
The Influence of Tetrodotoxin (TTX) on the Distribution and Chemical Coding of Caudal Mesenteric Ganglion (CaMG) Neurons Supplying the Porcine Urinary Bladder

 , ,
, ,
Abstract
:1. Introduction
2. Results
2.1. Distribution of Fast Blue-Positive (FB+) Neurons in the Control Pigs
2.2. Distribution of FB+ Neurons in the TTX-Treated Pigs
2.3. Immunohistochemical Characteristics of FB+ Neurons in the Control Pigs
2.4. Immunohistochemical Characteristics of FB+ Neurons in TTX-Treated Animals
3. Discussion
4. Materials and Methods
4.1. Laboratory Animals
4.2. Surgical Procedures
4.3. Sectioning of the Ganglia, Estimation of the Total Number of the CaMG-UBPN and Immunohistochemical Procedure
4.4. Control of Specificity of the Tracer Staining and Immunohistochemical Procedures
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Miyazawa, K.; Noguchi, T. Distribution and origin of tetrodotoxin. J. Toxicol. Toxin Rev. 2001, 20, 11–33. [Google Scholar] [CrossRef]
- Kim, Y.H.; Brown, G.B.; Mosher, F.A. Tetrodotoxin: Occurrence in atelopid frogs of Costa Rica. Science 1975, 189, 151–152. [Google Scholar] [CrossRef] [PubMed]
- Yasumoto, T.; Yotsu, M.; Murate, M.; Naoki, H. New tetrodotoxin analogues from the newt Cynops ensicauda. J. Am. Chem. Soc. 1988, 110, 2344–2345. [Google Scholar] [CrossRef]
- Yotsu-Yamashita, M.; Gilhen, J.; Russell, R.W.; Krysko, K.L.; Melaun, C.; Kurz, A.; Kauferstein, S.; Kordis, D.; Mebs, D. Variability of tetrodotoxin and of its analogues in the red-spotted newt, Notophthalmus viridescens (Amphibia: Urodela: Salamandridae). Toxicon 2012, 59, 257–264. [Google Scholar] [CrossRef] [PubMed]
- Yotsu-Yamashita, M.; Mebs, D.; Kwet, A.; Schneider, M. Tetrodotoxin and its analogue 6-epitetrodotoxin in newts (Triturus spp.; Urodela, Salamandridae) from southern Germany. Toxicon 2007, 50, 306–309. [Google Scholar] [CrossRef] [PubMed]
- Noguchi, T.; Arakawa, O. Tetrodotoxin–Distribution and Accumulation in Aquatic Organisms, and Cases of Human Intoxication. Mar. Drugs 2008, 6, 220–242. [Google Scholar] [CrossRef] [PubMed]
- Lago, J.; Rodríguez, L.P.; Blanco, L.; Vieites, J.M.; Cabado, A.G. Tetrodotoxin, an Extremely Potent Marine Neurotoxin: Distribution, Toxicity, Origin and Therapeutical Uses. Mar. Drugs 2015, 13, 6384–6406. [Google Scholar] [CrossRef] [PubMed]
- Clark, R.F.; Williams, S.R.; Nordt, S.P.; Manoguerra, A.S. A review of selected seafood poisonings. Undersea Hyperb. Med. 1999, 26, 175–184. [Google Scholar] [PubMed]
- Denac, H.; Mevissen, M.; Scholtysik, G. Structure, function and pharmacology of voltage-gated sodium channels. Naunyn-Schmiedebergs Arch. Pharmacol. 2000, 362, 453–479. [Google Scholar] [CrossRef] [PubMed]
- Suehiro, M. Historical review on chemical and medical studies of globefish toxin before World War II (Article in Japanese). Yakushigaku Zasshi 1994, 29, 428–434. [Google Scholar] [PubMed]
- Marcil, J.; Walczak, J.S.; Guindon, J.; Ngoc, A.H.; Lu, S.; Beaulieu, P. Antinociceptive effects of tetrodotoxin (TTX) in rodents. Br. J. Anaesth. 2006, 96, 761–768. [Google Scholar] [CrossRef] [PubMed]
- Shi, J.; Liu, T.T.; Wang, X.; Epstein, D.H.; Zhao, L.Y.; Zhang, X.L.; Lu, L. Tetrodotoxin reduces cue-induced drug craving and anxiety in abstinent heroin addicts. Pharmacol. Biochem. Behav. 2009, 92, 603–607. [Google Scholar] [CrossRef] [PubMed]
- Green, P.G.; Alvarez, P.; Levine, J.D. Topical tetrodotoxin attenuates photophobia induced by corneal injury in the rat. J. Pain 2015, 16, 881–886. [Google Scholar] [CrossRef] [PubMed]
- Lodge, D.J.; Grace, A.A. Aberrant hippocampal activity underlies the dopamine dysregulation in an animal model of schizophrenia. J. Neurosci. 2007, 27, 11424–11430. [Google Scholar] [CrossRef] [PubMed]
- Song, H.; Li, J.; Lu, C.L.; Kang, L.; Xie, L.; Zhang, Y.Y.; Zhou, X.B.; Zhong, S. Tetrodotoxin alleviates acute heroin withdrawal syndrome: A multicentre, randomized, double-blind, placebo-controlled study. Clin. Exp. Pharmacol. Physiol. 2011, 38, 510–514. [Google Scholar] [CrossRef] [PubMed]
- Stewart, W.F.; Van Rooyen, J.B.; Cundiff, G.W.; Abrams, P.; Herzog, A.R.; Corey, R.; Hunt, T.L.; Wein, A.J. Prevalence and burden of overactive bladder in the United States. World J. Urol. 2003, 20, 327–336. [Google Scholar] [PubMed]
- Pinto, R.; Lopes, T.; Silva, J.; Silva, C.; Dinis, P.; Cruz, F. Persistent therapeutic effect of repeated injections of onabotulinum toxin a in refractory bladder pain syndrome/interstitial cystitis. J. Urol. 2013, 189, 548–553. [Google Scholar] [CrossRef] [PubMed]
- Santos-Silva, A.; da Silva, C.M.; Cruz, F. Botulinum toxin treatment for bladder dysfunction. Int. J. Urol. 2013, 20, 956–962. [Google Scholar] [CrossRef] [PubMed]
- Foster, H.E., Jr.; Lake, A.G. Use of vanilloids in urologic disorders. Prog. Drug Res. 2014, 68, 307–317. [Google Scholar] [PubMed]
- Szallasi, A.; Blumberg, P.M. Resiniferatoxin, a phorbol-related diterpene, acts as an ultrapotent analog of capsaicin, the irritant constituent in red pepper. Neuroscience 1989, 30, 515–520. [Google Scholar] [CrossRef]
- Cruz, F.; Guimaräes, M.; Silva, C.; Reis, M. Suppression of bladder hyperreflexia by intravesical resiniferatoxin. Lancet 1997, 350, 640–641. [Google Scholar] [CrossRef]
- Gray, S.M.; McGeown, J.G.; McMurray, G.; McCloskey, K.D. Functional innervation of Guinea-pig bladder interstitial cells of cajal subtypes: Neurogenic stimulation evokes in situ calcium transients. PLoS ONE 2013, 8, e53423. [Google Scholar] [CrossRef] [PubMed]
- Ramos-Filho, A.C.; Shah, A.; Augusto, T.M.; Barbosa, G.O.; Leiria, L.O.; de Carvalho, H.F.; Antunes, E.; Grant, A.D. Menthol inhibits detrusor contractility independently of TRPM8 activation. PLoS ONE 2014, 9, e111616. [Google Scholar]
- Kuga, N.; Tanioka, A.; Hagihara, K.; Kawai, T. Modulation of afferent nerve activity by prostaglandin E2 upon urinary bladder distension in rats. Exp. Physiol. 2016, 101, 577–587. [Google Scholar] [CrossRef] [PubMed]
- Lepiarczyk, E.; Bossowska, A.; Majewski, M. Changes in chemical coding of sympathetic chain ganglia (SChG) neurons supplying porcine urinary bladder after botulinum toxin (BTX) treatment. Cell Tissue Res. 2015, 360, 263–272. [Google Scholar] [CrossRef] [PubMed]
- Lepiarczyk, E.; Majewski, M.; Bossowska, A. The influence of intravesical administration of resiniferatoxin (RTX) on the chemical coding of sympathetic chain ganglia (SChG) neurons supplying the porcine urinary bladder. Histochem. Cell Biol. 2015, 144, 479–489. [Google Scholar] [CrossRef] [PubMed]
- Lepiarczyk, E.; Dudek, A.; Kaleczyc, J.; Majewski, M.; Markiewicz, W.; Radziszewski, P.; Bossowska, A. The influence of resiniferatoxin on the chemical coding of caudal mesenteric ganglion neurons supplying the urinary bladder in the pig. J. Physiol. Pharmacol. 2016, 67, 625–632. [Google Scholar] [PubMed]
- Dalmose, A.L.; Hvistendahl, J.J.; Olsen, L.H.; Eskild-Jensen, A.; Djurhuus, J.C.; Swindle, M.M. Surgically induced urologic models in swine. J. Investig. Surg. 2000, 13, 133–145. [Google Scholar]
- Kuzmuk, K.N.; Schook, L.B. Pigs as a model for biomedical sciences. In The Genetics of the Pig, 2nd ed.; Rothschild, M.F., Ruvinsky, A., Eds.; CAB International: Oxford Shire, UK, 2011; pp. 426–444. [Google Scholar]
- Swindle, M.M.; Makin, A.; Herron, A.J.; Clubb, F.J., Jr.; Frazier, K.S. Swine as models in biomedical research and toxicology testing. Vet. Pathol. 2012, 49, 344–356. [Google Scholar] [CrossRef] [PubMed]
- Bossowska, A.; Majewski, M. The influence of resiniferatoxin on the chemical coding of neurons in dorsal root ganglia supplying the urinary bladder in the female pig. Pol. J. Vet. Sci. 2012, 15, 135–142. [Google Scholar] [CrossRef] [PubMed]
- Bossowska, A.; Majewski, M. Tetrodotoxin induced changes in the chemical coding of dorsal root ganglion neurons supplying the porcine urinary bladder. Pol. J. Vet. Sci. 2012, 15, 355–363. [Google Scholar] [CrossRef] [PubMed]
- Bossowska, A. Distribution of primary afferent neurons associated with the porcine inferior mesenteric ganglion (IMG). Folia Histochem. Cytobiol. 2002, 40, 367–372. [Google Scholar] [PubMed]
- Fishbein, I.; Segal, M. Miniature synaptic currents become neurotoxic to chronically silenced neurons. Cereb. Cortex 2007, 17, 1292–1306. [Google Scholar] [CrossRef] [PubMed]
- Schonfeld-Dado, E.; Segal, M. Activity deprivation induces neuronal cell death: Mediation by tissue-type plasminogen activator. PLoS ONE 2011, 6, e25919. [Google Scholar] [CrossRef] [PubMed]
- Lysko, P.G.; Webb, C.L.; Yue, T.L.; Gu, J.L.; Feuerstein, G. Neuroprotective effects of tetrodotoxin as a Na+ channel modulator and glutamate release inhibitor in cultured rat cerebellar neurons and in gerbil global brain ischemia. Stroke 1994, 25, 2476–2482. [Google Scholar] [CrossRef] [PubMed]
- Andersson, K.E.; Arner, A. Urinary bladder contraction and relaxation: Physiology and pathophysiology. Physiol. Rev. 2004, 84, 935–986. [Google Scholar] [CrossRef] [PubMed]
- Burliński, P.J.; Burlińska, A.M.; Gonkowski, S.; Całka, J. Resiniferatoxin and tetrodotoxin induced NPY and TH immunoreactivity changes within the paracervical ganglion neurons supplying the urinary bladder. J. Mol. Neurosci. 2013, 49, 62–67. [Google Scholar] [CrossRef] [PubMed]
- Burliński, P.J.; Czujkowska, A.; Arciszewski, M.B.; Całka, J. Upregulation of LENK and VIP in paracervical ganglion neurons supplying the urinary bladder of tetrodotoxin- and resiniferatoxin-treated female pigs. Acta Vet. Hung. 2012, 60, 383–393. [Google Scholar] [CrossRef] [PubMed]
- Burliński, P.J.; Gonkowski, S.; Całka, J. Tetrodotoxin- and resiniferatoxin-induced changes in paracervical ganglion ChAT- and nNOS-IR neurons supplying the urinary bladder in female pigs. Acta Vet. Hung. 2011, 59, 455–463. [Google Scholar] [CrossRef] [PubMed]
- Keast, J.R. Unusual autonomic ganglia: Connections, chemistry and plasticity of pelvic ganglia. Int. Rev. Cytol. 1999, 193, 1–69. [Google Scholar] [PubMed]
- Tran, L.V.; Somogyi, G.T.; De Groat, W.C. Inhibitory effect of neuropeptide Y on adrenergic and cholinergic transmission in rat urinary bladder and urethra. Am. J. Physiol. 1994, 266 4 Pt 2, R1411–R1417. [Google Scholar] [PubMed]
- Ellis, J.L.; Burnstock, G. Neuropeptide Y neuromodulation of sympathetic co-transmission in the guinea-pig vas deferens. Br. J. Pharmacol. 1990, 100, 457–462. [Google Scholar] [CrossRef] [PubMed]
- Honda, M.; Yoshimura, N.; Inoue, S.; Hikita, K.; Muraoka, K.; Saito, M.; Chancellor, M.B.; Takenaka, A. Inhibitory role of the spinal galanin system in the control of micturition. Urology 2013, 82, 1188.e9–1188.e13. [Google Scholar] [CrossRef] [PubMed]
- Kaleczyc, J.; Timmermans, J.P.; Majewski, M.; Łakomy, M.; Mayer, B.; Scheuermann, D.W. NO-synthase-containing neurons of the pig inferior mesenteric ganglion, part of them innervating the ductus deferens. Acta Anat. (Basel) 1994, 151, 62–67. [Google Scholar] [CrossRef] [PubMed]
- Pidsudko, Z. Immunohistochemical characteristics and distribution of neurons in the paravertebral, prevertebral and pelvic ganglia supplying the urinary bladder in the male pig. J. Mol. Neurosci. 2014, 52, 56–70. [Google Scholar] [CrossRef] [PubMed]
- Bossowska, A.; Majewski, M. Botulinum toxin type A-induced changes in the chemical coding of dorsal root ganglion neurons supplying the porcine urinary bladder. Pol. J. Vet. Sci. 2012, 15, 345–353. [Google Scholar] [CrossRef] [PubMed]
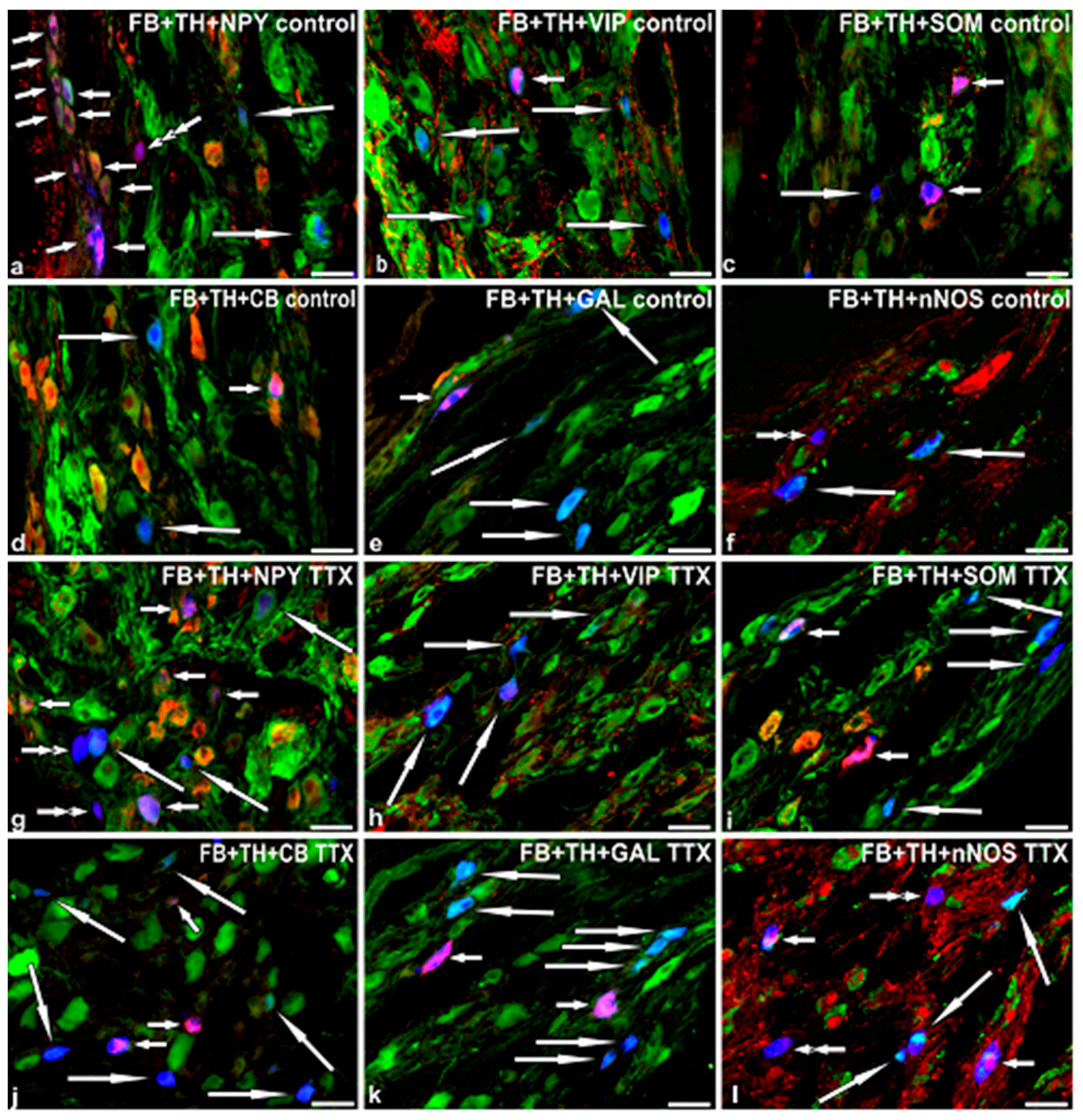

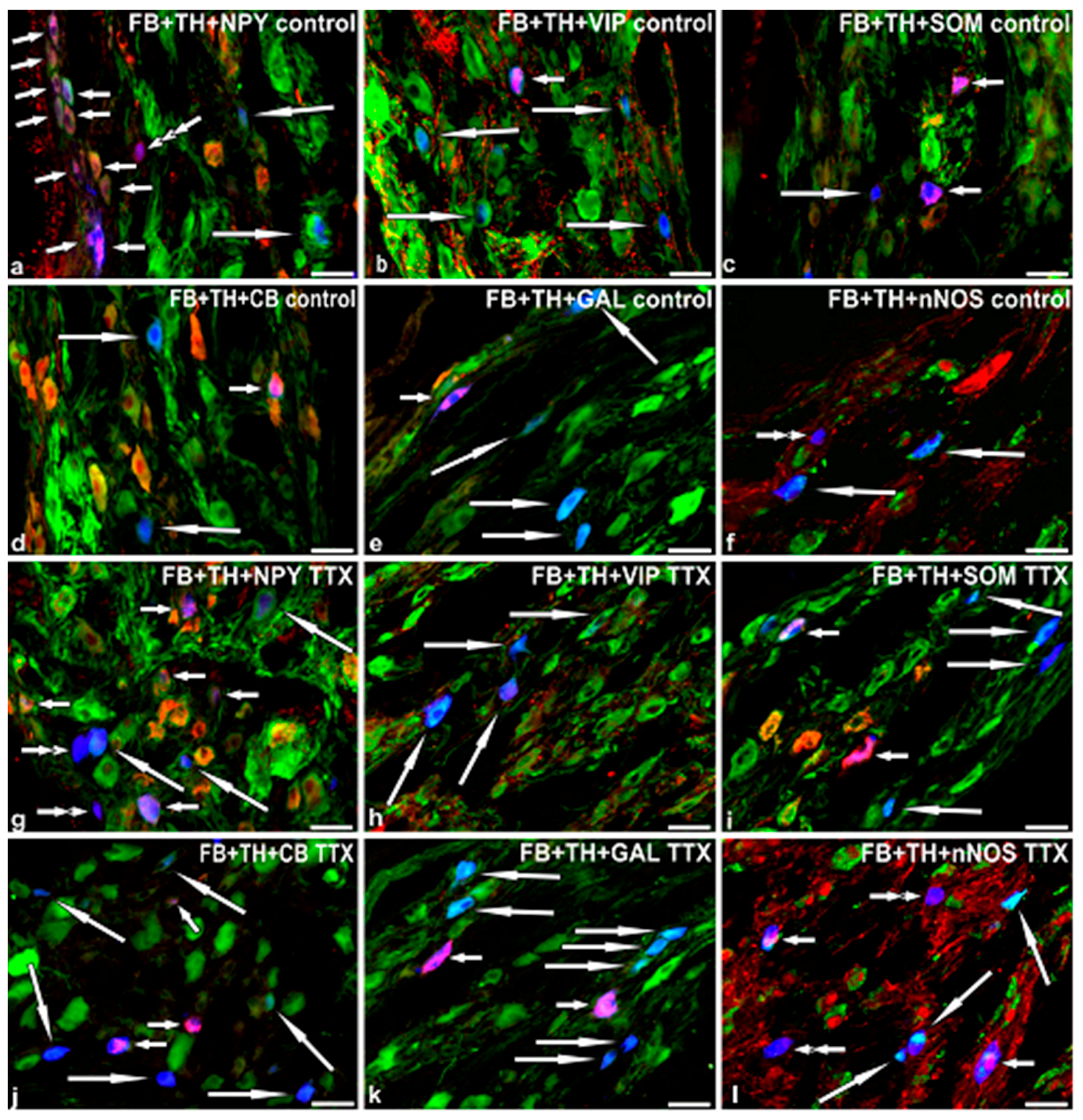{kind=link}
| Experimental Group | FB+/TH+ | FB+/TH+/NPY+ | FB+/TH+/VIP+ | FB+/TH+/SOM+ | FB+/TH+/CB+ | FB+/TH+/GAL+ | FB+/TH+/nNOS+ |
|---|---|---|---|---|---|---|---|
| Control pigs | 92.6% ± 3.4% | 88.9% ± 5.3% | 1.7% ± 0.6% | 8.8% ± 1.6% | 1.8% ± 0.7% | 1.7% ± 0.8% | 0% |
| TTX-treated pigs | 88.8% ± 2% * | 10.6% ± 5.3% *** | 0% *** | 39% ± 12.8% *** | 12.6% ± 2.7% *** | 10.9% ± 2.6% *** | 1.1% ± 0.3% ** |
| Experimental Group | FB+/TH− | FB+/TH−/NPY+ | FB+/TH−/VIP+ | FB+/TH−/SOM+ | FB+/TH−/CB+ | FB+/TH−/GAL+ | FB+/TH−/nNOS+ |
|---|---|---|---|---|---|---|---|
| Control pigs | 7.4% ± 3.4% | 27.3% ± 17.6% | 8.9% ± 6.8% | 0% | 0% | 0% | 0% |
| TTX-treated pigs | 11.2% ± 2% * | 1.2% ± 2% ** | 0% | 2.7% ± 1.5% ** | 1.9% ± 0.8% *** | 3% ± 1.4% *** | 4.2% ± 2.7% *** |
| Antigen | Code | Dilution | Host | Supplier |
|---|---|---|---|---|
| Primary antibodies | ||||
| TH | MAB318 | 1:400 | Mouse | Millipore, Temecula, CA, USA |
| NPY | NA 1233 | 1:8000 | Rabbit | Enzo Life Sciences; Farmingdale, NY, USA |
| VIP | VA 1285 | 1:4000 | Rabbit | Enzo Life Sciences; Farmingdale, NY, USA |
| SOM | 11180 | 1:4000 | Rabbit | Icn-Cappel, Aurora, OH, USA |
| CB | Lot No.: 0.7 | 1:9000 | Rabbit | Swant, Marly, Fribourg, Switzerland |
| GAL | AB 5909 | 1:1000 | Rabbit | Millipore, Temecula, CA, USA |
| nNOS | AB 5380 | 1:6000 | Rabbit | Millipore, Temecula, CA, USA |
| Secondary reagents | ||||
| Biotinylated anti-rabbit immunoglobulins | E 0432 | 1:1000 | Goat | Dako, Germany |
| CY3-conjugated streptavidin | 711-165-152 | 1:13,000 | - | Jackson I.R.; West Grove, PA, USA |
| FITC-conjugated anti-mouse IgG | 715-096-151 | 1:700 | Donkey | Jackson I.R.; West Grove, PA, USA |
| Antigen | Code | Supplier |
|---|---|---|
| TH | AC21-0699-P | Abcore, Ramona, CA, USA |
| NPY | N3266 | Sigma, St. Louis, MO, USA |
| VIP | V6130 | Sigma, St. Louis, MO, USA |
| SOM | S9129 | Sigma, St. Louis, MO, USA |
| CB | AC21-2748-P | Abcore, Ramona, CA, USA |
| GAL | G5773 | Sigma, St. Louis, MO, USA |
| nNOS | N3033 | Sigma, St. Louis, MO, USA |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lepiarczyk, E.; Bossowska, A.; Kaleczyc, J.; Majewska, M.; Gonkowski, S.; Majewski, M. The Influence of Tetrodotoxin (TTX) on the Distribution and Chemical Coding of Caudal Mesenteric Ganglion (CaMG) Neurons Supplying the Porcine Urinary Bladder. Mar. Drugs 2017, 15, 101. https://doi.org/10.3390/md15040101
Lepiarczyk E, Bossowska A, Kaleczyc J, Majewska M, Gonkowski S, Majewski M. The Influence of Tetrodotoxin (TTX) on the Distribution and Chemical Coding of Caudal Mesenteric Ganglion (CaMG) Neurons Supplying the Porcine Urinary Bladder. Marine Drugs. 2017; 15(4):101. https://doi.org/10.3390/md15040101
Chicago/Turabian StyleLepiarczyk, Ewa, Agnieszka Bossowska, Jerzy Kaleczyc, Marta Majewska, Sławomir Gonkowski, and Mariusz Majewski. 2017. "The Influence of Tetrodotoxin (TTX) on the Distribution and Chemical Coding of Caudal Mesenteric Ganglion (CaMG) Neurons Supplying the Porcine Urinary Bladder" Marine Drugs 15, no. 4: 101. https://doi.org/10.3390/md15040101