
The Mucus of Actinia equina (Anthozoa, Cnidaria): An Unexplored Resource for Potential Applicative Purposes

 ,
,  ,
,  and
and
Abstract
:1. Introduction
2. Results
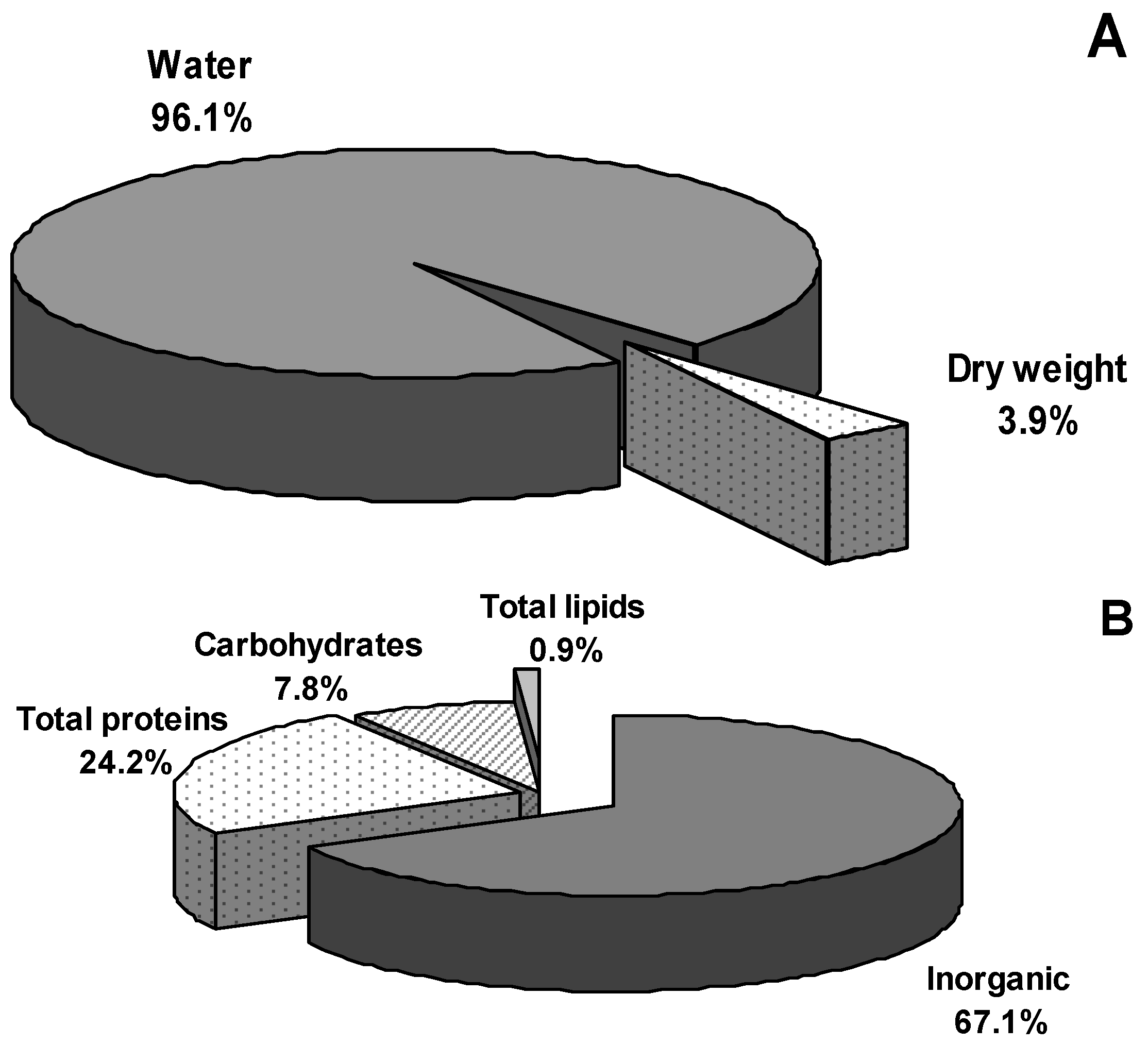
2.1. Mucus Viscosity, Osmolarity, and Electrical Conductivity

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Physico-Chemical Feature | Mean ± SD |
|---|---|
| Inorganic matter (%) | 67.1 ± 2.3 |
| Organic matter (%) | 32.9 ± 0.2 |
| Viscosity 20 °C (cps) | 2.1 ± 0.02 |
| Osmolarity (mOsmol/L) | 1205 ± 10 |
| Conductivity (mS·cm−1) | 124 ± 4.0 |
2.2. Water and Inorganic Content

| Element | Content (%) |
|---|---|
| Cl | 44.48 ± 0.12 |
| Na | 13.38 ± 0.11 |
| Mg | 2.41 ± 0.03 |
| H | 1.53 ± 0.10 |
| K | 2.11 ± 0.02 |
| Ca | 0.71 ± 0.02 |
| C | 2.13 ± 0.02 |
| N | 0.45 ± 0.02 |
| Zn | 0.06 ± 0.005 |
| Cu | absent |
| Fe | absent |
| P | absent |
| Se | absent |
| Sn | absent |
2.3. Protein, Carbohydrate, and Lipids Concentration

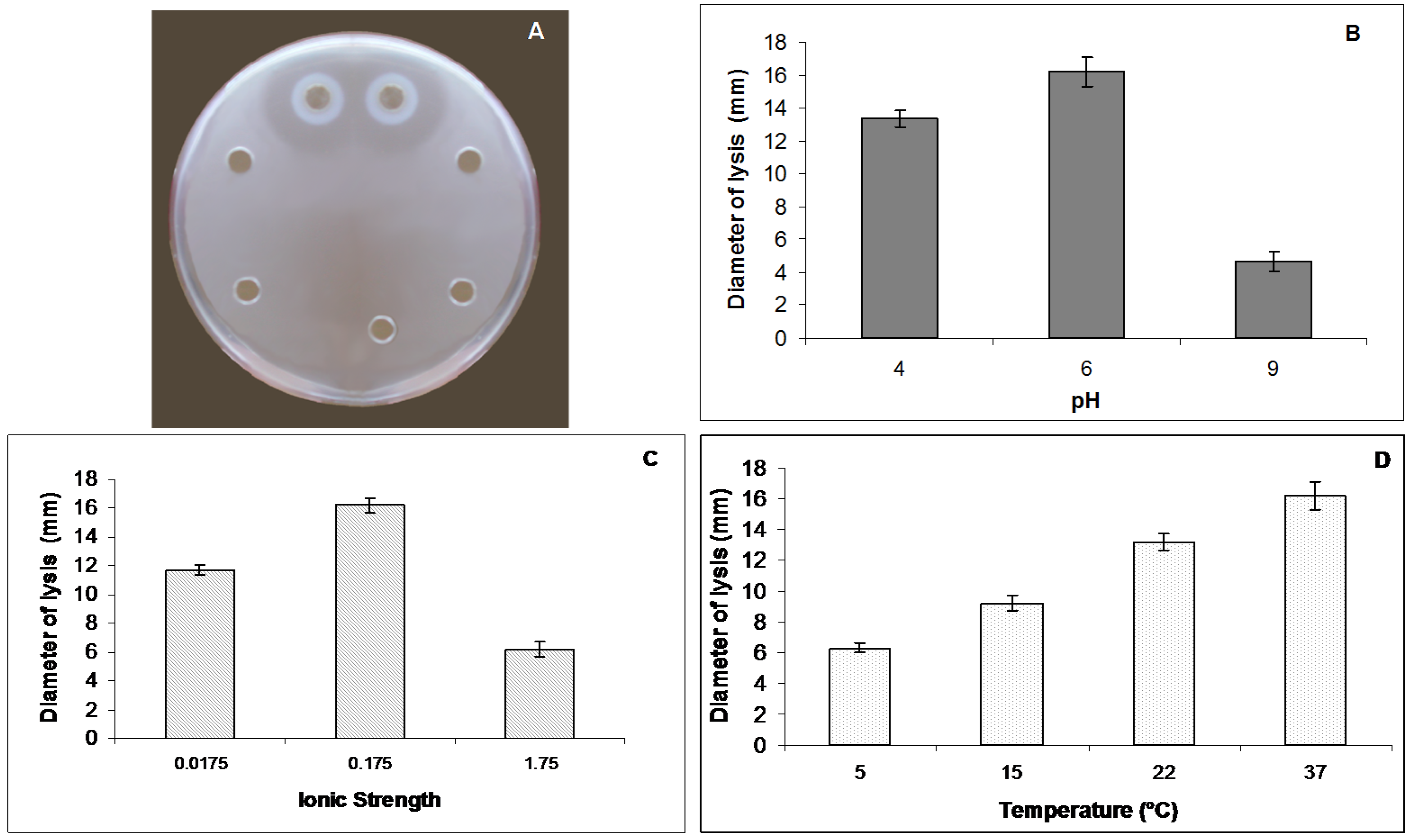
2.4. Lysozyme Like Activity

2.5. Hemolytic Activity
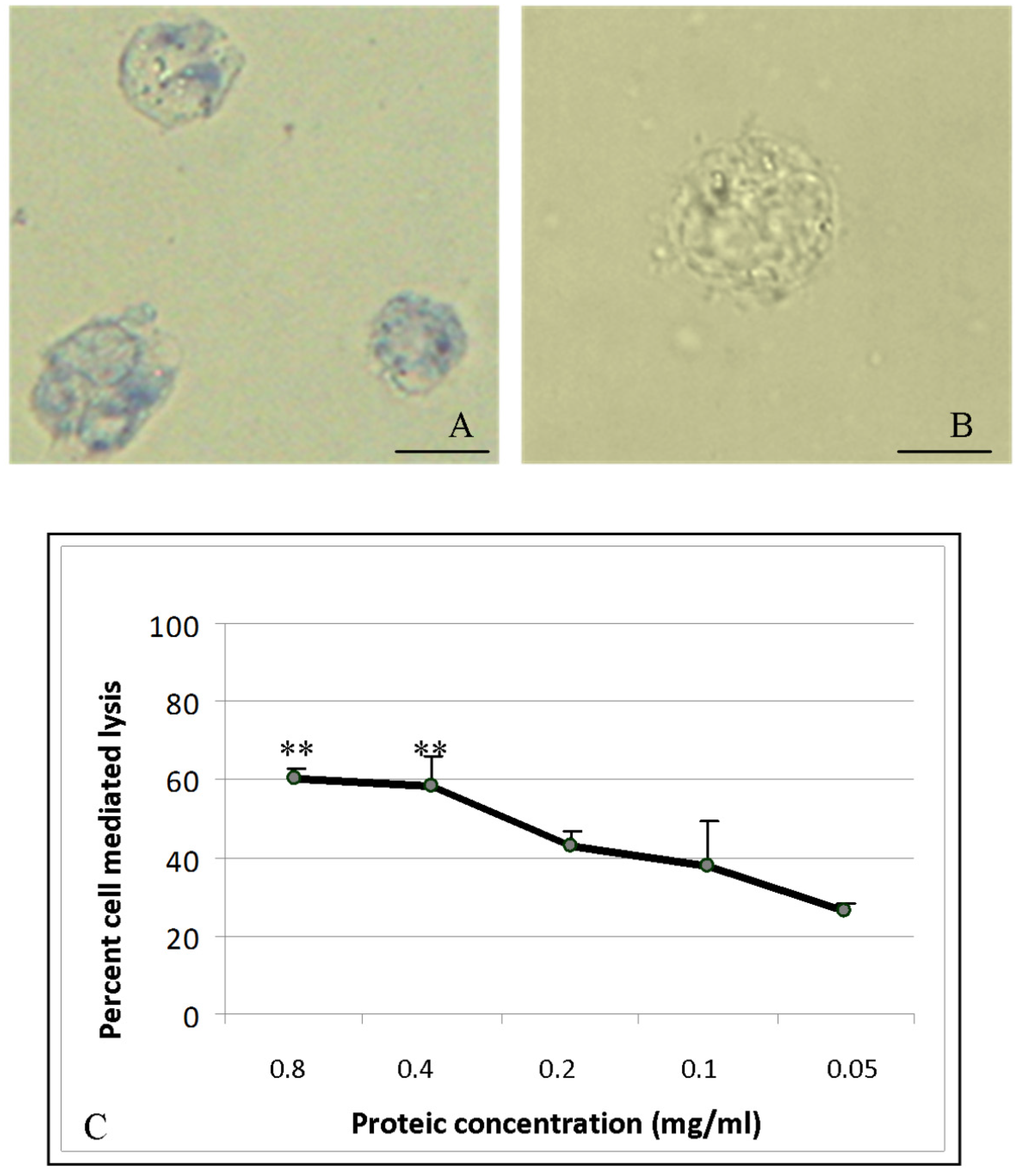
2.6. Cytotoxic Activity

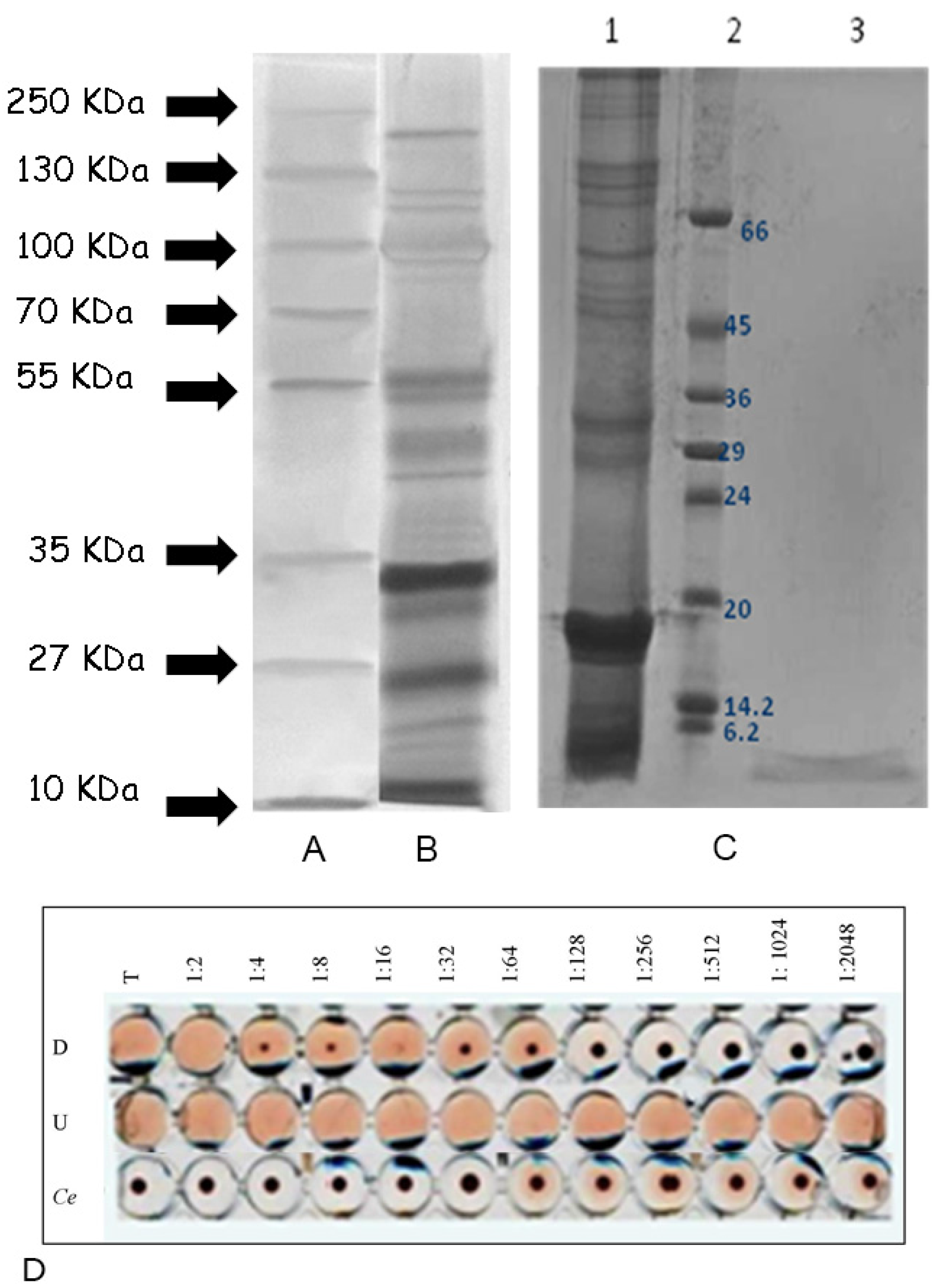
2.7. Fractionation of Actinia Equina Mucus

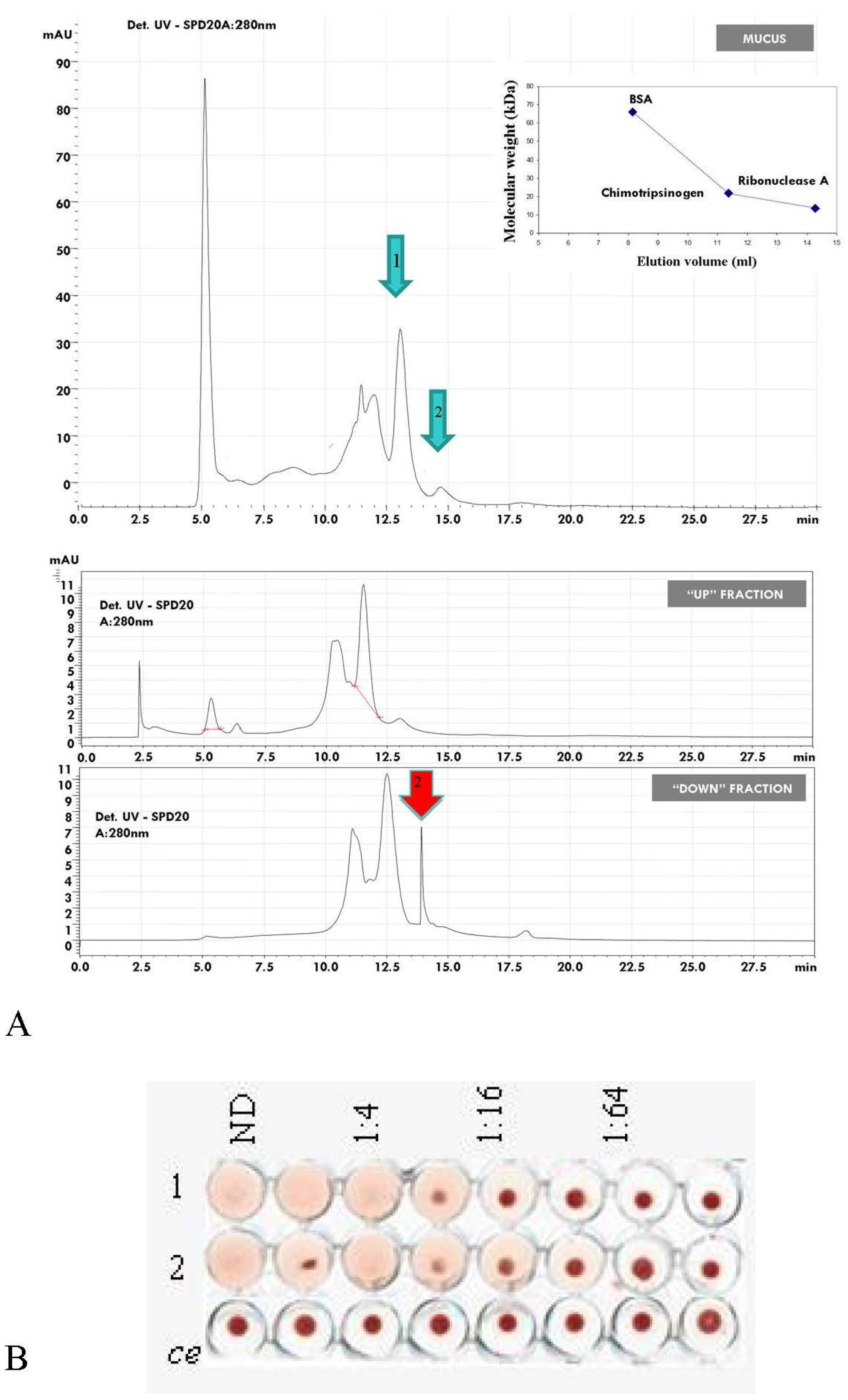
2.8. HPLC Separation of Mucus Components
3. Discussion
4. Experimental Section
4.1. Animals and Samples Preparation
4.2. Mucus Viscosity, Osmolarity, Electrical Conductivity and Water Content
4.3. Determination of the Inorganic Composition
4.4. Lipid, Protein, and Carbohydrate Concentration
4.5. Electrophoresis
4.6. Lysozyme-Like Activity
4.7. Hemolytic Activity
4.8. Cytoxicity Assay against the Tumor Cell Line K562
4.9. Fractionation of Actinia Equina Mucus
4.10. HPLC Size Exclusion Chromatography
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Connor, V.M. The use of mucous trails by intertidal limpets to enhance food resources. Biol. Bull. 1986, 171, 548–564. [Google Scholar] [CrossRef]
- Davies, M.S.; Jones, H.D.; Hawkins, S.J. Seasonal variation in the composition of pedal mucus from Patella vulgata L. J. Exp. Mar. Biol. Ecol. 1990, 144, 101–112. [Google Scholar] [CrossRef]
- Davis, J.M.; Viney, C. Water-mucin phases: Conditions for mucus liquid crystallinity. Thermochim. Acta 1998, 315, 39–49. [Google Scholar] [CrossRef]
- Smith, A.M.; Quick, T.J.; Peter, S.T.R.L. Differences in the Composition of Adhesive and Non-Adhesive Mucus from the Limpet Lottia limatula. Biol. Bull. 1999, 196, 34–44. [Google Scholar] [CrossRef] [PubMed]
- Kamino, K. Underwater adhesive of marine organisms as the vital link between biological science and material science. Mar. Biotechnol. 2008, 10, 111–121. [Google Scholar] [CrossRef] [PubMed]
- Branch, G.M. The biology of limpets: Physical factors, energy flow and ecological interactions. Oceanogr. Mar. Biol. Ann. Rev. 1981, 19, 235–380. [Google Scholar]
- Martin, R.; Walther, P. Protective mechanisms against the action of nematocysts in the epidermis of Cratena peregrina and Flabellina affinis (Gastropoda, Nudibranchia). Zoomorphology 2003, 122, 25–35. [Google Scholar]
- Baier, R.E.; Gucinski, H.; Meenaghan, M.A.; Wirth, J.; Glantz, P.Q. Biophysical studies of mucosal surfaces. In Oral Interfacial Reactions of Bone, Soft Tissue and Saliva; Glantz, P.Q., Leach, S.A., Ericson, T., Eds.; IRL Press: Oxford, UK, 1985; pp. 83–95. [Google Scholar]
- Clare, A.S. Marine natural product antifoulants: Status and potential. Biofouling 1995, 9, 211–229. [Google Scholar] [CrossRef]
- Stabili, L.; Schirosi, R.; Licciano, M.; Giangrande, A. The mucus of Sabella spallanzanii (Annelida, Polychaeta): Its involvement in chemical defence and fertilization success. J. Exp. Mar. Biol. Ecol. 2009, 374, 144–149. [Google Scholar] [CrossRef]
- Iori, D.; Forti, L.; Massamba-N’Siala, G.; Prevedelli, D.; Simonini, R. Toxicity of the purple mucus of the polychaete Halla parthenopeia (Oenonidae) revealed by a battery of ecotoxicological bioassays. Sci. Mar. 2014, 78, 589–595. [Google Scholar]
- Stabili, L.; Schirosi, R.; Licciano, M.; Giangrande, A. Role of Myxicola infundibulum (Polychaeta, Annelida) mucus: From bacterial control to nutritional home site. J. Exp. Mar. Biol. Ecol. 2014, 461, 344–349. [Google Scholar] [CrossRef]
- Waite, J.H. Nature’s underwater adhesive specialist. Int. J. Adhes. Adhes. 1987, 7, 9–14. [Google Scholar] [CrossRef]
- Derby, C.D. Escape by inking and secreting: Marine molluscs avoid predators through a rich array of chemicals and mechanisms. Biol. Bull. 2007, 213, 274–289. [Google Scholar] [CrossRef] [PubMed]
- Mayer, A.M.; Rodriguez, A.D.; Taglialatela-Scafati, O.; Fusetani, N. Marine Pharmacology in 2009–2011: Marine Compounds with Antibacterial, Antidiabetic, Antifungal, Anti-Inflammatory, Antiprotozoal, Antituberculosis, and Antiviral Activities; Affecting the Immune and Nervous Systems, and other Miscellaneous Mechanisms of Action. Mar. Drugs 2013, 11, 2510–2573. [Google Scholar] [PubMed]
- Aneiros, A.; Garateix, A. Bioactive peptides from marine sources: Pharmacological properties and isolation procedures. J. Chromatogr. B Anal. Technol. Biomed. Life Sci. 2004, 803, 41–53. [Google Scholar] [CrossRef] [PubMed]
- Otero Gonzalez, A.J.; Magalhaes, B.S.; Garcia Villarino, M.; Lopez Abarrategui, C.; Sousa, D.A.; Dias, S.C.; Franco, O.L. Antimicrobial peptides from marine invertebrates as a new frontier for microbial infection control. FASEB J. 2010, 24, 1320–1334. [Google Scholar] [CrossRef] [PubMed]
- Smith, V.J.; Desbois, A.P.; Dyrynda, E.A. Conventional and unconventional antimicrobials from fish, marine invertebrates and micro-algae. Mar. Drugs 2010, 8, 1213–1262. [Google Scholar] [CrossRef] [PubMed]
- Bavington, C.D.; Lever, R.; Mulloy, B.; Grundy, M.M.; Page, C.P.; Richardson, N.V.; McKenzie, J.D. Anti-adhesive glycoproteins in echinoderm mucus secretions. Comp. Biochem. Physiol. B Biochem. Mol. Biol. 2004, 139, 607–617. [Google Scholar] [CrossRef] [PubMed]
- Fountain, D.W.; Campbell, B.A. A lectin isolated from mucus of Helix aspersa. Comp. Biochem. Physiol. 1984, 77, 419–425. [Google Scholar]
- McKenzie, J.D.; Grigovala, I.V. The echinoderm surface and its role in preventing microfouling. Biofouling 1996, 10, 261–272. [Google Scholar] [CrossRef] [PubMed]
- Canicatti, C.; D’Ancona, G. Biological protective substances in Marthasterias glacialis (Asteroidea) epidermal secretion. J. Zool. 1990, 222, 445–454. [Google Scholar] [CrossRef]
- Stabili, L.; Schirosi, R.; Di Benedetto, A.; Merendino, A.; Villanova, L.; Giangrande, A. First insights into the biochemistry of Sabella spallanzanii (Annelida: Polychaeta) Mucus: A potentially unexplored resource for applicative purposes. J. Mar. Biol. Assoc. UK 2011, 91, 199–208. [Google Scholar] [CrossRef]
- Calow, P. Why some metazoan mucus secretions are more susceptible to microbial attack than others. Am. Nat. 1979, 114, 149–152. [Google Scholar] [CrossRef]
- Azam, F. Microbial control of oceanic carbon flux: The plot thickens. Science 1998, 280, 694–696. [Google Scholar] [CrossRef]
- Azam, F.; Smith, D.C.; Steward, G.F.; Hagström, A. Bacteria-organic matter coupling and its significance for oceanic carbon cycling. Microbial Ecol. 1993, 28, 167–179. [Google Scholar] [CrossRef] [PubMed]
- Peduzzi, P.; Herndl, G.J. Mucus trails in the rocky intertidal: A highly active microenvironment. Mar. Ecol. Prog. Ser. 1991, 75, 267–274. [Google Scholar] [CrossRef]
- Davies, M.S.; Hawkins, S.J.; Jones, H.D. Pedal mucus and its influence on the microbial food supply of two intertidal gastropods, Patella vulgata L. and Littorina littorea (L.). J. Exp. Mar. Biol. Ecol. 1992, 161, 57–77. [Google Scholar] [CrossRef]
- Imrie, D.W. The role of pedal mucus in the feeding behaviour of Littorina littorea (L.). In Proceedings of the 3rd International Symposium on Littorinid Biology, Dale Fort Field Centre, Wales, UK, 5–12 September 1990; Grahame, J., Mill, P.J., Reid, D.G., Eds.; The Malacological Society of London: London, UK, 1992; p. 221. [Google Scholar]
- Wild, C.; Woyt, H.; Markus Huettel, M. Influence of coral mucus on nutrient fluxes in carbonate sand. Mar. Ecol. Progr. Ser. 2005, 287, 87–98. [Google Scholar] [CrossRef]
- Gunasundari, V.; Ajith Kumar, T.T.; Kumaresan, S.; Balagurunathan, R.; Balasubramanian, T. Isolation of aliphatic-antibiotic compounds from marine invertebrate, Heteractis magnifica “Quoy & Gaimard1833” against captive marine ornamental fish pathogens. Indian J. Geomar. Sci. 2013, 42, 807–811. [Google Scholar]
- Lin, X.Y.; Ishida, M.; Nagashima, Y.; Shiomi, K. A polypeptide toxin in the sea anemone Actinia equina homologous with other sea anemone sodium channel toxins: Isolation and amino acid sequence. Toxicon 1996, 34, 57–65. [Google Scholar] [CrossRef]
- Frazão, B.; Vasconcelos, V.; Antunes, A. Sea Anemone (Cnidaria, Anthozoa, Actiniaria) Toxins: An Overview. Mar. Drugs 2012, 10, 1812–1851. [Google Scholar] [CrossRef] [PubMed]
- Flammang, P.; Santos, R.; Haesaerts, D. Echinoderm adhesive secretions: From experimental characterization to Evidence-Based Complementary and Alternative Medicine biotechnological applications. In Progress in Molecular and Subcellular Biology, Marine Molecular Biotechnology, Echinodermata; Matranga, V., Ed.; Springer: Berlin, Germany, 2005; pp. 201–220. [Google Scholar]
- Taylor, S.W.; Waite, J.H. Marine adhesives: From molecular dissection to application. In Protein-Based Materials; McGrath, K., Kaplan, D., Eds.; Birkhauser: Boston, MA, USA, 1997; pp. 217–248. [Google Scholar]
- Hamwood, T.E.; Cribb, B.W.; Halliday, J.A.; Kearn, G.C.; Whittington, I.D. Preliminary characterization and extraction of anterior adhesive secretion in monogean (plathyelminth) parasites. Folia Parasitol. 2002, 49, 39–49. [Google Scholar] [CrossRef] [PubMed]
- DeMoor, S.; Waite, H.; Jangoux, M.; Patrick, P. Characterization of the adhesive from Cuvierian tubules of the sea cucumber Holothuria forskali (Echinodermata, Holothuroidea). Mar. Biotechnol. 2003, 5, 45–57. [Google Scholar] [CrossRef] [PubMed]
- Pawlicki, J.M.; Pease, L.B.; Pierce, C.M.; Startz, T.P.; Zhang, Y.; Smith, A.M. The effect of molluscan glue proteins on gel mechanics. J. Exp. Biol. 2004, 207, 1127–1135. [Google Scholar] [CrossRef] [PubMed]
- Santos, R.; da Costa, G.; Franco, C.; Gomes-Alves, P.; Flammang, P.; Coelho, A.V. First insights into the biochemistry of tube foot adhesive from the sea urchin Paracentrotus lividus (Echinoidea, Echinodermata). Mar. Biotechnol. 2009, 11, 686–698. [Google Scholar] [CrossRef] [PubMed]
- Smith, A.M.; Morin, M.C. Biochemical differences between trail mucus and adhesive mucus from marsh periwinkles. Biol. Bull. 2002, 203, 338–346. [Google Scholar] [CrossRef] [PubMed]
- Flammang, P.; Walker, G. Measurement of the adhesion of the podia in the asteroid Asterias rubens (Echinodermata). J. Mar. Biol. Assoc. UK 1997, 77, 1251–1254. [Google Scholar] [CrossRef]
- Flammang, P.A.; van Cauwenberge, M.A.; Alexandre, H.; Jangoux, M. A study of temporary adhesion of the podia in the sea star Asterias rubens (Echinodermata, Asteroidea) through their footprints. J. Exp. Biol. 1998, 201, 2383–2395. [Google Scholar] [PubMed]
- Smith, A.M. The biochemistry and mechanics of gastropod adhesive gels. In Biological Adhesives; Smith, A.M., Callow, J.A., Eds.; Springer-Verlag: Berlin, Germany, 2006; pp. 167–182. [Google Scholar]
- Kamino, K. Barnacle underwater attachment. In Biological Adhesives; Smith, A.M., Callow, J.A., Eds.; Springer-Verlag: Berlin, Germany, 2006; pp. 145–166. [Google Scholar]
- Jolles, P.; Jolles, J. What’s new in lysozyme research? Mol. Cell. Biochem. 1984, 63, 165–189. [Google Scholar] [CrossRef] [PubMed]
- Lesser, M.P.; Stochaj, W.R.; Tapley, D.W.; Shick, J.M. Bleaching in coral reef anthozoans: Effects of irradiance, ultraviolet radiation, and temperature on the activities of protective enzymes against active oxygen. Coral Reefs 1995, 8, 225–232. [Google Scholar] [CrossRef]
- Leclerc, M. Humoral factors in marine invertebrates. In Molecular and Subcellular Biology: Invertebrate Immunology; Rinkevich, B., Muller, W.E.G., Eds.; Springer-Verlag: Berlin, Germany, 1996; pp. 1–9. [Google Scholar]
- Cheng, T.C.; Rodrick, G.E. Identification and characterization of lysozyme from the hemolymph of the soft-shelled clam, Mya arenaria. Biol. Bull. 1974, 147, 311–320. [Google Scholar] [CrossRef] [PubMed]
- Maginot, N.; Samain, J.F.; Daniel, J.Y.; Le Coz, J.R.; Moal, J. Kinetic properties of lysozyme from the digestive glands of Ruditapes philippinarum. Oceanis 1989, 15, 451–464. [Google Scholar]
- Sotelo-Mundo, R.R.; Islas-Osuna, M.A.; de-la-Re-Vega, E.; Hernandez-Lopez, J.; Vargas-Albores, F.; Yepiz-Plascencia, G. cDNA cloning of the lysozyme of the white shrimp Penaeus vannamei. Fish Shellfish Immunol. 2003, 15, 325–333. [Google Scholar] [CrossRef]
- Huag, K.; Olsen, Ø.M.; Sandsdalen, E.; Styrvold, O.B. Antibacterial activities in various tissues of the horse mussel Modiolus modiolus. J. Invertebr. Pathol. 2004, 85, 112–119. [Google Scholar] [CrossRef] [PubMed]
- Xue, O.G.; Schey, K.L.; Volety, A.K.; Chu, F.L.E.; La Peyre, J.F. Purification and characterization of lysozyme from plasma of the eastern oyster Crassostrea virginica. Comp. Biochem. Physiol. B Biochem. Mol. Biol. 2004, 139, 1–25. [Google Scholar] [CrossRef] [PubMed]
- Kristan, K.; Viero, G.; Dalla Serra, M.; Macek, P.; Anderluh, G. Molecular mechanism of pore formation by actinoporins. Toxicon 2009, 54, 1125–1134. [Google Scholar] [CrossRef] [PubMed]
- Monastyrnaya, M.I.; Leychenko, E.; Isaeva, M.; Likhatskaya, G.; Zelepuga, E.; Kostina, E.; Trifonov, E.; Nurminski, E.; Kozlovskaya, E. Actinoporins from the sea anemones, tropical Radianthus macrodactylus and northern Oulactis orientalis: Comparative analysis of structure-function relationships. Toxicon 2010, 56, 1299–1314. [Google Scholar] [CrossRef] [PubMed]
- Slattery, M.; McClintock, J.B.; Heine, J.N. Chemical defences in Antarctic soft corals: Evidence for anti-fouling compounds. J. Exp. Mar. Biol. Ecol. 1995, 190, 61–77. [Google Scholar] [CrossRef]
- Ibrahim, H.R.; Aoki, T.; Pellegrini, A. Strategies for new antimicrobial proteins and peptides: Lysozyme and aprotinin as model molecules. Curr. Pharm. Des. 2002, 8, 671–693. [Google Scholar] [CrossRef] [PubMed]
- Niyonsaba, F.; Ogawa, H. Protective roles of the skin against infection: Implication of naturally occurring human antimicrobial agents β-defensins, cathelicidin LL-37 and lysozyme. J. Dermatol. Sci. 2005, 40, 157–168. [Google Scholar] [CrossRef] [PubMed]
- Giraldi, T.; Ferlan, I.; Romeo, D. Antitumor activity of equinatoxin. Chem. Biol. Interact. 1976, 13, 199–203. [Google Scholar] [CrossRef]
- Mariottini, G.L.; Robbiano, L.; Carli, A. Toxicity of Actinia equina (Cnidaria: Anthozoa) crude venom on cultured cells. Boll. Soc. Ital. Biol. Sper. 1998, 74, 103–110. [Google Scholar] [PubMed]
- Soletti, R.C.; de Faria, G.P.; Vernal, J.; Terenzi, H.; Anderluh, G.; Borges, H.L.; Moura-Neto, V.; Gabilan, N.H. Potentiation of anticancer-drug cytotoxicity by sea anemone pore-forming proteins in human glioblastoma cells. Anticancer Drugs 2008, 19, 517–525. [Google Scholar] [CrossRef] [PubMed]
- Soletti, R.C.; Alves, T.; Vernal, J.; Terenzi, H.; Anderluh, G.; Borges, H.L.; Gabilan, N.H.; Moura-Neto, V. Inhibition of MAPK/ERK, PKC and CaMKII signaling blocks cytolysin-induced human glioma cell death. Anticancer Res. 2010, 30, 1209–1215. [Google Scholar] [PubMed]
- Parisi, M.G.; Trapani, M.R.; Cammarata, M. Granulocytes of sea anemone Actinia equina (Linnaeus, 1758) body fluid contain and release cytolysins forming plaques of lysis. ISJ 2014, 11, 39–46. [Google Scholar]
- Jelic-Mrcelic, G.; Sliskovic, M.; Antolic, B. Biofouling communities on test panels coated with TBT and TBT-free copper-based antifouling paints. Biofouling 2006, 22, 293–230. [Google Scholar] [CrossRef] [PubMed]
- Turley, P.A.; Fenn, R.J.; Ritter, J.C.; Callow, M.E. Pyrithiones as antifoulants: Environmental fate and loss of toxicity. Biofouling 2005, 21, 31–40. [Google Scholar] [CrossRef] [PubMed]
- Stupak, M.E.; Garcia, M.T.; Perez, M.C. Non-toxic alternative compounds for marine antifouling paints. Int. Biodeter. Biodegr. 2003, 52, 49–52. [Google Scholar] [CrossRef]
- Konstantinou, I.K.; Albanis, T.A. Worldwide occurrence and effects of antifouling paint booster biocides in the aquatic environment: A review. Environ. Int. 2004, 30, 235–248. [Google Scholar] [CrossRef]
- Ostroumov, S.A. On the concepts of biochemical ecology and hydrobiology: Ecological chemomediators. Contemp. Probl. Ecol. 2008, 1, 238–244. [Google Scholar] [CrossRef]
- Barresi, G.; di Carlo, E.; Trapani, M.R.; Parisi, M.G.; Chille, C.; Mule, M.F.; Cammarata, M.; Palla, F. Marine organisms as source of bioactive molecules applied in restoration projects. Herit. Sci. 2015, 3. [Google Scholar] [CrossRef]
- Rosen, M.W.; Cornford, N.E. Fluid friction of fish slimes. Nature 1971, 234, 49–51. [Google Scholar] [CrossRef]
- Cone, R.A. Mucus. In Mucosal Immunology; Ogra, P.L., Mestecky, J., Lamm, M.E., Strober, W., Bienenstock, J., McGhee, J.R., Eds.; Academic Press: San Diego, CA, USA, 1999; pp. 43–64. [Google Scholar]
- Withers, P.C. Comparative Animal Physiology; Saunders College Publishing: Ft. Worth, TX, USA, 1992; p. 669. [Google Scholar]
- Kitson, R.E.; Mellon, M.G. Colorimetric determination of phosphorus as molybdivanadophosphoric acid. Ind. Eng. Chem. Anal. 1944, 16, 379–383. [Google Scholar] [CrossRef]
- Quinlan, K.P.; Desesa, M.A. Spectrophotometric determination of phosphorus as molybdovanadophosphoric acid. Anal. Chem. 1955, 27, 1626–1629. [Google Scholar] [CrossRef]
- Folch, J.; Less, M.; Stone Stanley, G.H. A simple method for the isolation and purification of total lipids from animal tissues. J. Biol. Chem. 1957, 226, 497–508. [Google Scholar] [PubMed]
- Zöllner, N.; Kirsch, K. Determination of the total lipid concentration in serum. Z. Gesamte Exp. Med. 1962, 135, 545–549. [Google Scholar] [CrossRef]
- Bradford, M. A rapid and sensitive method for the quantification of microgram quantities of protein utilizing the principle of proteins dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Dubois, M.; Gilles, K.A.; Hamilton, J.K.; Rebers, P.A.; Smith, F. Colorimetric method for determination of sugars and related substances. Anal. Chem. 1956, 28, 350–356. [Google Scholar] [CrossRef]
- Kennedy, J.F.; Pagliuca, G. Chapter 2. Oligosaccharides. In Carbohydrate Analysis A Practical Approach, 2nd ed.; Chapling, M.F., Kennedy, J.F., Eds.; Oxford University Press: New York, NY, USA, 1994; pp. 43–72. [Google Scholar]
- Laemmli, V.H. Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nat. Lond. 1970, 227, 680–682. [Google Scholar] [CrossRef]
- Hames, B.D. One-dimensional polyacrylamide gel electrophoresis. In IRL Gel Electrophoresis of Proteins. A Practical Approach; Hames, B.D., Rickwood, D., Eds.; Press Oxford: Oxford, UK, 1990; pp. 1–147. [Google Scholar]
- Korzeniewski, C.; Callewaert, D.M. An enzyme-release assay for natural cytotoxicity. J. Immunol. Methods 1983, 64, 313–320. [Google Scholar] [CrossRef]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Stabili, L.; Schirosi, R.; Parisi, M.G.; Piraino, S.; Cammarata, M. The Mucus of Actinia equina (Anthozoa, Cnidaria): An Unexplored Resource for Potential Applicative Purposes. Mar. Drugs 2015, 13, 5276-5296. https://doi.org/10.3390/md13085276
Stabili L, Schirosi R, Parisi MG, Piraino S, Cammarata M. The Mucus of Actinia equina (Anthozoa, Cnidaria): An Unexplored Resource for Potential Applicative Purposes. Marine Drugs. 2015; 13(8):5276-5296. https://doi.org/10.3390/md13085276
Chicago/Turabian StyleStabili, Loredana, Roberto Schirosi, Maria Giovanna Parisi, Stefano Piraino, and Matteo Cammarata. 2015. "The Mucus of Actinia equina (Anthozoa, Cnidaria): An Unexplored Resource for Potential Applicative Purposes" Marine Drugs 13, no. 8: 5276-5296. https://doi.org/10.3390/md13085276
APA StyleStabili, L., Schirosi, R., Parisi, M. G., Piraino, S., & Cammarata, M. (2015). The Mucus of Actinia equina (Anthozoa, Cnidaria): An Unexplored Resource for Potential Applicative Purposes. Marine Drugs, 13(8), 5276-5296. https://doi.org/10.3390/md13085276