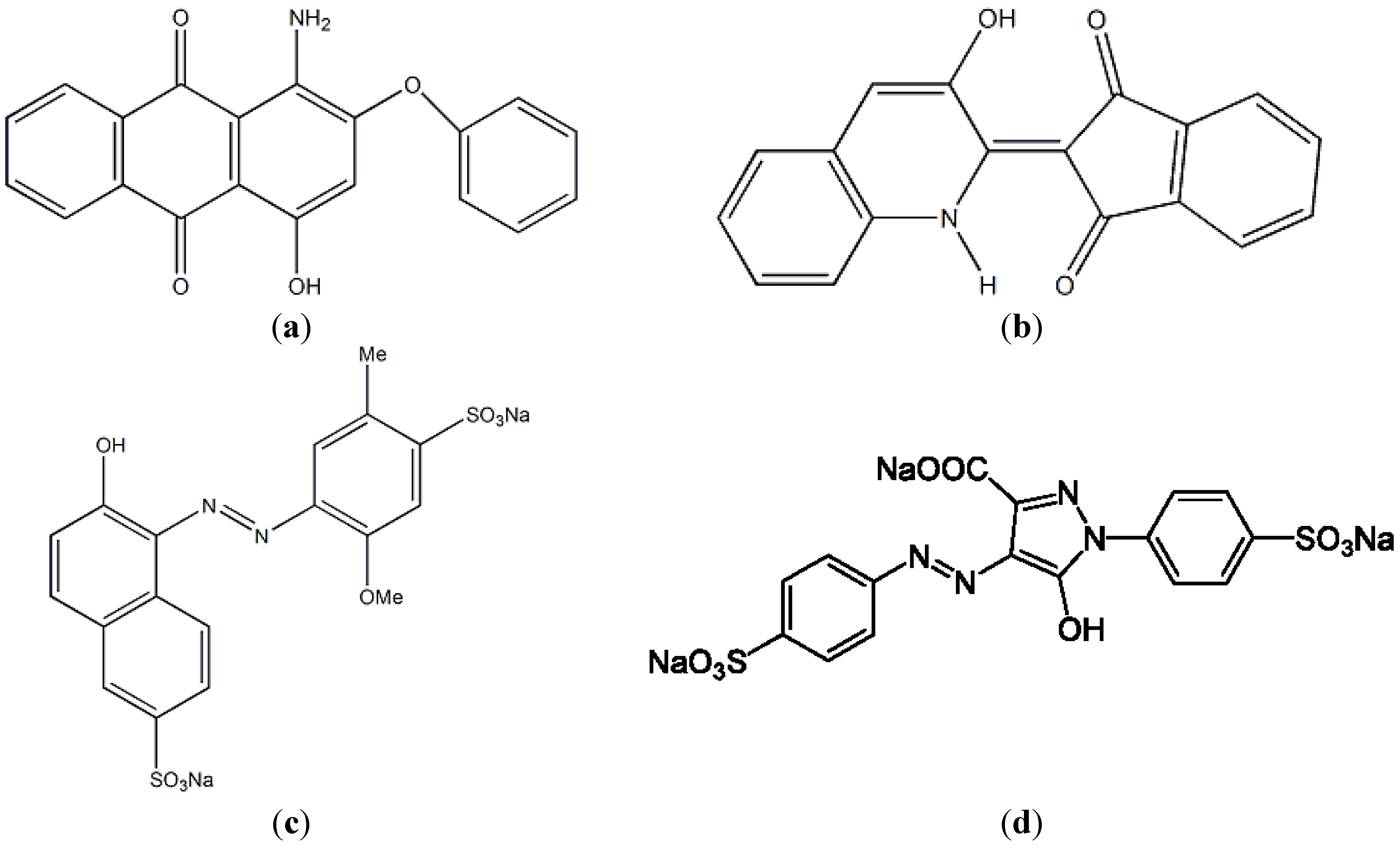
2.7. Fermented SPP as a Biosorbent for Dye Removal
| Substrate | Adsorption (%) | |
|---|---|---|
| Fermented SPP | Unfermented SPP | |
| Disperse red 60 | 87 | 70 |
| Disperse yellow 54 | 96 | 80 |
| Food colorant (R40) | 90 | 95 |
| Food colorant (Y4) | 68 | 82 |

3. Materials and Methods
3.1. Materials
3.2. Screening and Identification of Chitosanase-Producing Strains
3.3. Preparation of the TKU033 Chitosanase
3.4. Measurement of Chitosanase Activity
3.5. Purification of the TKU033 Chitosanase
3.6. Protein Determination
3.7. Determination of Molecular Mass
3.8. Effects of pH and Temperature on Enzyme Activities
3.9. Effects of Various Chemicals and Surfactants on Enzyme Activities
3.10. Measurement of Total Sugars
3.11. Adsorption Effects of SPP and Fermented SPP for Dye Removal
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Chiang, C.L.; Chang, C.T.; Sung, H.Y. Purification and properties of chitosanase from a mutant of Bacillus subtilis IMR-NK1. Enzyme Microb. Technol. 2003, 32, 260–267. [Google Scholar] [CrossRef]
- Gao, X.A.; Ju, W.T.; Jung, W.J.; Park, R.D. Purification and characterization of chitosanase from Bacillus cereus D-11. Carbohydr. Polym. 2008, 72, 513–520. [Google Scholar] [CrossRef]
- Jiang, X.; Chen, D.; Chen, L.; Yang, G.; Zou, S. Purification, characterization, and action mode of a chitosanase from Streptomyces roseolus induced by chitin. Carbohydr. Res. 2012, 355, 40–44. [Google Scholar] [CrossRef] [PubMed]
- Kurakake, M.; Yo-U, S.; Nakagawa, K.; Sugihara, M.; Komaki, T. Properties of chitosanase from Bacillus cereus S1. Curr. Microbiol. 2000, 40, 6–9. [Google Scholar] [CrossRef] [PubMed]
- Kobayashi, T.; Koide, O.; Deguchi, S.; Horikoshi, K. Characterization of chitosanase of a deep biosphere Bacillus strain. Biosci. Biotechnol. Biochem. 2011, 75, 669–673. [Google Scholar] [CrossRef] [PubMed]
- Pechsrichuang, P.; Yoohat, K.; Yamabhai, M. Production of recombinant Bacillus subtilis chitosanase, suitable for biosynthesis of chitosan-oligosaccharides. Bioresour. Technol. 2013, 127, 407–414. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.L.; Su, J.W.; Liang, T.W.; Nguyen, A.D.; Wang, S.L. Production, purification and characterization of a chitosanase from Bacillus cereus. Res. Chem. Intermed. 2014, 40, 2237–2248. [Google Scholar] [CrossRef]
- Wang, C.L.; Chen, C.J.; Nguyen, A.D.; Liang, T.W.; Twu, Y.K.; Huang, S.Y.; Wang, S.L. Environmental chitinous materials as adsorbents for the one-step purification of protease and chitosanase. Res. Chem. Intermed. 2014, 40, 2363–2370. [Google Scholar] [CrossRef]
- Wang, S.L.; Liang, T.W.; Yen, Y.H. Bioconversion of chitin-containing wastes for the production of enzymes and bioactive materials. Carbohydr. Polym. 2011, 84, 732–742. [Google Scholar] [CrossRef]
- La Thi, K.N.; Wang, S.L.; Dinh, M.H.; Phung, M.L.; Nguyen, T.V.; Tran, M.D.; Nguyen, A.D. Preparation of chitosan nanoparticles by spray dryer and their antibacterial activity. Res. Chem. Intermed. 2014, 40, 2165–2176. [Google Scholar] [CrossRef]
- Wang, S.L.; Liu, K.C.; Liang, T.W.; Kuo, Y.H.; Wang, C.Y. In vitro antioxidant activity of liquor and semi-purified fractions from squid pen biowaste by Serratia ureilytica TKU013. Food Chem. 2010, 119, 1380–1385. [Google Scholar] [CrossRef]
- Busilacchi, A.; Gigante, A.; Mattioli-Belmonte, M.; Manzotti, S.; Muzzarelli, R.A.A. Chitosan stabilizes platelet growth factors and modulates stem cell differentiation toward tissue regeneration. Carbohydr. Polym. 2013, 98, 665–676. [Google Scholar] [CrossRef] [PubMed]
- Liang, T.W.; Chen, Y.J.; Yen, Y.H.; Wang, S.L. The antitumor activity of the hydrolysates of chitinous materials hydrolyzed by crude enzyme from Bacillus amyloliquefaciens V656. Process Biochem. 2007, 42, 527–534. [Google Scholar] [CrossRef]
- Wang, S.L.; Lin, T.Y.; Yen, Y.H.; Liao, H.F.; Chen, Y.J. Bioconversion of shellfish chitin wastes for the production of Bacillus subtilis W-118 chitinase. Carbohydr. Res. 2006, 341, 2507–2515. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.L.; Lin, H.T.; Liang, T.W.; Chen, Y.J.; Yen, Y.H.; Guo, S.P. Reclamation of chitinous materials by bromelain for the preparation of antitumor and antifungal materials. Bioresour. Technol. 2008, 99, 4386–4393. [Google Scholar] [CrossRef] [PubMed]
- Dotto, G.L.; Pinto, L.A.A. Adsorption of food dyes onto chitosan: Optimization process and kinetic. Carbohydr. Polym. 2011, 84, 231–238. [Google Scholar] [CrossRef]
- Wan Ngah, W.S.; Ariff, N.F.M.; Hanafiah, M.A.K.M. Preparation, characterization, and environmental application of crosslinked chitosan-coated bentonite for tartrazine adsorption from aqueous solutions. Water Air Soil Pollut. 2010, 206, 225–236. [Google Scholar] [CrossRef]
- Wang, S.L.; Chen, S.Y.; Yen, Y.H.; Liang, T.W. Utilization of chitinous materials in pigment adsorption. Food Chem. 2012, 135, 1134–1140. [Google Scholar] [CrossRef] [PubMed]
- Jung, W.J.; Jo, G.H.; Kuk, J.H.; Kim, Y.J.; Oh, K.T.; Park, R.D. Production of chitin from red crab shell waste by successive fermentation with Lactobacillus paracasei KCTC-3037 and Serratia marcescens FS-3. Carbohydr. Polym. 2007, 68, 746–750. [Google Scholar] [CrossRef]
- Liang, T.W.; Chen, Y.Y.; Pan, P.S.; Wang, S.L. Purification of chitinase/chitosanase from Bacillus cereus and discovery of an enzyme inhibitor. Int. J. Biol. Macromol. 2014, 63, 8–14. [Google Scholar] [CrossRef] [PubMed]
- Katsumi, T.; Lacombe-Harvey, M.; Tremblay, H.; Brzezinski, R.; Fukamizo, T. Role of acidic amino acid residues in chitooligosaccharide-binding to Streptomyces sp. N174 chitosanase. Biochem. Biophys. Res. Commun. 2005, 338, 1839–1844. [Google Scholar] [CrossRef] [PubMed]
- Shimosaka, M.; Nogawa, M.; Wang, X.Y.; Kumehara, M.; Okazaki, M. Production of two chitosanases from a chitosan-assimilating bacterium, Acinetobacter sp. Strain CHB101. Appl. Environ. Microbiol. 1995, 61, 438–442. [Google Scholar] [PubMed]
- Chen, X.; Xia, W.; Yu, X. Purification and characterization of two types of chitosanase from Aspergillus sp. CJ22–326. Food Res. Int. 2005, 38, 315–322. [Google Scholar] [CrossRef]
- Wang, S.L.; Pen, J.H.; Liang, T.W.; Liu, K.C. Purification and characterization of a chitosanase from Serratia marcescens TKU011. Carbohydr. Res. 2008, 343, 1316–1323. [Google Scholar] [CrossRef] [PubMed]
- Fenton, D.M.; Eveleigh, D.E. Purification and mode of action of a chitosanase from Penicillium islandicum. J. Gen. Microbiol. 1981, 126, 151–165. [Google Scholar]
- Alfonos, C.; Martines, M.; Reyes, F. Purification and properties of two endochitosananse from Mucor rouxii implication on its cell wall degradation. FEMS Microbiol. Lett. 1992, 95, 187–194. [Google Scholar] [CrossRef]
- Cheng, C.Y.; Li, Y. An Aspergillus chitosanase with potential for large-scale preparation of chitosan oligosaccharides. Biotechnol. Appl. Biochem. 2000, 32, 197–203. [Google Scholar] [CrossRef] [PubMed]
- Nogawa, M.; Takahashi, H.; Kashiwagi, A.; Ohshima, K.; Okada, H.; Morikawa, Y. Purification and characterization of exo-β-d-glucosaminidase from a cellulolytic fungus, Trichoderma reesei PC-3–7. Appl. Environ. Microbiol. 1998, 64, 890–895. [Google Scholar] [PubMed]
- Wang, J.; Zhou, W.; Yuan, H.; Wang, Y. Characterization of a novel fungal chitosanase Csn2 from Gongronella sp. JG. Carbohydr. Res. 2008, 343, 2583–2588. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.Y.; Dai, A.L.; Zhang, X.K.; Kuroiwa, K.; Kodaira, R.; Shimosaka, M.; Okazaki, M. Purification and characterization of chitosanase and exo-β-d-glucoseaminidase from a koji mold, Aspergillus oryza IAM2660. Biosci. Biotechnol. Biochem. 2000, 64, 1896–1902. [Google Scholar] [CrossRef] [PubMed]
- Zhou, W.; Yuan, H.; Wang, J.; Yao, J. Production, purification and characterization of chitosanase produced by Gongronella sp. JG. Lett. Appl. Microbiol. 2008, 46, 49. [Google Scholar] [PubMed]
- Dubois, M.; Gilles, K.A.; Hamilton, J.K.; Rebers, P.A.; Smith, F. Colorimetric method for determination of sugars and related substances. Anal. Chem. 1956, 28, 350–356. [Google Scholar] [CrossRef]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liang, T.-W.; Huang, C.-T.; Dzung, N.A.; Wang, S.-L. Squid Pen Chitin Chitooligomers as Food Colorants Absorbers. Mar. Drugs 2015, 13, 681-696. https://doi.org/10.3390/md13010681
Liang T-W, Huang C-T, Dzung NA, Wang S-L. Squid Pen Chitin Chitooligomers as Food Colorants Absorbers. Marine Drugs. 2015; 13(1):681-696. https://doi.org/10.3390/md13010681
Chicago/Turabian StyleLiang, Tzu-Wen, Chih-Ting Huang, Nguyen Anh Dzung, and San-Lang Wang. 2015. "Squid Pen Chitin Chitooligomers as Food Colorants Absorbers" Marine Drugs 13, no. 1: 681-696. https://doi.org/10.3390/md13010681